Medicinal plants have long been a component of human civilization. Plants are utilized to heal ailments and have saved people from countless pandemics, according to Ayurvedic writings and kindred scriptures. Medicinal plants are high in pharmacologically significant bioactive chemicals that define the plant’s medicinal properties. Artemisinin is one such medicinally important metabolite derived principally from the aerial parts of the medicinal herb Artemisia annua (L.). Artemisinin is a sesquiterpene lactone molecule with an endoperoxide structure that has anti-malarial properties. A. annua is a member of the Asteraceae family that grows in tropical and subtropical climates. The plant is native to China and it is known as qinghao, commonly called sweet annie and worm wood in English. The plant is recorded in ancient Chinese texts as being used to cure fevers
- Artemisia annua
- Artemisinin
- bioreactor
- biotechnology
- bioactive compounds
- secondary metabolites
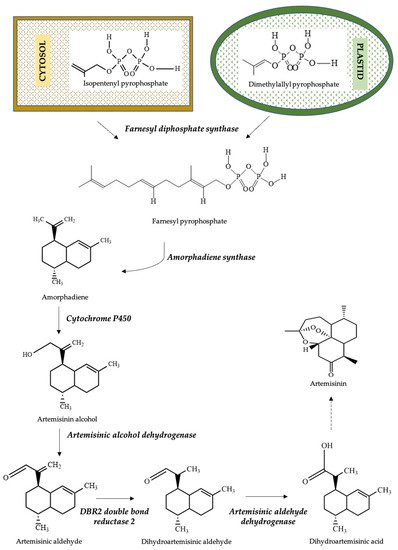
1. Biosynthesis of Artemisinin from Artemisia annua L.
2. In Vitro Production of Artemisinin from Artemisia
2.1. Organ Cultures
2.1.1. Shoot Culture
2.1.2. Hairy Root Culture (Agrobacterium rhizogenes Mediated Transformation)
2.2. Callus and Cell Suspension Cultures
3. Elicitation Strategy for Production of Artemisinin
Plant cells are considered to be biochemical factories for producing high-value secondary metabolites. There are different strategies employed to improve the yield of bioactive compounds for commercial purposes. Elicitation is one of such successful strategies for the production of secondary metabolites. The addition of biotic and abiotic components into the culture medium, with organ cultures and cell suspensions, would aid in the regulation of the metabolism of metabolite production. For artemisinin production, a conventional breeding technique was employed that resulted in new superior breeds, yielding 2% artemisinin DW. A major limitation of this method is the formation of heterozygotes that can show a huge variation in their ability to produce artemisinin. Elicitation of plant cell cultures with physical, chemical and biological agents can drastically improve the yield of metabolites from plants. Furthermore, for its best performance, one has to optimize various influential parameters such as elicitor dose, contact time, growth regulator, age and type of the explant selected and nutritional requirements of the cell [4].3.1. Biotic Elicitation for Artemisinin Production
Endophytic fungi Colletotrichum sp. B501 cell wall-derived oligosaccharide was tested for its elicitation effect on A. annua hairy root cultures for better yield of artemisinin. Hairy root culture in the late growth phase was exposed to elicitor at a concentration of 20 mg/L for four days, which enhanced the yield by 68.29% in comparison to the control. When examined, the peroxidase activity increased in the culture supplemented with elicitor-indicating cells, which were under stress conditions. Furthermore, cellular morphological studies revealed nuclear fragmentation and cell shrinkage leading to programmed cell death. As a part of the plant’s defense strategy, the hairy root culture reportedly produced a more valuable secondary metabolite—artemisinin [28]. Cerebroside C is a natural glycosphingolipid associated with the fungal plant pathogens Fusarium oxysporum, Pythium sps, and Botrytis sps and is reported to be a non-race specific elicitor for plant secondary metabolites. Studies emphasize the significant role of cerebroside as an effective elicitor in the production of artemisinin using hairy root culture. Studies have reported cerebroside to be a novel and potential lipid-based elicitor that can result in a higher accumulation of artemisinin produced by both MAV and MEP pathways. Exposure of cerebroside induced oxidative bursts in the hairy root culture, liberating nitric oxide (NO). This is one of the significant signaling molecules for the biosynthesis of artemisinin. NO inhibitors and NO scavengers when supplemented in media were able to deteriorate artemisinin production [29]. Zhang et al. (2008) [30] studied the potential role of NO in the biosynthesis of artemisinin using a hairy root culture of A. annua root that was elicited with a fungal-derived oligosaccharide. The oligosaccharide isolated from Fusarium oxysporum mycelial biomass was supplemented into the culture media which resulted in the generation of NO and an enhanced accumulation of artemisinin by 20-day-old hairy root culture, after four days of exposure to elicitor in order to confirm the participation of NO as a signaling molecule in the biosynthesis of artemisinin. The culture was treated with sodium nitroprusside (SNP) the NO donor and OE (oligosaccharide elicitor), which has resulted in a maximum yield of the metabolite recorded as 28.5 mg/L—a two-fold increase compared to the culture treated with OE alone. This suggests that generations of endogenous NO in the biosynthesis of artemisinin can trigger with the fungal-derived oligosaccharide elicitor [30]. Colletotrichum sp. is an endophytic fungus living in mutual association with A. annua. Mycelial extract of this fungus was proved to be a potential