1. Introduction
Extracellular vesicles (EVs), membranous structures secreted by nearly all cell types, consist of microparticles and exosomes
[1]. The classification of EVs is based on the differences in formation mechanism and size. Microparticles are secreted by cells through direct membrane budding, whereas exosomes are secreted through the endosomal transport pathway. According to the guideline recommended by the International Society for Extracellular Vesicles (ISEV) in 2018, EVs are divided into small EVs (sEVs, <200 nm) and large EVs (lEVs, >200 nm). Additionally, sEVs are mainly composed of exosomes and a small amount of microparticles that are less than 200 nm
[2]. Exosomes inherit bioactive molecules from their parental cells, including nucleotides, proteins, and lipids. The crucial role of exosomes in intercellular communication has been widely recognized in the last 10 years, although they were once thought to be the excretion of cellular wastes for cell homeostasis. Numerous studies on targeted and functional interactions between exosomes and cells have revealed the significant biological functions of exosomes. Exosomes are involved in various physiopathological processes, such as embryonic development, tissue repair and regeneration, material metabolism, and immunoregulation
[3]. Specially, exosomes play a critical role in regulating tumorigenesis and tumor progression. Accumulating studies have reported the tumorigenic effect of exosomes, especially in resistance to therapy
[4]. A deeper investigation of exosomes in the disease state may contribute to a better understanding of the pathogenic mechanisms and help to develop innovative diagnostic or therapeutic strategies. This
re
ntryview summarizes the immunosuppressive effects of exosomes and discusses the potential clinical applications of exosomes in the diagnosis and treatment of tumor.
2. Life Course of Exosomes
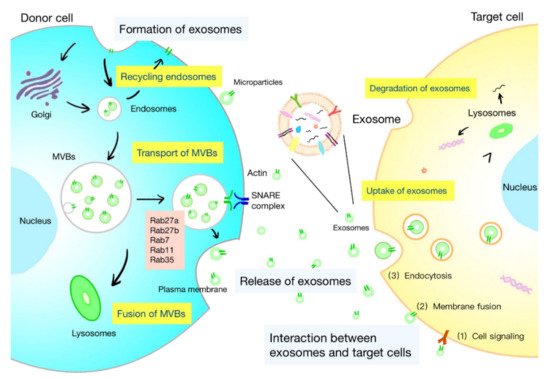
The life course of exosomes comprises the generation, secretion, transport of exosomes, and interaction with target cells, which is under the control of a series of complex regulatory molecules (Figure 1). A deeper understanding of the life course of exosomes is helpful to identify the functions of exosomes and develop strategies for specific regulation of exosomes.
Figure 1. Life course of exosomes. The formation of exosomes: initiation, endocytosis, multivesicular bodies (MVBs) formation, and secretion. The intracellular trafficking of MVBs is mediated by Rab GTPases. The fusion of MVBs with the plasma membrane is facilitated by SNAREs. There are three kinds of interactions between exosomes and cells: (1) the membrane proteins on the exosomes and target cells bind directly, and then trigger the intracellular signaling cascade; (2) exosomes transport their contents to target cells by fusing with the cell membrane; and (3) exosomes are engulfed by cells and degraded by lysosomes to release signal molecules.
2.1. Formation of Exosomes
The biogenesis of exosomes is a strictly regulated process that comprises four main stages: initiation, endocytosis, multivesicular bodies (MVBs) formation, and secretion (
Figure 1)
[5]. The generation of exosomes is mostly dependent on the endosomal sorting complex required for transport (ESCRT), which is composed of four complexes: ESCRT-0 (Hrs, Stam1, Stam2), ESCRT-I (Vps28, Vps37, Tsg101, etc.), ESCRT-II (Vps25, Vps36, etc.), and ESCRT-III (Vps2, Alix, etc.). It can recognize ubiquitinated proteins in vivo and assist in the transport of proteins and release of exosomes. The formation of MVBs is caused by an inward budding of the early endosomal membrane that is triggered by ceramide
[6]. MVBs are normally degraded by fusion with lysosomes, while some are secreted into the extracellular space by fusion with the plasma membrane, which are called exosomes.
