Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Beatrix Zheng and Version 1 by Christian Bailly.
Bladder pathologies, very common in the aged population, have a considerable negative impact on quality of life. Novel targets are needed to design drugs and combinations to treat diseases such as overactive bladder and bladder cancers. A promising new target is the ubiquitous Rho GTPase Rac1, frequently dysregulated and overexpressed in bladder pathologies. Dysregulations of Rac signaling have been reported in atherosclerosis, neurodevelopmental disorders, rheumatic diseases, pulmonary hypertension and different types of cancers, including urothelial carcinoma. Protein Rac1 (RAS-related C3 botulinum toxin substrate (1) is considered a prime target to combat a variety of solid tumors and certain onco-hematological malignancies.
- bladder cancer
- Rho GTPase
- bladder dysfunction
- Rac inhibitors
- metastasis
- overactive bladder
- Rac1 protein
1. Rac1 Structure and Function
Like many other GTPases, Rac1 switches between an inactive GDP-bound and an active GTP-bound state during signal transduction [32][1] (Figure 21c). The protein is involved in a wide range of cellular and physiological processes via a multiplicity of protein partners, among which a variety of guanine nucleotide exchange factors (GEFs) and GTPase-activating proteins (GAPs) essential to control Rac1 activity [33][2]. In addition, a diversity of effector proteins can modulate Rac1 function, such as the serine-threonine kinases PAK1 (p21-activated kinase 1), MLK-1-3 (mixed-lineage kinases), p70 S6 kinase, CaMKII and many other kinases [20][3]. The local availability of GTP in cells plays a role in the control of Rac1 activity [34][4]. The expression and subcellular localization of Rac1 is also regulated at the post-transcriptional level via structural modifications, including phosphorylation, ubiquitination, adenylylation, and lipidation (prenylation, geranylgeranylation, palmitoylation) [35][5]. The lipid anchor is positioned in a hypervariable region, distant from the guanine nucleotide-binding domain, but contributing importantly to the interaction with effectors [36][6]. The protein is generally attached to the plasma membrane, but it can also be found in the nucleus and/or mitochondria [37][7]). At the membrane level, Rac1 can form nanoclusters acting as lipid-based signaling platforms [38][8].
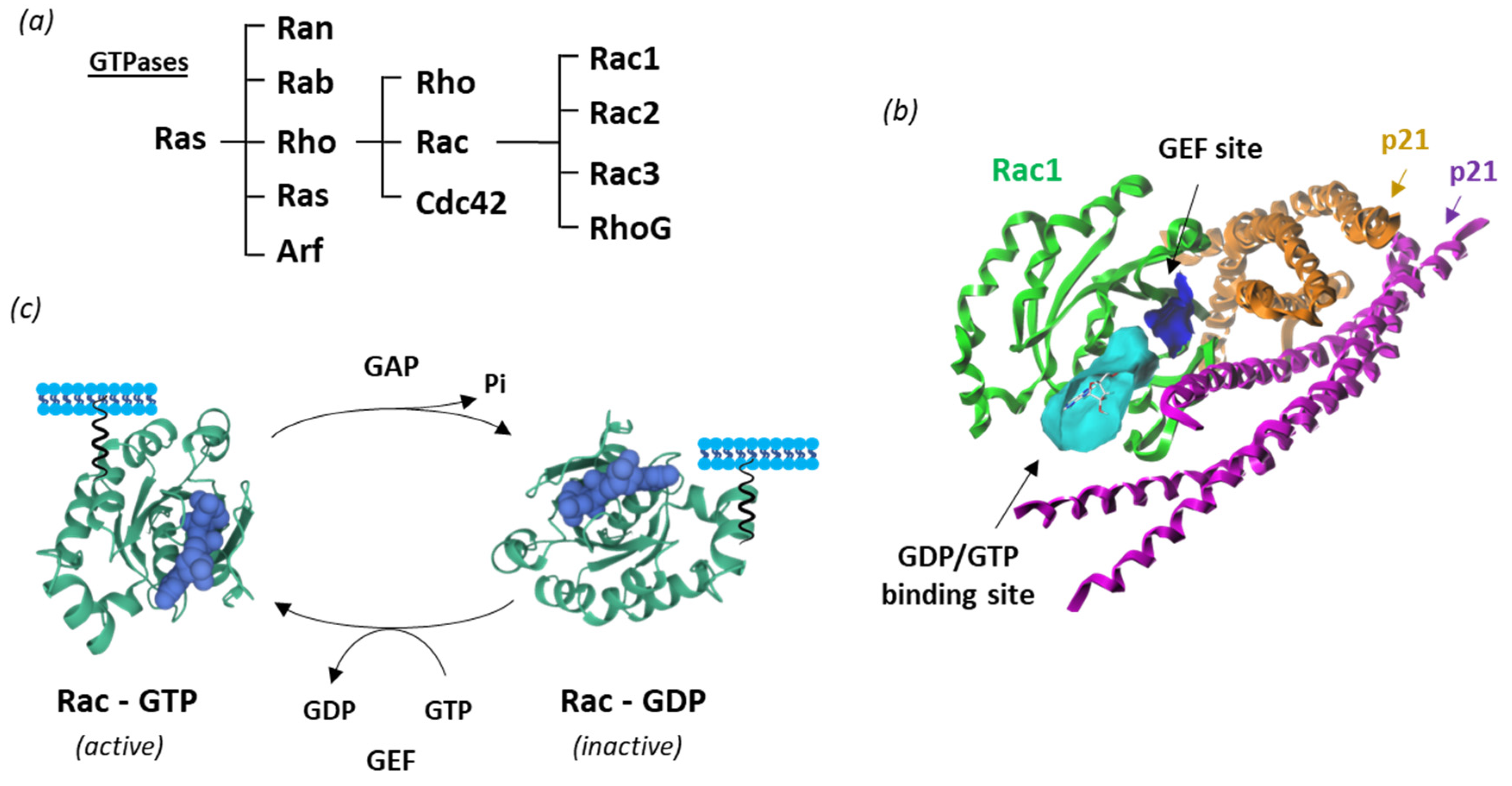
Figure 1. Rac1 structure and activity. (a) Classification of Ras GTPases. (b) A molecular model of Rac1 interacting with p21 (arfaptin) (PDB: 1I4D) with the GTP/GDP (cyan) and GEF (blue) binding sites illustrated [21] (c) Cycle of Rac1 activation. The GTPase cycles between an active GTP-bound state and an inactive GDP-bound state. Guanine nucleotide exchange factors (GEF) turn on signaling by catalyzing the exchange from G-protein-bound GDP to GTP, whereas GTPase activating proteins (GAP) terminate signaling by inducing GTP hydrolysis. GEF and GAP regulate the activity of Rac1 and other small guanine nucleotide-binding (G) proteins to control cellular functions.
The GDP/GTP loading status and cycling rate of Rac1 determine the protein activity. The nucleotide cycling process is impacted by the intrinsic conformational flexibility of the protein and the Mg2+ abundance [39][9]. The level of expression of the protein can vary significantly. An epigenetic downregulation of Rac1 has been reported in patients suffering from depression [40][10]. Conversely, Rac1 is often overexpressed and hyperactivated in cancers, notably in breast, colon, skin (melanoma), liver and lung cancers [31,41,42,43][11][12][13][14]. Moreover, Rac1 gain-of-function mutations have been identified in recent years, such as the two somatic mutations Rac1P29S and Rac1A159V, respectively detected in melanoma and in head-and-neck cancers [44[15][16][17],45,46], and occasionally observed in colon, thyroid, and lung cancers [47][18]. These variants represent fast cycling mutants that contribute to expand tumor phenotypes and confer resistance to targeted therapies. There exists also an alternatively spliced isoform designated Rac1b, with versatile functions, generally involved in tumor progression, but occasionally described as being engaged in the blockade of tumors [48,49][19][20]. To outhe researchers' knowledge, neither the fast-cycling oncogenic mutant enzymes nor the spliced variant Rac1b have been reported in bladder cancer or bladder pathologies.