elicitor of the hairy root culture of A. annua. The 23-day-old late-phase culture was exposed to 0.4 mg total sugar ml−1 of the elicitor for four days, showing a 44% enhancement of artemisinin accumulation. There was an increase in metabolite production with an increase in dose concentration of the elicitor [31]. P. indica, as an endophytic fungus, can colonize on the root and penetrates the root system with its hyphae, further proliferating in the root cortex. Fungi can induce the root cells to produce a precursor for artemisinin production such as artemisinic acid and dihydroartemisinin in roots. These are translocated to shoot where they are transformed into artemisinin [32]. An embryogenic callus obtained from the leaf disc of A. annua was induced to produce regenerated plantlets on MS media with 5.0 mg/L 6-BA and 1.0 mg/L NAA and incubated at 25 ± 1 °C under 16 h photoperiod. Both calluses and callus-regenerated plantlets were exposed to endophytic fungi P. oxalicum B4 to elicit the cultures for artemisinin production. Calluses showed a negative response with browning of tissue, necrosis and no artemisinin, while rooted plantlets after 30 days of exposure to endophytic fungi resulted in a better yield of artemisinin, which was estimated to be 1.32 mg/g DW, about 43.5% more than the control [33]. A. annua shoots derived from nodal explants were cultured on different media such as MS basal media, MS media + 5 μM IBA, MS + P. indica and MS + 5 μM IBA + P. indica. Root regeneration was less in media without P. indica. Artemisinin content was significantly high in shoots established on MS media with 5 μM IBA + P. indica [32].3.2. Abiotic Elicitation for Artemisinin Production
Nano elicitors are gaining more importance in secondary metabolite production using plant cell culture as they can alter physiological activities and biochemical processes inducing the plant cells to make secondary metabolites. Ag-SiO2 NPs were used as an elicitor for hairy root culture for a better yield of artemisinin. NPs sized 101.8 ± 8.9 nm were supplemented into culture media at a concentration of 900 mg/L for three days. This has induced oxidative stress in a hairy root culture due to the gradual release of Ag+ resulting in malonyldialdehyde (MDA), an indicator of lipid oxidation, along with a high generation of hydrogen peroxide and catalase activity. Artemisinin production was estimated to be 13.3 mg/L, which was 3.9-fold better in yield compared to the control [34]. Cell suspension cultures of A. annua were subjected to chitosan nanoparticles of varying concentrations (5–15 mg/L) that were examined for their effect on artemisinin production and expression levels of genes associated with the biosynthesis of artemisinin after 8, 24, 48 and 72 h of treatment. Studies confirmed that 5.0 mg/L was more effective in elicitation with improved yields of artemisinin and upregulated expression of ADS, CYP, CPR, DBR2 and ALDH genes [35]. A callus derived from the leaf disc of A. annua on MS media with 0.5 mg/L NAA, 0.5 mg/L BAP, and 30 g/L sucrose was elicited with cobalt nanoparticles for better artemisinin production. A total of 5.0 mg/L cobalt nanoparticles supplemented into media enhanced artemisinin production after 24 h of elicitation, while its concentration decreased with time. To understand the effect of nanoparticle elicitation on the expression of two key genes involved in biosynthesis of artemisinin, SQS and DBR2, it was analyzed using qRT-PCR. These genes had a negative influence on artemisinin production. Expressions of these genes were low when the callus was elicited with cobalt nanoparticles. On the other hand, HPLC quantification reported an increase in artemisinin yields. The increase in the artemisinin content can be attributed to the activation of other genes involved in the biosynthetic pathway. Furthermore, to have a very clear understanding of the elicited product, it is essential to check the expression of various genes involved in the biosynthesis of the product rather than just studying two or three genes of the biosynthetic pathway [36]. A low artemisinin-producing cell suspension culture of A. annua, sp. was subjected to elicitors, such as ultraviolet B (UV-B) and dimethyl sulfoxide (DMSO), to improve the yield, but elicitation had no effect on the yield of artemisinin. PCR studies revealed the absence of ADS, DBR2 and ALDH1, whose products have a significant role in the biosynthesis of artemisinin [37]. DMSO was used as an abiotic elicitor for in vitro regenerated A. annua shoots with roots. DMSO elicitation was performed on unrooted shoots which did not show any improvement in artemisinin production compared to the rooted shoot that was stimulated by the elicitor. This emphasizes the significant role of the root system in artemisinin production in shoots. Furthermore, DMSO elicitation resulted in the liberation of ROS, which was studied with DAB staining. The rooted plantlets showed an increased DAB stain, indicating ROS is used in shoots as a defense strategy, while unrooted plantlets failed to show any ROS in their leaves. The study has shown that the expression of two gene ADS (amorphadiene synthase) and CRP (amorpha-4,11-diene monooxygenase) involved in the