2.2. Release of Exosomes
Exosomes are released into the extracellular space through the fusion of the MVB limiting membrane with the plasma membrane (
Figure 1). Rab GTPases mediate intracellular trafficking of MVBs and determine the fate of MVBs. Several Rab proteins, including Rab7, Rab11, Rab27, and Rab35, participate in exosome biogenesis. The release of flotillin-enriched exosomes and the transport of cargoes, such as transferrin receptors and signaling molecules, are reported to be regulated by Rab11 and Rab35
[1][7][1,7]. Certain membrane lipids are also found to participate in the secretion of exosomes. Phosphatidic acid originating from the activity of diacylglycerol kinase (DGK) or phospholipase D influences the secretion of exosomes
[8][9][8,9]. The effects of the ethylmaleimide-sensitive factor attachment protein receptor (SNARE) in exosome secretion have been proved
[10]. The fusion of MVBs with the plasma membrane is facilitated by the interaction between vesicular soluble SNAREs (v-SNAREs) on MVBs and target SNAREs (t-SNAREs) on the target membrane.
2.3. Interaction between Exosomes and Target Cells
After being released, exosomes are widely distributed to the blood, saliva, urine, cerebrospinal fluid, ascites, and pleural fluid. Exosomes can interact with both adjacent and distant cells to affect their cell activity and functions. Understanding the interplay between exosomes and target cells may help interpret the influences of exosomes on target cells. The interaction between exosomes and cells can be summarized as follows (
Figure 1): (1) the membranal proteins on the exosomes and target cells bind directly, and then trigger the intracellular signaling cascade in target cells; (2) exosomes transport their contents to target cells by fusing with the cell membrane; and (3) exosomes are engulfed by cells and degraded by lysosomes to release signal molecules
[5]. Besides, some mediators of these interactions have been found, including integrins, lipids, tetrapeptide, heparan sulfate proteoglycans, and extracellular matrix components
[1]. The internalization of exosomes is suggested to be the primary interaction between exosomes and target cells. Exosomes are internalized through the endocytic pathway and transported to lysosomes, where the proteins and lipids in the exosomes are degraded to offer relevant metabolite sources to target cells
[11]. Studies have shown the restricted colocalization of exosomes with early endosomes. It is believed that intraluminal vesicles could transfer contents to recipient cells by fusion with MVBs instead of lysosome
[1].
3. Exosomes as Cancer Biomarkers
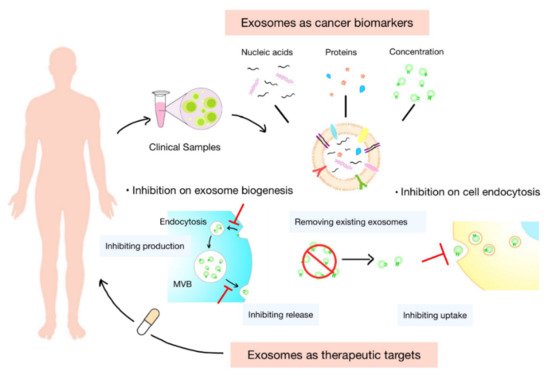
Exosomes are ideal substitutes for several biologically active molecules, such as proteins and transcripts. Cargoes carried by exosomes are inherited from their parental cells and partly reflect cell characteristics. They are protected by the membrane of exosomes from degradation during transportation. In addition, exosomes are widely distributed in various body fluids. These features of exosomes offer enormous potential for their use in the diagnosis and prognosis of tumor, as well as the prediction of therapeutic response (Figure 2 3).
Figure 23. Clinical potential of exosomes. Exosomes normally serve as biomarkers or therapeutic targets in clinical settings. The concentration of exosomes and cargoes carried by them, such as nucleic acids and proteins, are potential diagnostic biomarkers of tumor and prognostic indicators of treatments. Inhibiting the biogenesis, release, or uptake of exosomes and removing the circulating cancer exosomes could be novel targets for anticancer therapies.