2. Rac1 in Non-Cancerous Bladder Pathologies
2.1. Rac1 and Bacterial Infections of the Bladder
The ubiquitous Rho GTPase Rac1 plays key roles in the regulation of the cytoskeleton dynamic and cell motility in general. The protein is associated with the formation of protrusions at leading edge of migrating cells (lamellipodia, filopodia), whatever the cell type. As such, Rac1, actively participates to control cellular proliferation and cell mobility. It also plays a role in bacterial attachment to host cells and infections, notably in bladder infections caused by uropathogenic Escherichia coli [50][21]. Urinary tract infection (UTI) produced by uropathogenic E. coli (UPEC) promotes the sensitization of bladder afferent sensory neurons and the virulence factors produced by those bacteria contribute to the sensitization of bladder afferents in UTI [51][22]. The uroepithelial invasion by the bacteria occurs through lipid rafts, and Rac1 associated with caveolin-1 in those rafts is required for the bacterial invasion [52][23]. Rac1 activation enhances the accumulation of actin filaments at sites of bacterial entry (Figure 32). The use of bladder epithelial cells overexpressing constitutively activated Rac1, or conversely, cells with the dominant negative form, has clearly demonstrated that Rac1 activation is essential to the invasion of bladder epithelial cells by type 1 fimbriated E. coli. Moreover, the inhibition of Rac-1 activation via a Toll-like receptor 4 (TLR4)-mediated mechanism was found to suppress bacterial invasion [53][24]. In fact, bacterial lipopolysaccharides engage the TLR4/Rac1/Akt signaling pathway to enter cells and mediate the proliferation of vascular smooth muscle cells [54][25]. Once in the cells, the bacteria produce a toxin CNF1 (cytotoxic necrotizing factor type (1), which constitutively activates different Rho GTPases, including Rac1 critical to phagocytosis, to promote further infection [55][26]. The key role of Rac1 in the invasion of bladder epithelial cells by uropathogenic bacteria suggests that a negative regulation of Rac1 can be an option to reduce and combat infections of the urinary bladder. This can be achieved directly with Rac1-targeting small molecules (discussed below) or indirectly with compounds interfering with Rac1-mediated actin polymerization, as shown with the dietary flavonoid luteolin, for example [56][27].
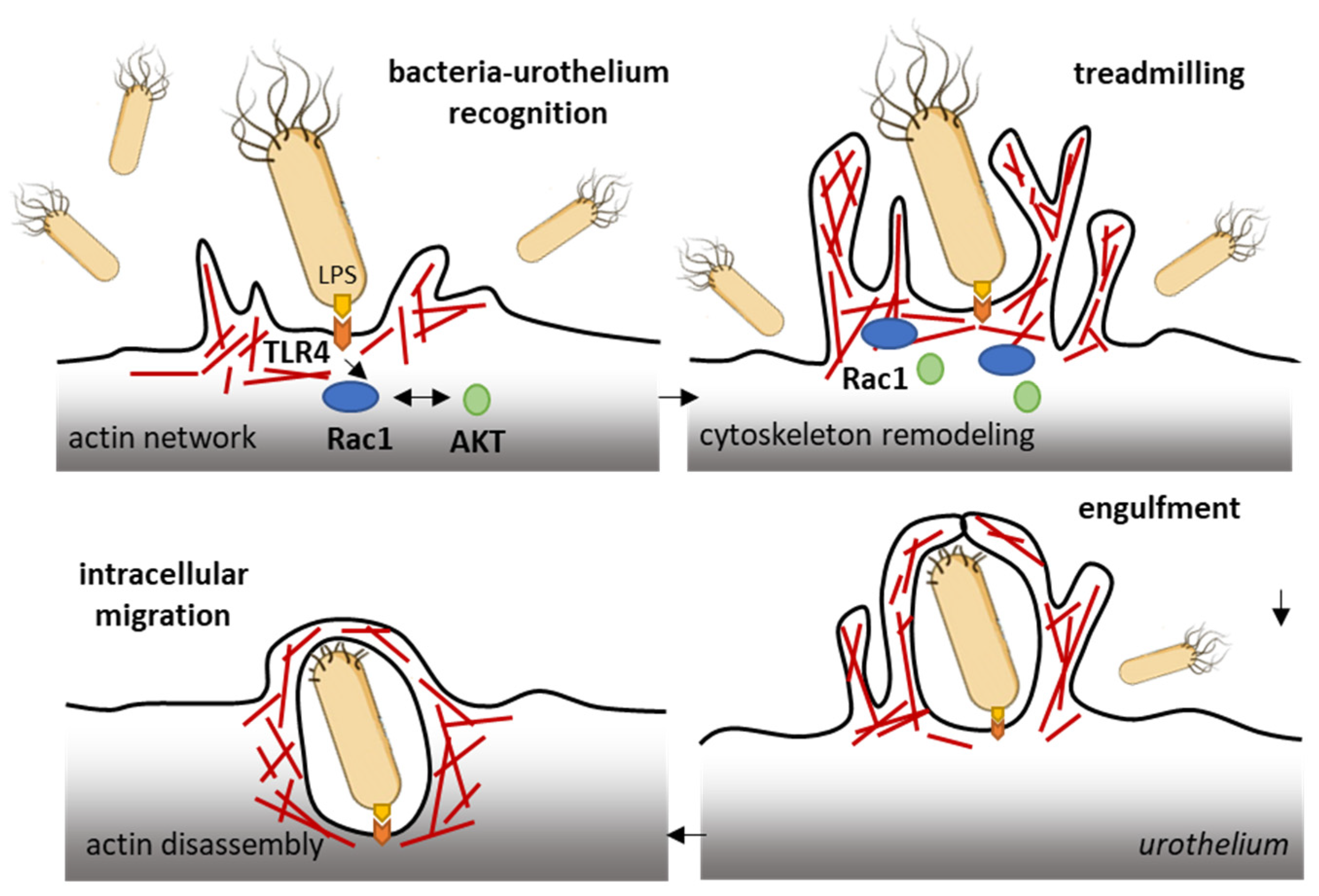
Figure 32. Rac1 plays a role in the invasion of bladder epithelial cells by type 1 fimbriated E. coli. Bacterial lipopolysaccharides activate the TLR4/Rac1/Akt signaling pathway to enter cells vascular smooth muscle cells and colonize the bladder tissue [54][25]. The Rac1 GTPase-mediated contributes to actin cytoskeleton remodeling and regulation of actin filaments.
Rac1 is used by different types of microbial organisms to enter cells. This is also the case for the Mycobacterium bovis Bacille Calmette–Guerin (BCG) strain, which is used as a vaccine for tuberculosis. The BCG infection of primary airway epithelial cells has been shown to induce Rac1 up-regulation and to cause actin redistribution [57][28]. In bladder cancer cells, the entry of the BCG was found to rely on the expression of Rac1 and its effector kinase Pak1 (as well as Cdc42) via a process of micropinocytosis [58][29]. A BCG-induced enhanced expression of Rac1 has been reported in a study with infected macrophages, both in vitro and in vivo. The mycobacteria activated the p38K/JNK/b1-integrin/Rac1 signaling cascade in the frame of the infection [59][30].
