Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Cristina Cristofoletti and Version 2 by Conner Chen.
Sézary syndrome (SS) is an aggressive variant of cutaneous t-cell lymphoma characterized by the accumulation of neoplastic CD4+ lymphocytes—the SS cells—mainly in blood, lymph nodes, and skin. The tumor spread pattern of SS makes this lymphoma a unique model of disease that allows a concurrent blood and skin sampling for analysis.
- cutaneous T-cell lymphoma
- Sézary syndrome
1. Introduction
Cutaneous T-cell lymphomas (CTCLs) include a large spectrum of mature T-cell neoplasms characterized by the accumulation of neoplastic CD4+ T lymphocytes in the skin.
The most common subtype of CTCL is mycosis fungoides (MF), which represents 60% of CTCL cases, while the much rarer variant is Sézary syndrome (SS), which accounts for approximately 5% [1]. The majority of patients with an early MF show a skin-restricted infiltration of malignant cells and an indolent course [1], but about 15% of them can progress toward advanced stages characterized by tumors and erythroderma and that show a decreased survival at less than 5 years [2][3][2,3]. On the contrary, SS is a rare and leukemic subtype of CTCL showing, ab initio, the simultaneous presence of neoplastic lymphocytes mainly in the blood, lymph nodes, and skin.
The relationship between MF and SS is still debated. For a long time, SS was considered an aggressive form of MF, but growing data from genome analyses have demonstrated specific chromosomal alterations occurring in MF but not in SS, and vice versa, suggesting that they are two separate clinical entities [4]. These findings are also reinforced by the different transcriptomic profiles and cell surface markers expressed by MF and SS. According to these data, Campbell et al. indicated that MFs arise from skin resident effector memory (EM) T cells, whereas SS arises from central memory (CM) T-lymphocytes [5]. However, the cell of origin of SS and MF is still undetermined.
Despite the great efforts to characterize SS pathogenesis, it remains an incurable disease, with 5-year overall survival (OS) rates of 15% to 40% for stage IVA and of 0% to 15% for stage IVB [2][6][2,6]. To date, most studies have explored the intrinsic molecular features of SS, demonstrating that it is characterized by a complex profile of chromosome aberrations [7][8][9][7,8,9] and a broad range of genes variously affected by somatic copy-number alterations and somatic single-nucleotide variants that are involved predominantly in T-cell activation and apoptosis, activation of NF-kB, JAK/STAT signaling, chromatin remodeling, and DNA damage response [10][11][10,11].
More recently, attention has shifted toward extrinsic mechanisms that impact the pathophysiology of cancer. Indeed, understanding how the tumor microenvironment sustains neoplastic cells has become focal in cancer research [12]. Regarding CTCL, many reports have already described the complex cellular interactions between malignant lymphocytes and skin elements that play a key role in CTCL pathogenesis [13][14][15][16][17][13,14,15,16,17].
Unlike metastatic spread, cutaneous lymphoma dissemination does not reflect tumor progression, but rather a conserved physiological behavior of neoplastic lymphocytes [18]. This characteristic indicates the strong dependence of these cells on the cutaneous niche that supports their proliferation and survival through the release of nutrients and the induction of cellular signals [19][20][21][22][23][19,20,21,22,23]. Tumor is a dynamic disease, and during its progression usually acquires a greater heterogeneity. This implies that the original cancer cells acquire different molecular signatures, generating an intratumor heterogeneity. Such a condition is the greatest cause of failure in cancer therapies because the spatial and temporal genetic and epigenetic diversity and/or plastic gene expression in cancer cells is often associated with the mechanism of drug resistance [24]. A strong contribution to heterogeneity is also made by the tumor microenvironment, which triggers distinct and specific signals to tumor cells. This last aspect assumes a particular importance in CTCL, given the SS cells simultaneously invade the bloodstream and the skin district, where they interact with keratinocytes, as well as bystander stromal and immune cells [25]. The impact of fluid and solid environment on tumor cells adds complexity to the disease and produces intratumor heterogeneity.
2. Blood and Skin Microenvironments Drive the Transcriptional Program and Signaling of SS Cells
A role for the skin microenvironment in SS pathogenesis has long been hypothesized, but its function in vivo is poorly known. Lately, some investigations have analyzed matched blood- and skin-derived SS cells, revealing substantial differences between these two cell subsets. In this section, we discuss the main results obtained so far will be discussed (Figure 1).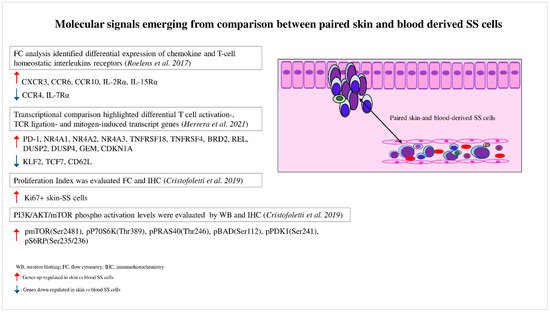
Figure 1. Molecular signals emerging from comparison between paired skin- and blood-derived SS cells. Comparison analyses performed between paired skin- and blood-derived SS cells highlight the molecular signals triggered in these two SS cell subpopulations.
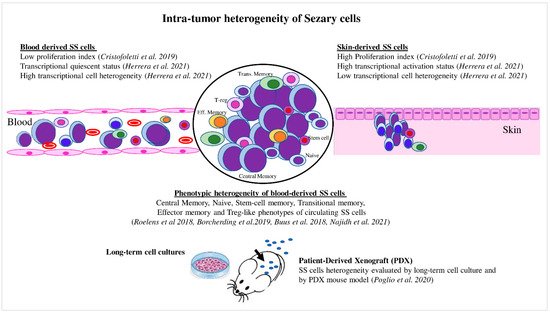
Figure 2. Intratumor heterogeneity of Sézary cells. Skin- and blood-derived SS cells showed differences in proliferation capacity, expression of activation markers, and level of transcriptional heterogeneity. Blood SS cells showed phenotypic heterogeneity, which also was evaluated under specific culture conditions and in the PDX mouse model.