3.1. Concentration of Exosomes
Exosomes are widely distributed to a variety of body fluids, including the blood, urine, saliva, cerebrospinal fluid
[12][104]. Exosomes can serve as diagnostic indicators in “liquid biopsy” for various malignancies
[13][14][105,106]. Numerous preclinical studies have verified the potential utility of exosomes in detecting and monitoring cancer. It is reported that the concentration of exosomes in the circulation increases in cancer patients (e.g., breast, ovarian, oral, and pancreatic cancer)
[15][107]. Compared with the level of total exosomes, exosomes with specific markers are considered to be more accurate or sensitive for the diagnosis of tumor.
3.2. Nucleic Acids in Exosomes
Exosomes carry a large number of nucleic acids, such as mRNA, microRNAs (miRNAs), and long noncoding RNA (lncRNA), which can be used as biomarkers
[16][108]. In addition, nucleic acids in exosomes can be amplified by PCR, guaranteeing the amount needed for detection
[6]. There have been studies isolating exosomes from the plasma of glioma patients before and after treatment with vaccination. Analysis of 24 immunoregulatory genes in exosomes showed that the mRNA levels of four genes, consisting of IL-8, TGF-β, TIMP-1, and ZAP-70, are significantly downregulated after treatment. All of these genes are known to be associated with the clinical outcomes of glioma patients
[17][109]. Specific exosomal mRNAs could also serve as diagnostic biomarkers for different tumors
[18][19][110,111]. In addition, a lot of miRNAs are found to be potential diagnostic and prognostic biomarkers of tumor, such as miR-638 and miR-150-3p, which are downregulated in patients with hepatocellular cancer
[20][21][22][112,113,114]. LncRNA in circulating exosomes have also been reported to be superior diagnostic biomarkers of tumor and be associated with the tumor progress and survival of patients
[23][24][115,116].
3.3. Proteins in Exosomes
Exosomes inherit many origin-specific proteins from patient cells. The protein levels of circulating exosomes are reported to be increased in cancer patients and related to tumor grade, stage, treatment response, and survival rate of patients with different malignancies
[25][26][27][28][29][117,118,119,120,121]. In addition, the contents of specific proteins in exosomes can also provide diagnostic or prognostic information. TGF-β1 in circulating exosomes is found to be biologically active and increased in patients with acute myeloid leukemia
[30][122]. The high expression and high level of phosphorylation of MET, a hepatocyte growth factor receptor, are found to be increased in patients with advanced stages
[6]. The level of circulating exosomal PD-L1 has been proved to predict response with immunotherapy in NSCLC patients
[31][123]. All findings confirm the promise of exosomes as sensitive and specific biomarkers in the diagnosis and treatment of tumor.
3.4. Isolation and Identification of Exosomes
The isolation and purification of exosomes from biological samples are the basis of their detection. Ultracentrifugation, served as the most commonly used method, possesses significant advantages in the extraction of exosomes from a large number of samples. However, their disadvantages are also obvious: the long time consumption, special equipment, limited recovery, and low purity
[32][124]. In addition to ultracentrifugation, a variety of separation methods based on the physical and chemical properties of exosomes have also been developed. The size-based isolation of exosomes includes exclusion chromatography and ultrafiltration. Compared with ultracentrifugation, ultrafiltration preserves the intact structures of exosomes with similar yield and purity
[33][125]. On the other hand, exclusion chromatography guarantees the purity of exosomes with the efficient separation of proteins and lipids
[34][126]. However, it also faces the challenges of low yield, consumed materials, and long time consumption. Precipitation achieves isolation of exosomes by binding with water molecules to accelerate the precipitate of exosomes. It is also trapped by the contamination of proteins and lipids
[35][127]. The separation of exosomes dependent on immunoaffinity possesses the highest specificity with a simple operation. Through antibodies and aptamers targeting surface proteins on exosomes, exosomes with high expression of specific proteins are immobilized on various carriers, such as magnetic and latex beads, and isolated by magnetic and centrifugal forces, respectively
[36][128]. However, the isolated exosomes are only a small group of exosomes, and the binding sites of the targeted proteins may be blocked, which interferes with the function or detection of the proteins. At present, the detection and quantification of exosomes are mainly focused on the proteins in exosomes. Western blotting, flow cytometry, and ELISA are the most commonly used methods to quantify the protein levels in exosomes by virtue of antibodies. All of these methods possess limited detection limits from ng to pg and complex operations that impede the real-time monitoring of exosomes
[37][129]. In recent years, detection methods targeting nucleic acids and lipids carried by exosomes are constantly emerging, which expands the clinical indicators and contributes to the monitoring of the total population of exosomes
[38][130]. New techniques, such as surface plasmon resonance (SPR) and Raman tweezers microspectroscopy (RTM), have also been introduced in the detection of exosomes
[38][39][130,131]. Benefiting from the development of microfluidic chip technology, innovative methods with low demand for sample volume, high recovery, and short time consumption are established
[40][132]. They can complete the simultaneous isolation and detection of exosomes in one chip.