The treatment of recurrent urinary tract infections usually relies on the use of intravenous antibiotic therapy (which can lead to complications due to allergy or drug-resistance). Repeated intravesical drug delivery is also possible, but it is more challenging [60][31]. The efficacy is these treatments is suboptimal at present. There is a need for new therapeutic options, new drugs and novel approaches in general to address the pathophysiology of the disease [61][32].
2.2. Rac1 and Diabetes-Induced Bladder Dysfunctions
Urinary bladder dysfunction is a complication in diabetes mellitus (DM) [62][33]. Diabetes causes bladder remodeling leading to uropathy in a mulitfactorial way, with neurogenic and myogenic detrusor overactivity and changes in transmitter regulation leading to a hyper-excitability of the detrusor [63][34]. DM is also a risk factor for bladder cancer prognosis and outcome [64][35]. Diabetic cystopathy (urinary disturbances) is one of the most common complications of diabetes mellitus [65][36]. The pathophysiology of the disease is complex and multifactorial, but it seems clear that Rac1 plays a role in the inflammatory mechanism, via binding to and the activation of the NOD-like receptor protein 3 (NLRP3) inflammasome. Indeed, under hyperglycemia conditions, Rac1 can promote NLRP3 inflammasome activation and induces cell damage [66][37]. The oxidative stress that occurs in the bladder of diabetic subjects causes oxidative damage to the urothelial and smooth muscle cells. A markedly enhanced expression of Pak1 (RAC1/p21 activated kinase 1) has been observed in the smooth muscle of diabetic mouse bladders versus the control group [67][38]. In a rat model of streptozotocin-induced diabetic bladder, an increased expression of Rac1 has been observed by immunohistochemistry. The Rac1 immunoreactivity was found to increase significantly in all the layers of the bladder tissue (epithelium, lamina propria, and tunica muscularis) for the diabetic group compared to the control group [68][39]. This restudyearch is important because it also showed that a Rac1 inhibitor (NSC23766) can inhibit the contractile responses of the bladder detrusor smooth muscle. This pharmacological aspect is discussed further below.
Interestingly, the expression of Rac1 in bladder tissue is increased not only due to the diabetes context, but it is also enhanced mechanically through the induced and cyclic hydrodynamic pressure exerted on bladder smooth muscle cells [69][40]. The expression of both Rac1 and phospho-Rac1 was found to be increased when a hydrodynamic pressure was mechanically applied onto human bladder smooth muscle cells. The expression of Rac1 downstream effectors, such as phospho-MEK1/2 and ERK-1/2, was also increased, and the effects were abrogated when cells were treated with a small molecule Rac1 inhibitor (NSC23766) or a Rac1 siRNA [69][40]. Rac1 seems to play an important role in the proliferation and response of bladder smooth muscle cells to hydrodynamic pressure. The data suggest that, in this situation, the use of Rac1 inhibitors could permit a reduction in bladder dysfunctions.
Another line of evidence showing that Rac1 plays an important role and is required for active contraction in smooth muscle comes from experiments using a conditional Rac1 knockout mouse strain. In this case, the loss of about 50% of Rac1 protein in the urinary bladder resulted in a significant decrease in the contractile responses to different agonists, without causing a remodeling of the vessels in the bladder tissue [70][41]. Similar effects were obtained using Rac1 inhibitors, as discussed below. In a recent study, the silencing of Rac1 expression in human bladder smooth muscle cells was found to reduce cell viability by 50–70% after 48 h and to increase the percentage of cells in (early/late) apoptosis compared to wild-type cells. The effects were associated with alterations in actin organization [71][42].
There are currently multiple pharmacological options to treat diabetes-induced bladder dysfunctions, notably using α1-adrenoceptor and muscarinic receptor antagonists, β3-adrenoceptor agonists and phosphodiesterase type 5 inhibitors [72][43]. However, here also, newer treatments and drugs are needed to improve long-term efficacy.
3. Rac1 in Bladder Cancer
The Rac1 gene, like other Rho-related genes, is frequently overexpressed in urothelial cell carcinoma, and the altered expression of the corresponding proteins plays an important role in the genesis and progression of cancers of the urinary bladder [73,74][44][45]. The gene overexpression and alterations not only concern Rac1 but also the associated regulatory elements, such as kinases PAK1 and PAK4 (P21 activated kinase 1/4), which are amplified and/or overexpressed in muscle-invasive bladder carcinomas [75,76,77][46][47][48]. A moderate or strong positive expression of both Rac1 and PAK1 are considered independent factors for shortened disease-specific survival time in patients with upper urinary tract urothelial carcinoma [78][49]. Numerous studies have reported alterations of Rac1 expression and function in bladder cancer, and the expressed protein has been associated with a variety of functional alterations. For the sake of clarity, wthe researchers can refer to four categories of effects, briefly discussed in turn hereafter.
3.1. Rac1 in Bladder Tumorigenesis
A bioinformatic analysis of mRNA from patients with urothelial carcinoma of the bladder has revealed the presence of a shorter 3’-UTR (3′-untranslated region) isoform of Rac1 and this specific isoform was associated with an upregulation of Rac1 protein expression (Figure 43). The formation of this isoform was apparently mediated by the recruitment of the cleavage stimulation factor 2 (CSTF2) at a polyadenylation site of Rac1, thereby reducing the recruitment of two transcription factors (AFF1 and AFF4), thus causing defects in elongation. The short 3’UTR isoform of Rac1 apparently plays an essential oncogenic role in the pathogenesis of bladder cancer [79][50]. The enhanced expression of Rac1 is certainly not the sole key element contributing to bladder carcinogenesis; the modulation of the full Rho-GTPase axis has been implicated in bladder cancer tumorigenesis [80][51]. Rac1 plays a role in the carcinogenesis of various cancers, including bladder cancer but also hepatocarcinoma, breast cancer, non-small-cell-lung cancers and others [81,82][52][53].
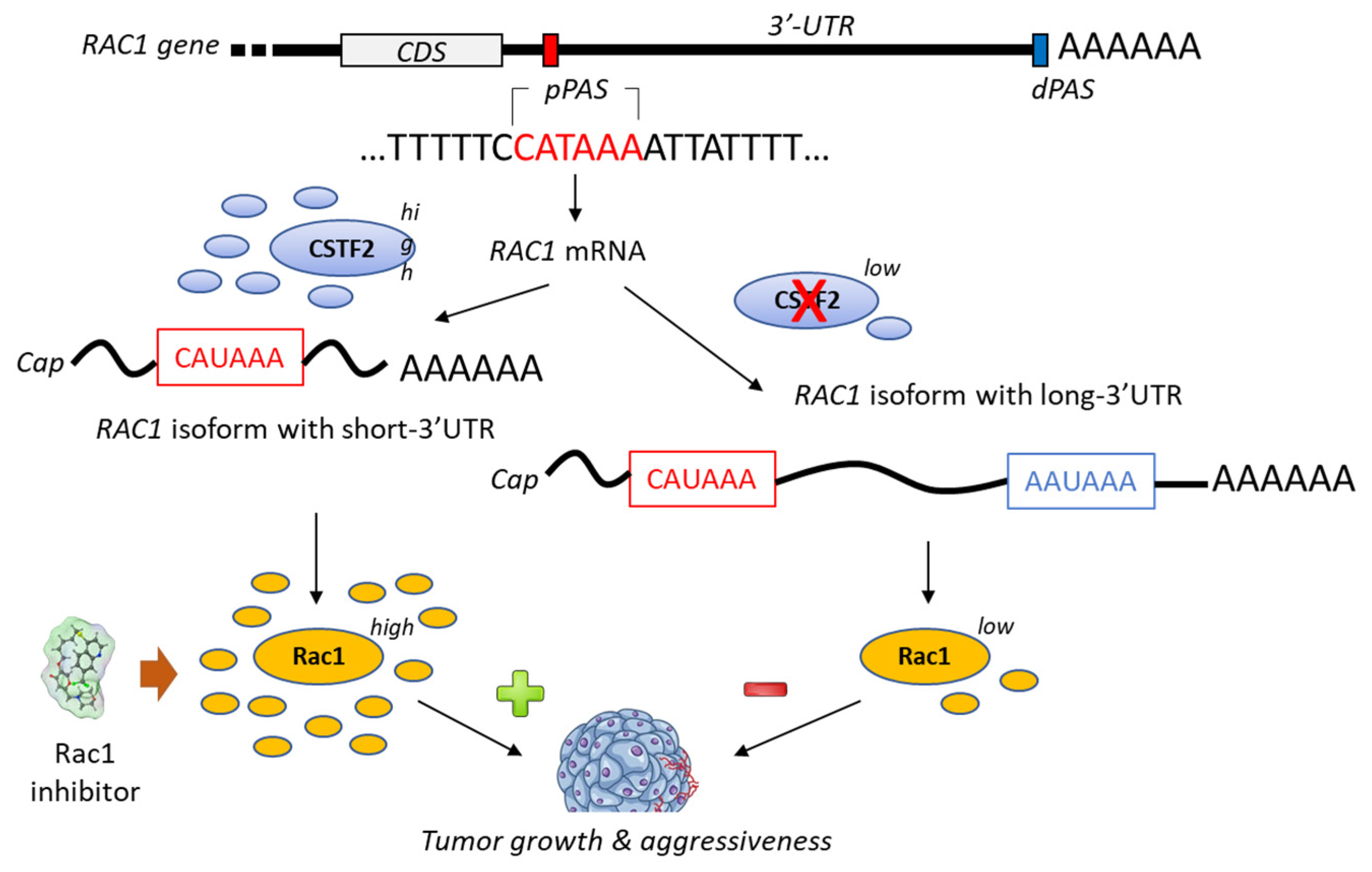
Figure 43. Proposed mechanism for the generation of short-3′-UTR (3′-untranslated region) isoform of Rac1 in the presence of a high level of the cleavage/polyadenylation factor CSTF2 (cleavage stimulation factor (2). The RAC1 short-3′UTR isoform has oncogenic functions and increases aggressiveness of cancer cells from urothelial bladder carcinoma UBC). The dual high expression of CSTF2 and Rac1 with short 3′-UTR predicts worse prognosis for UBC patients [79][50]. The proximal and distal polyadenylation sites (pPAS, dPAS) are located within the terminal exon. CDS, protein-coding sequence.
3.2. Rac1 in Bladder Cancer Cells Proliferation and Tumor Progression
The Rac1 protein is one of the elements that contributes to the proliferation and dissemination of bladder cancer cells. The Rac1 axis is a regulatory mechanism of bladder cancer progression. A recent study has shown that the adaptor protein RacGAP1 inactivated GTP-bound Rac1 in bladder cancer, but the activation was inhibited by protein SHCBP1 (SHC-binding protein 1), which is a regulator of EGF (epidermal growth factor). Via this relay, SHCBP1 can inactivate Rac1 and promote bladder cancer progression [83][54]. The EGF/EGFR ligand/receptor couple is frequently overexpressed in bladder cancers, with squamous bladder cancers qualified as being EGFR-addicted [84][55]. This trend encourages the use of EGFR-targeted drugs to treat these cancers [85,86][56][57]. EGFR signaling generally follows the PAK1/Rac1 route to convey the signal and to regulate tumor progression [87,88][58][59]. EGF is known to stimulate both Rac1 and Pak1 in vascular smooth muscle cells (Figure 54) [89][60]. It is therefore possible to slow down the proliferation and migration of bladder cancer cells via a brake on Rac1. This can be done directly with Rac1 inhibitors or indirectly with molecules capable of controlling Rac1 expression and function. This is the case, for example, for the microRNA miR-142-3p, which interacts directly with Rac1 in bladder cancer cells to inhibit their proliferation but also their migration and invasion [90][61].
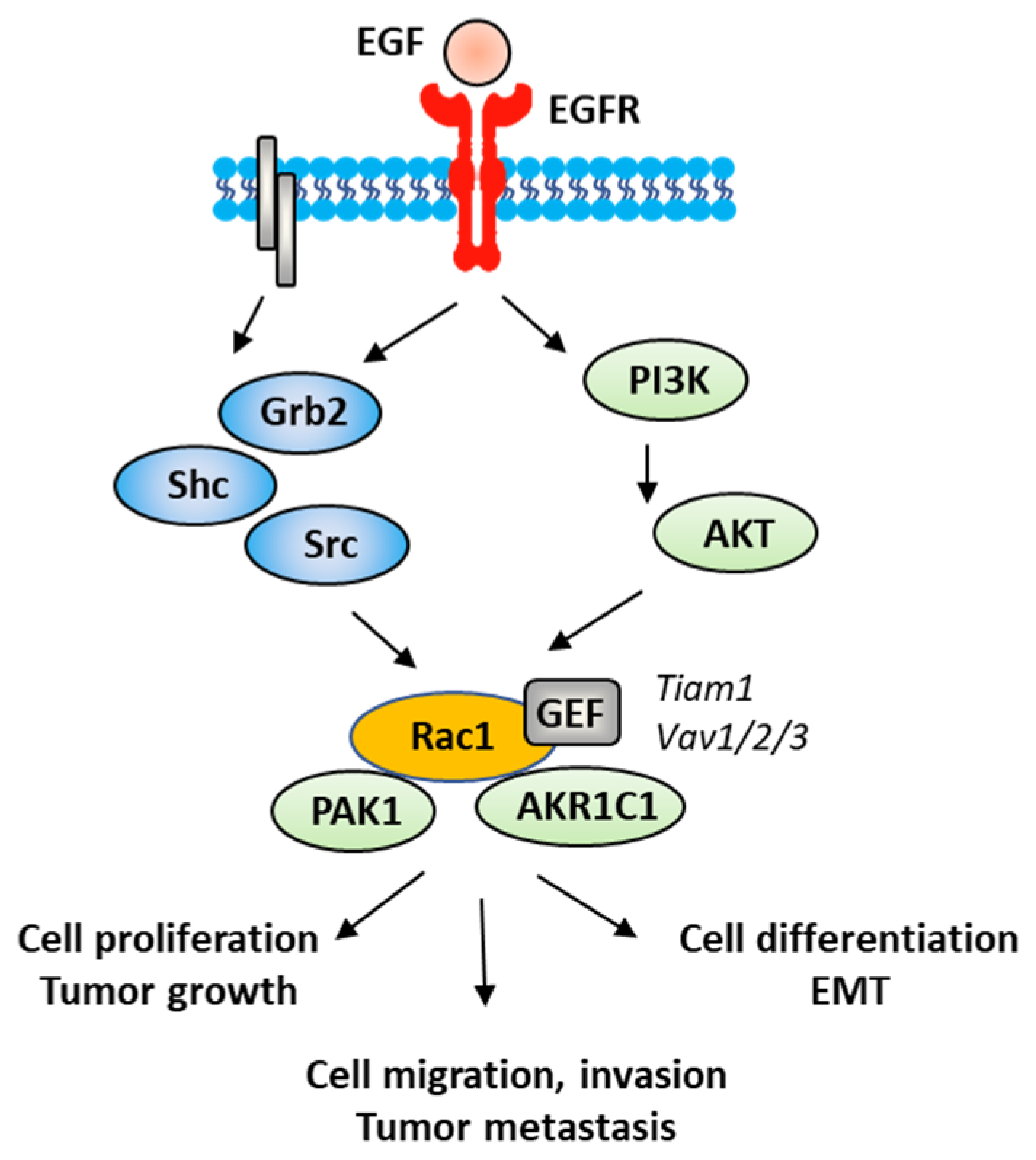
Figure 54.