4. Exosomes as Therapeutic Targets
Cells in the TME constantly release exosomes into the surrounding tumor environment and circulation. These exosomes play pivotal roles in tumor immune escape and immune therapeutic resistance in vivo. Therapeutic approaches that rely on modulating the level or function of exosomes may help address the above-mentioned problems
[41][133]. At present, the regulation of exosomes in vivo mainly focuses on inhibiting their secretion or blocking their interaction with cells. Moreover, some studies try to promote the clearance of exosomes from circulation (
Figure 23).
4.1. Inhibition of Biogenesis of Exosomes
The inhibition of exosome biogenesis is an essential step when targeting the regulation of exosome in circulation. Due to the complicated nature of exosome biogenesis, developing inhibitors that effectively and specially block this process remains a challenge. Inhibitors of ESCRT-dependent transportation of exosomes have been found in recent years, including manumycin A, tipifarnib, and sulfisoxazole. Manumycin A mainly diminish the production level of exosomes by inhibiting the Ras/Raf/ERK1/2 pathway and the expression of hnRNPH1 to attenuate exosome biogenesis and secretion. Manumycin A also inhibits the activity of neutral sphingomyelinase 2 (nSMase2). About 50–65% of exosome biogenesis is inhibited by 250 M manumycin A in different cancer cell lines
[42][134]. Tipifarnib reduces the production level of exosomes by inhibiting the phosphorylation of ERK, which is also necessary for tumor growth. An amount of 1 µM of tipifarnib could inhibit exosome biogenesis by 70% in the prostate cancer cell line C4-2B
[43][135]. Sulfisoxazole is an antibacterial drug that performs inhibition on the secretion of exosomes by targeting ESCRT- or Rab-related proteins in breast adenocarcinoma cell lines
[44][136]. In addition, exosome biogenesis inhibition can generally occur through inhibition on the release of exosomes. GW4869 is one of the most widely used noncompetitive inhibitors of nSMase, which is important for the membrane invagination of MVB
[45][137]. In a previous study, the secretion of vesicles with a size range of 100–200 nm was decreased in breast cancer cells treated with GW4869
[46][138].
4.2. Inhibition of Endocytosis of Exosomes by Recipient Cells
Another theoretical approach is to block interactions between cells and exosomes. However, the unclear mechanism of exosome trafficking and its target definition limit this approach. It is pointed out that both exosomes and the recipient cells determine their interactions
[47][139]. Dynasore is a widely used highly efficient noncompetitive inhibitor of GTPase activities
[48][140]. It exerts an inhibitory effect on the production of clathrin-coated endocytic vesicle
[49][141]. Another inhibitor targeting exosome uptake is heparin. The co-localization of heparin and exosomes is found under microscopy
[50][142]. It is found to completely interact with cell-surface heparan sulfate proteoglycans (HSPGs) and interfere with the transfer of brain tumor cell-derived exosomes into recipient cells
[51][143].