A signaling network between EGFR and Rac1, in vascular smooth muscle cells with various signaling effectors implicated.
3.3. Rac1 in Epithelial-Mesenchymal Transition (EMT) of Bladder Cancer Cells
EMT is a biological process through which epithelial cells lose their epithelial phenotype and gain mesenchymal features. This process reflects the aggressive and invasive character of the tumor and is often correlated with metastasis. EMT is vital for the progression of bladder cancer tumors because it plays a crucial role in cancer cells spreading and invasion [91][62]. Numerous signaling proteins contribute to this cellular differentiation process and Rac1 is one of them [92][63]. The targeting of the EGFR-Rac1 axis can permit to reverse EMT [93][64]. The activation of Rac1 in the frame of the EMT implicates various modulating proteins, such as SPAG9 (sperm-associated antigen 9) and HEF1 (human enhancer of filamentation 1), which are both connected to Rac1 expression [94][65]. Other factors are implicated in Rac1-mediated EMT, such as the metabolic enzyme AKR1C1 (aldo-keto reductase 1C1), which mediates the invasive potential and drug resistance of metastatic bladder cancer cells (Figure 54). The inhibition of AKR1C1 reduces the invasion/metastatic potential of bladder cancer cells via the regulation of the Rac1/Src/Akt pathway and modulation of the production of inflammatory cytokines, such as interleukin 1β (IL-1β) [95][66]. The link between EMT and Rac1 has been studied more deeply in other types of cancers, notably in lung and colon cancers [96,97][67][68]. The contribution of Rac1 to the inflammation process shall not be neglected. Different studies have pointed out a marked reduction in the production of inflammatory cytokines upon the inhibition of the activation of Rac1, directly or through intermediate effectors [98,99,100][69][70][71]. Rac1 is a major actor of the crosstalk between the inflammatory state and tumor cell migration [101][72]. The expression and activation of Rac1 has frequently been found to enhance the production of pro-inflammatory cytokines (IL-1β, but also IL-6, IL-8 and TNFα) in different pathological situations [102,103][73][74]. For example, the Rac1-GEF interaction inhibitor 1D-142 reduces the nuclear translocation of the transcription factor NFκB induced by the cytokine TNFα in NSCLC cells, and this activity contributes significantly to the antitumor effect of this guanidine-type Rac1 inhibitor in vivo [104][75]. Rac1 can interact directly with specific cytokines, such as IL-37, which controls the membrane translocation of the protein and its signaling activities, at least in lung adenocarcinoma [105][76]. In this context, more attention should be paid to the alternatively spliced isoform Rac1B, the expression of which can be induced by pro-inflammatory extracellular signals in polarized colorectal cancer cells [106,107][77][78]. Rac1 protects cells from undergoing EMT in pancreatic and breast epithelial cells. Similar studies should be conducted with bladder cancer cells.
3.4. Rac1 in Bladder Cancer Metastasis
The role of Rac1 activation in tumor metastasis has been amply discussed, notably in the frame of various solid tumor types [28,46,108][17][79][80]. A comparable situation can be underlined in bladder cancer. Rac1 is a major player of the metastasis of bladder cancer [109][81]. The invasion and migration of bladder cancer cells depend, to some extent, on the activation status of Rac1 and the activity of its regulators, notably the aforementioned Rac1-binding protein aldoketo reductase 1C1 (AKR1C1), up-regulated in metastatic human bladder cancer specimens. AKR1C1 antagonists, such as the anti-inflammatory drug flufenamic acid, can be used to decrease the invasion potential of metastatic bladder cell lines [95][66]. A high activity of GTP-bound Rac1 (coupled with high expression of Pak1) has been measured in the lymph node metastasis of urothelial carcinoma of the upper urinary tract, thus providing a potential prognostic marker for this disease, but also reinforcing the idea that targeting Rac1 can reduce dissemination of the tumor [110][82].
Rac1 plays roles in tumorigenesis, tumor progression, EMT and metastasis of bladder cancer cells. The GTPase has been also implicated in other hallmarks of cancer, notably in stemness, immune escape and drug resistance [30,111][83][84]. These aspects have not been significantly studied in the frame of bladder cancer. For these reasons, wthe researchers will not discuss further these aspects here, but they provide additional indirect lines of evidence supporting the interest of targeting activated Rac1 in bladder cancer.
The management of bladder cancer is excessively complex and variable, depending on the tumor types and stages. Endoscopic transurethral resection of bladder tumor represents the standard of care for non-muscle invasive bladder cancer. However, for more advanced bladder cancers, chemotherapy is required. Immunotherapeutic strategies for bladder cancers have also been largely developed in recent years through the use of immune checkpoint inhibitors (antibodies), adoptive cell therapy, cytokine-based therapy and antibody–drug conjugates [112][85]. However, there is always a need for new drugs and combinations to improve treatment efficacy and patients’ survival.
References
- Kulhanek, K.R.; Roose, J.P.; Rubio, I. Regulation of the Small GTPase Ras and Its Relevance to Human Disease. Methods Mol. Biol. 2021, 2262, 19–43.
- Marei, H.; Malliri, A. GEFs: Dual regulation of Rac1 signaling. Small GTPases 2017, 8, 90–99.
- Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 1831.
- Bianchi-Smiraglia, A.; Wolff, D.W.; Marston, D.J.; Deng, Z.; Han, Z.; Moparthy, S.; Wombacher, R.M.; Mussell, A.L.; Shen, S.; Chen, J.; et al. Regulation of local GTP availability controls RAC1 activity and cell invasion. Nat. Commun. 2021, 12, 6091, Correction in Nat. Commun. 2021, 12, 6482.
- Abdrabou, A.; Wang, Z. Post-Translational Modification and Subcellular Distribution of Rac1: An Update. Cells 2018, 7, 263.
- Lam, B.D.; Hordijk, P.L. The Rac1 hypervariable region in targeting and signaling: A tail of many stories. Small GTPases 2013, 4, 78–89.
- Payapilly, A.; Malliri, A. Compartmentalisation of RAC1 signalling. Curr. Opin. Cell Biol. 2018, 54, 50–56.
- Maxwell, K.N.; Zhou, Y.; Hancock, J.F. Rac1 Nanoscale Organization on the Plasma Membrane Is Driven by Lipid Binding Specificity Encoded in the Membrane Anchor. Mol. Cell. Biol. 2018, 38, e00186-18.
- Killoran, R.C.; Smith, M.J. Conformational resolution of nucleotide cycling and effector interactions for multiple small GTPases determined in parallel. J. Biol. Chem. 2019, 294, 9937–9948.
- Sutton, S.S.; Magagnoli, J.; Cummings, T.; Hardin, J.W.; Love, B.L. Association between thiopurine exposure and depression in patients with inflammatory bowel disease and rheumatoid arthritis. J. Psychopharmacol. 2020, 34, 1163–1167.
- Sauzeau, V.; Beignet, J.; Vergoten, G.; Bailly, C. Overexpressed or hyperactivated Rac1 as a target to treat hepatocellular carcinoma. Pharmacol. Res. 2022, 179, 106220.
- Schnelzer, A.; Prechtel, D.; Knaus, U.; Dehne, K.; Gerhard, M.; Graeff, H.; Harbeck, N.; Schmitt, M.; Lengyel, E. Rac1 in human breast cancer: Overexpression, mutation analysis, and characterization of a new isoform, Rac1b. Oncogene 2000, 19, 3013–3020.
- Casado-Medrano, V.; Baker, M.J.; Lopez-Haber, C.; Cooke, M.; Wang, S.; Caloca, M.J.; Kazanietz, M.G. The role of Rac in tumor susceptibility and disease progression: From biochemistry to the clinic. Biochem. Soc. Trans. 2018, 46, 1003–1012.
- Cannon, A.C.; Uribe-Alvarez, C.; Chernoff, J. RAC1 as a Therapeutic Target in Malignant Melanoma. Trends Cancer 2020, 6, 478–488.
- Uribe-Alvarez, C.; Guerrero-Rodríguez, S.L.; Rhodes, J.; Cannon, A.; Chernoff, J.; Araiza-Olivera, D. Targeting effector pathways in RAC1P29S-driven malignant melanoma. Small GTPases 2021, 12, 273–281.
- Acuner, S.E.; Sumbul, F.; Torun, H.; Haliloglu, T. Oncogenic mutations on Rac1 affect global intrinsic dynamics underlying GTP and PAK1 binding. Biophys. J. 2021, 120, 866–876.
- Colón-Bolea, P.; García-Gómez, R.; Casar, B. RAC1 Activation as a Potential Therapeutic Option in Metastatic Cutaneous Melanoma. Biomolecules 2021, 11, 1554.
- Hodge, R.G.; Schaefer, A.; Howard, S.V.; Der, C.J. RAS and RHO family GTPase mutations in cancer: Twin sons of different mothers? Crit. Rev. Biochem. Mol. Biol. 2020, 55, 386–407.
- Melzer, C.; Hass, R.; Lehnert, H.; Ungefroren, H. RAC1B: A Rho GTPase with Versatile Functions in Malignant Transformation and Tumor Progression. Cells 2019, 8, 21.
- Ungefroren, H.; Wellner, U.F.; Keck, T.; Lehnert, H.; Marquardt, J.U. The Small GTPase RAC1B: A Potent Negative Regulator of-and Useful Tool to Study-TGFbeta Signaling. Cancers 2020, 12, 3475.
- Martinez, J.J.; Hultgren, S.J. Requirement of Rho-family GTPases in the invasion of Type 1-piliated uropathogenic Escherichia coli. Cell. Microbiol. 2002, 4, 19–28.
- Montalbetti, N.; Dalghi, M.G.; Bastacky, S.I.; Clayton, D.R.; Ruiz, W.G.; Apodaca, G.; Carattino, M.D. Bladder infection with uropathogenic Escherichia coli increases the excitability of afferent neurons. Am. J. Physiol. Renal. Physiol. 2022, 322, F1–F13.
- Duncan, M.J.; Li, G.; Shin, J.S.; Carson, J.L.; Abraham, S.N. Bacterial penetration of bladder epithelium through lipid rafts. J. Biol. Chem. 2004, 279, 18944–18951.
- Song, J.; Bishop, B.L.; Li, G.; Duncan, M.J.; Abraham, S.N. TLR4-initiated and cAMP-mediated abrogation of bacterial invasion of the bladder. Cell Host Microbe 2007, 1, 287–298.
- Yin, Q.; Jiang, D.; Li, L.; Yang, Y.; Wu, P.; Luo, Y.; Yang, R.; Li, D. LPS Promotes Vascular Smooth Muscle Cells Proliferation through the TLR4/Rac1/Akt Signalling Pathway. Cell. Physiol. Biochem. 2017, 44, 2189–2200.
- Davis, J.M.; Rasmussen, S.B.; O’Brien, A.D. Cytotoxic necrotizing factor type 1 production by uropathogenic Escherichia coli modulates polymorphonuclear leukocyte function. Infect. Immun. 2005, 73, 5301–5310.
- Shen, X.F.; Teng, Y.; Sha, K.H.; Wang, X.Y.; Yang, X.L.; Guo, X.J.; Ren, L.B.; Wang, X.Y.; Li, J.; Huang, N. Dietary flavonoid luteolin attenuates uropathogenic Escherichia. Coli invasion of the urinary bladder. Biofactors 2016, 42, 674–685.
- Alaridah, N.; Lutay, N.; Tenland, E.; Rönnholm, A.; Hallgren, O.; Puthia, M.; Westergren-Thorsson, G.; Godaly, G. Mycobacteria Manipulate G-Protein-Coupled Receptors to Increase Mucosal Rac1 Expression in the Lungs. J. Innate Immun. 2017, 9, 318–329.
- Redelman-Sidi, G.; Iyer, G.; Solit, D.B.; Glickman, M.S. Oncogenic activation of Pak1-dependent pathway of macropinocytosis determines BCG entry into bladder cancer cells. Cancer Res. 2013, 73, 1156–1167.
- Wu, Y.; Li, C.; Riehle, A.; Pollmeier, B.; Gulbins, E.; Grassmé, H. Mycobacterial Infection is Promoted by Neutral Sphingomyelinase 2 Regulating a Signaling Cascade Leading to Activation of β1-Integrin. Cell. Physiol. Biochem. 2018, 51, 1815–1829.
- Palugan, L.; Cerea, M.; Cirilli, M.; Moutaharrik, S.; Maroni, A.; Zema, L.; Melocchi, A.; Uboldi, M.; Filippin, I.; Foppoli, A.; et al. Intravesical drug delivery approaches for improved therapy of urinary bladder diseases. Int. J. Pharm. X 2021, 3, 100100.
- Josephs-Spaulding, J.; Krogh, T.J.; Rettig, H.C.; Lyng, M.; Chkonia, M.; Waschina, S.; Graspeuntner, S.; Rupp, J.; Møller-Jensen, J.; Kaleta, C. Recurrent Urinary Tract Infections: Unraveling the Complicated Environment of Uncomplicated rUTIs. Front. Cell. Infect. Microbiol. 2021, 11, 562525.
- Bolgeo, T.; Maconi, A.; Bertolotti, M.; Roveta, A.; Betti, M.; Gatti, D.; Boccafoschi, C. Physiopathology of the diabetic bladder. Arch. Ital. Urol. Androl. 2020, 92, 314–317.
- Kirschner-Hermanns, R.; Daneshgari, F.; Vahabi, B.; Birder, L.; Oelke, M.; Chacko, S. Does diabetes mellitus-induced bladder remodeling affect lower urinary tract function? ICI-RS 2011. Neurourol. Urodyn. 2012, 31, 359–364.
- Lu, Y.; Tao, J. Diabetes Mellitus and Obesity as Risk Factors for Bladder Cancer Prognosis: A Systematic Review and Meta-Analysis. Front. Endocrinol. (Lausanne) 2021, 12, 699732.
- Yuan, Z.; Tang, Z.; He, C.; Tang, W. Diabetic cystopathy: A review. J. Diabetes. 2015, 7, 442–447, Erratum in J. Diabetes. 2016, 8, 170.
- Ying, C.; Zhou, Z.; Dai, J.; Wang, M.; Xiang, J.; Sun, D.; Zhou, X. Activation of the NLRP3 inflammasome by RAC1 mediates a new mechanism in diabetic nephropathy. Inflamm. Res. 2022, 71, 191–204.
- Poladia, D.P.; Bauer, J.A. Oxidant driven signaling pathways during diabetes: Role of Rac1 and modulation of protein kinase activity in mouse urinary bladder. Biochimie 2004, 86, 543–551.
- Evcim, A.S.; Micili, S.C.; Karaman, M.; Erbil, G.; Guneli, E.; Gidener, S.; Gumustekin, M. The Role of Rac1 on Carbachol-induced Contractile Activity in Detrusor Smooth Muscle from Streptozotocin-induced Diabetic Rats. Basic Clin. Pharmacol. Toxicol. 2015, 116, 476–484.
- Wu, T.; Chen, L.; Wei, T.; Wang, Y.; Xu, F.; Wang, K. Effect of cyclic hydrodynamic pressure-induced proliferation of human bladder smooth muscle through Ras-related C3 botulinum toxin substrate 1, mitogen-activated protein kinase kinase 1/2 and extracellular regulated protein kinases 1/2. Int. J. Urol. 2012, 19, 867–874.
- Rahman, A.; Davis, B.; Lövdahl, C.; Hanumaiah, V.T.; Feil, R.; Brakebusch, C.; Arner, A. The small GTPase Rac1 is required for smooth muscle contraction. J. Physiol. 2014, 592, 915–926.
- Wang, R.; Yu, Q.; Wang, X.; Li, B.; Ciotkowska, A.; Rutz, B.; Wang, Y.; Stief, C.G.; Hennenberg, M. Rac1 silencing, NSC23766 and EHT1864 reduce growth and actin organization of bladder smooth muscle cells. Life Sci. 2020, 261, 118468.
- Erdogan, B.R.; Liu, G.; Arioglu-Inan, E.; Michel, M.C. Established and emerging treatments for diabetes-associated lower urinary tract dysfunction. Naunyn-Schmiedebergs Arch. Pharmacol. 2022. Online ahead of print.
- Volanis, D.; Zaravinos, A.; Kadiyska, T.; Delakas, D.; Zoumpourlis, V.; Spandidos, D.A. Expression profile of Rho kinases in urinary bladder cancer. J. BUON 2011, 16, 511–521.
- Brait, M.; Munari, E.; LeBron, C.; Noordhuis, M.G.; Begum, S.; Michailidi, C.; Gonzalez-Roibon, N.; Maldonado, L.; Sen, T.; Guerrero-Preston, R.; et al. Genome-wide methylation profiling and the PI3K-AKT pathway analysis associated with smoking in urothelial cell carcinoma. Cell Cycle. 2013, 12, 1058–1070.
- Huang, K.; Chen, G.; Luo, J.; Zhang, Y.; Xu, G. Clinicopathological and cellular signature of PAK1 in human bladder cancer. Tumour Biol. 2015, 36, 2359–2368.
- Huang, K.; Chen, G.; Li, Y.; Liu, J.K.; Wang, Z.Y.; Zhou, G.C. . Zhonghua Yi Xue Za Zhi 2016, 96, 3227–3231.
- Chandrashekar, D.S.; Chakravarthi, B.V.S.K.; Robinson, A.D.; Anderson, J.C.; Agarwal, S.; Balasubramanya, S.A.H.; Eich, M.L.; Bajpai, A.K.; Davuluri, S.; Guru, M.S.; et al. Therapeutically actionable PAK4 is amplified, overexpressed, and involved in bladder cancer progression. Oncogene 2020, 39, 4077–4091.
- Kuroda, K.; Asano, T.; Horiguchi, A.; Ito, K. Effect of increased expression of both ras-related C3 botulinum toxin substrsate 1 and p21-activated kinase 1 in patients with N0M0 upper urinary tract urothelial carcinoma and cancer-free surgical margins. Jpn. J. Clin. Oncol. 2020, 50, 465–472.
- Chen, X.; Zhang, J.X.; Luo, J.H.; Wu, S.; Yuan, G.J.; Ma, N.F.; Feng, Y.; Cai, M.Y.; Chen, R.X.; Lu, J.; et al. CSTF2-Induced Shortening of the RAC1 3’UTR Promotes the Pathogenesis of Urothelial Carcinoma of the Bladder. Cancer Res. 2018, 78, 5848–5862.
- Hu, X.; Xiang, L.; He, D.; Zhu, R.; Fang, J.; Wang, Z.; Cao, K. The long noncoding RNA KTN1-AS1 promotes bladder cancer tumorigenesis via KTN1 cis-activation and the consequent initiation of Rho GTPase-mediated signaling. Clin. Sci. 2021, 135, 555–574.
- De Conti, A.; Tryndyak, V.; Heidor, R.; Jimenez, L.; Moreno, F.S.; Beland, F.A.; Rusyn, I.; Pogribny, I.P. Butyrate-containing structured lipids inhibit RAC1 and epithelial-to-mesenchymal transition markers: A chemopreventive mechanism against hepatocarcinogenesis. J. Nutr. Biochem. 2020, 86, 108496.
- Yue, X.; Wu, F.; Li, Y.; Liu, J.; Boateng, M.; Mandava, K.; Zhang, C.; Feng, Z.; Gao, J.; Hu, W. Gain of function mutant p53 protein activates AKT through the Rac1 signaling to promote tumorigenesis. Cell Cycle 2020, 19, 1338–1351.
- Yin, H.; Zhang, C.; Wei, Z.; He, W.; Xu, N.; Xu, Y.; Li, T.; Ren, K.; Kuang, Y.; Zhu, X.; et al. EGF-induced nuclear translocation of SHCBP1 promotes bladder cancer progression through inhibiting RACGAP1-mediated RAC1 inactivation. Cell Death Dis. 2022, 13, 39.
- Rose, M.; Maurer, A.; Wirtz, J.; Bleilevens, A.; Waldmann, T.; Wenz, M.; Eyll, M.; Geelvink, M.; Gereitzig, M.; Rüchel, N.; et al. EGFR activity addiction facilitates anti-ERBB based combination treatment of squamous bladder cancer. Oncogene 2020, 39, 6856–6870, Correction in Oncogene 2021, 40, 1390.
- Railkar, R.; Krane, L.S.; Li, Q.Q.; Sanford, T.; Siddiqui, M.R.; Haines, D.; Vourganti, S.; Brancato, S.J.; Choyke, P.L.; Kobayashi, H.; et al. Epidermal Growth Factor Receptor (EGFR)-targeted Photoimmunotherapy (PIT) for the Treatment of EGFR-expressing Bladder Cancer. Mol. Cancer Ther. 2017, 16, 2201–2214.
- Nagaya, T.; Okuyama, S.; Ogata, F.; Maruoka, Y.; Knapp, D.W.; Karagiannis, S.N.; Fazekas-Singer, J.; Choyke, P.L.; LeBlanc, A.K.; Jensen-Jarolim, E.; et al. Near infrared photoimmunotherapy targeting bladder cancer with a canine anti-epidermal growth factor receptor (EGFR) antibody. Oncotarget 2018, 9, 19026–19038.
- Lee, S.W.; Commisso, C. Rac1 and EGFR cooperate to activate Pak in response to nutrient stress. Biochem. Biophys. Res. Commun. 2020, 533, 437–441.
- Yao, D.; Li, C.; Rajoka, M.S.R.; He, Z.; Huang, J.; Wang, J.; Zhang, J. P21-Activated Kinase 1: Emerging biological functions and potential therapeutic targets in Cancer. Theranostics 2020, 10, 9741–9766.
- Beier, I.; Düsing, R.; Vetter, H.; Schmitz, U. Epidermal growth factor stimulates Rac1 and p21-activated kinase in vascular smooth muscle cells. Atherosclerosis 2008, 196, 92–97.
- Li, W.Q.; Zhao, W.C.; Xin, J.; Niu, T.L.; Chao, Y.F.; Zhou, P.; Zheng, M.H.; Xu, B. MicroRNA-142-3p suppresses cell proliferation and migration in bladder cancer via Rac1. J. Biol. Regul. Homeost. Agents 2020, 34. Online ahead of print.
- Ashrafizadeh, M.; Hushmandi, K.; Hashemi, M.; Akbari, M.E.; Kubatka, P.; Raei, M.; Koklesova, L.; Shahinozzaman, M.; Mohammadinejad, R.; Najafi, M.; et al. Role of microRNA/Epithelial-to-Mesenchymal Transition Axis in the Metastasis of Bladder Cancer. Biomolecules. 2020, 10, 1159.
- Crosas-Molist, E.; Samain, R.; Kohlhammer, L.; Orgaz, J.L.; George, S.L.; Maiques, O.; Barcelo, J.; Sanz-Moreno, V. Rho GTPase signaling in cancer progression and dissemination. Physiol. Rev. 2022, 102, 455–510.
- Kang, J.; Zhong, Y.; Tian, W.; Li, J.; Li, X.; Zhai, L.; Hou, H.; Li, D. A novel anthraquinone-quinazoline hybrid 7B blocks breast cancer metastasis and EMT via targeting EGFR and Rac1. Int. J. Oncol. 2021, 58, 19.
- Li, X.; Jiang, F.; Wang, X.; Gu, X. SPAG9 regulates HEF1 expression and drives EMT in bladder transitional cell carcinoma via rac1 signaling pathway. Am. J. Cancer Res. 2018, 8, 2467–2480.
- Matsumoto, R.; Tsuda, M.; Yoshida, K.; Tanino, M.; Kimura, T.; Nishihara, H.; Abe, T.; Shinohara, N.; Nonomura, K.; Tanaka, S. Aldo-keto reductase 1C1 induced by interleukin-1beta mediates the invasive potential and drug resistance of metastatic bladder cancer cells. Sci. Rep. 2016, 6, 34625.
- Xia, L.; Lin, J.; Su, J.; Oyang, L.; Wang, H.; Tan, S.; Tang, Y.; Chen, X.; Liu, W.; Luo, X.; et al. Diallyl disulfide inhibits colon cancer metastasis by suppressing Rac1-mediated epithelial-mesenchymal transition. Onco Targets Ther. 2019, 12, 5713–5728.
- Tan, S.; Yi, P.; Wang, H.; Xia, L.; Han, Y.; Wang, H.; Zeng, B.; Tang, L.; Pan, Q.; Tian, Y.; et al. RAC1 Involves in the Radioresistance by Mediating Epithelial-Mesenchymal Transition in Lung Cancer. Front. Oncol. 2020, 10, 649, Erratum in Front. Oncol. 2020, 10, 1106.
- Kim, S.; Jang, Y.W.; Ku, Y.A.; Shin, Y.; Rahman, M.M.; Chung, M.H.; Kim, Y.H.; Kim, D.H. Investigating the Anti-Inflammatory Effects of RCI001 for Treating Ocular Surface Diseases: Insight Into the Mechanism of Action. Front. Immunol. 2022, 13, 850287.
- Liu, H.; Wang, W.; Shen, W.; Wang, L.; Zuo, Y. ARHGAP24 ameliorates inflammatory response through inactivating Rac1/Akt/NF-κB pathway in acute pneumonia model of rat. Ann. Transl. Med. 2020, 8, 1289.
- Shin, B.; Kupferman, J.; Schmidt, E.; Polleux, F.; Delany, A.M.; Lee, S.K. Rac1 Inhibition Via Srgap2 Restrains Inflammatory Osteoclastogenesis and Limits the Clastokine, SLIT3. J. Bone Miner. Res. 2020, 35, 789–800.
- Alves, A.; Diel, L.; Ramos, G.; Pinto, A.; Bernardi, L.; Yates, J.; Lamers, M. Tumor microenvironment and Oral Squamous Cell Carcinoma: A crosstalk between the inflammatory state and tumor cell migration. Oral Oncol. 2021, 112, 105038.
- Ibrahim, J.N.; Jounblat, R.; Jalkh, N.; Abou Ghoch, J.; Al Hageh, C.; Chouery, E.; Mégarbané, A.; Lecron, J.C.; Medlej-Hashim, M. RAC1 expression and role in IL-1β production and oxidative stress generation in familial Mediterranean fever (FMF) patients. Eur. Cytokine Netw. 2018, 29, 127–135.
- Shi, H.; Li, S.; Geng, Y.; Fan, H.; Zhang, R.; Zhang, Y.; Pan, J.; Song, G.; Ge, L.; Xie, T.; et al. Euphorbia factor L3 ameliorates rheumatoid arthritis by suppressing the inflammatory response by targeting Rac family small GTPase 1. Bioengineered 2022, 13, 10984–10997.
- Ciarlantini, M.S.; Barquero, A.; Bayo, J.; Wetzler, D.; Dodes Traian, M.M.; Bucci, H.A.; Fiore, E.J.; Gandolfi Donadío, L.; Defelipe, L.; Turjanski, A.; et al. Development of an Improved Guanidine-Based Rac1 Inhibitor with in vivo Activity against Non-Small Cell Lung Cancer. Chem. Med. Chem. 2021, 16, 1011–1021.
- Li, Y.; Zhao, M.; Guo, C.; Chu, H.; Li, W.; Chen, X.; Wang, X.; Li, Y.; Jia, Y.; Koussatidjoa, S.; et al. Intracellular mature IL-37 suppresses tumor metastasis via inhibiting Rac1 activation. Oncogene 2018, 37, 1095–1106.
- Pereira, J.F.S.; Bessa, C.; Matos, P.; Jordan, P. Pro-Inflammatory Cytokines Trigger the Overexpression of Tumour-Related Splice Variant RAC1B in Polarized Colorectal Cells. Cancers 2022, 14, 1393.
- Zinn, R.; Otterbein, H.; Lehnert, H.; Ungefroren, H. RAC1B: A Guardian of the Epithelial Phenotype and Protector against Epithelial-Mesenchymal Transition. Cells 2019, 8, 1569.
- De, P.; Aske, J.C.; Dey, N. RAC1 Takes the Lead in Solid Tumors. Cells 2019, 8, 382.
- Kotelevets, L.; Chastre, E. Rac1 Signaling: From Intestinal Homeostasis to Colorectal Cancer Metastasis. Cancers 2020, 12, 665.
- Liu, L.; Cui, J.; Zhao, Y.; Liu, X.; Chen, L.; Xia, Y.; Wang, Y.; Chen, S.; Sun, S.; Shi, B.; et al. KDM6A-ARHGDIB axis blocks metastasis of bladder cancer by inhibiting Rac1. Mol. Cancer 2021, 20, 77.
- Kamai, T.; Shirataki, H.; Nakanishi, K.; Furuya, N.; Kambara, T.; Abe, H.; Oyama, T.; Yoshida, K. Increased Rac1 activity and Pak1 overexpression are associated with lymphovascular invasion and lymph node metastasis of upper urinary tract cancer. BMC Cancer 2010, 10, 164.
- Liang, J.; Oyang, L.; Rao, S.; Han, Y.; Luo, X.; Yi, P.; Lin, J.; Xia, L.; Hu, J.; Tan, S.; et al. Rac1, A Potential Target for Tumor Therapy. Front. Oncol. 2021, 11, 674426.
- Zeng, R.J.; Zheng, C.W.; Chen, W.X.; Xu, L.Y.; Li, E.M. Rho GTPases in cancer radiotherapy and metastasis. Cancer Metastasis Rev. 2020, 39, 1245–1262.
- Abd El-Salam, M.A.; Smith, C.E.P.; Pan, C.X. Insights on recent innovations in bladder cancer immunotherapy. Cancer Cytopathol. 2022. Online ahead of print.
More