biosynthesis was monitored using RT-PCR. The ADS transcript did not show any response to DMSO elicitation, while CYP transcript was inversely proportional to the artemisinin produced [38]. Native β-cyclodextrins were chemically modified to heptakis (2,6-di-O-methyl)-β-cyclodextrin (DIMEB), which was used as a chemical elicitor in cell suspension culture of A. annua for artemisinin production. The effect on DIMEB and methyl jasmonate was studied separately towards metabolite production. Studies confirmed the combined effect as 50 mM DIMEB and 100 μM methyl jasmonate were optimal with the highest artemisinin of 27 μmol/g dry weight which was 300-fold higher than untreated suspension culture [39]. Oligogalacturonides are damage-associated molecular patterns (DAMPs) released by wounded and injured plant cells. This can serve as a potent elicitor in plants stimulating early defense mechanisms, inducing the plants to produce ROS, which in turn induces plants to produce secondary metabolites [40]. Oligogalacturonides (OGA) are readily obtained from plant tissues using pectinase enzyme, which was further purified using column chromatography. This OGA with polymerization DP = 4.57 when supplemented into a hairy root culture media at a concentration of 60 g/mL for four days increased the yield of artemisinin production to 11.3 mg/L which was 55.2% more than the control without the elicitor [41]. Gamma radiation is an efficient abiotic elicitor that has enhanced the yield of artemisinin in the callus culture of A. annua. Gamma radiation of different doses (5–35 Gy) was used to study its effect on callus growth, cell survival and artemisinin production. The growth and regeneration of calluses are reduced with increasing doses of gamma radiation. To examine its impact on artemisinin production in five successive in vitro subcultures, artemisinin was quantified using HPLC. A low dose of gamma radiation was found to be effective as an elicitor for artemisinin production when compared to a non-irradiated callus culture. This confirms that a low dose of gamma radiation resulted in mutants with enhanced artemisinin production [42]. A low-yielding artemisinin-producing chemotype of the A. annua variety (136 P) was subjected to elicitation with UV-B radiation for different time intervals and varying concentrations of DMSO to stimulate the suspension culture and soil-grown 136 P. There was no elicitation or stimulation in artemisinin production in the case of suspension culture while there was enhanced yield of artemisinin in soil-grown plants [37]. Cell suspension culture of A. annua was set to study the effect of sodium acetate (SA), mevalonic acid lactone (MAL), casein acid hydrolysate (CAH) and cholesterol (CH) as precursors in artemisinin production. A positive response with a better yield of artemisinin was recorded with sodium acetate (25 mg/L), mevalonic acid lactone (50 mg/L) and casein acid hydrolysate (500 mg/L). Feeding the media with sodium acetate resulted in increased acetylco-A in the cell which can boost cellular activities for secondary metabolite production. When the culture was supplemented with mevalonic acid lactone (MAL), it served as the precursor of the lactone ring that enhanced the production of artemisinin. Supplementing casein acid hydrolysate into media will provide aromatic amino acids that can improve the yield of artemisinin. When abiotic elicitor methyl jasmonate was supplemented along with a mevalonic acid lactone (MAL) precursor, it proved to be the best combination as this has resulted in a synergetic effect, with a maximum yield of 110.20 mg/L, which was 5.93 times more than the control [27]. GA3 had a positive effect on the shoot culture of A. annua for artemisinin when supplemented at a concentration of 10 mg/L in MS media. A significant increase in artemisinin was observed at the blooming stage and its effect was only for one cycle, but its effect on artemisinin production ceased in later cycles. In another approach, sterol biosynthesis inhibitors (miconazole, naftifine and terbinafine) were used so that the C15 intermediates are channeled towards artemisinin production. Among the examined inhibitors, naftifine showed an excellent result with its increase in concentration of the product obtained, while terbinafine was toxic to culture and minconazole had a negative impact [13]. This is a study of the synergetic effect of two elicitors on artemisinin production using the cell suspension culture of A. annua. Cells when treated with varying concentrations of sorbitol resulted in osmotic stress in plant cells, which was monitored by estimation of MDA and H2O2 that are considered to be the indicators of oxidative stress. When the culture was pre-treated with coronatine (Cor), it greatly reduced this oxidative stress, as it can stabilize the cell membrane, maintain the integrity of plant cells and also stimulate antioxidant activities to overcome stress. Cell cultures showed a better yield of artemisinin when exposed to 30 g/L of sorbitol. Beyond this, the concentration had a negative impact on artemisinin production due to the ROS-mediated cell disruption. Pre-treatment of cell cultures with Cor also resulted in increasing the yield of artemisinin, but the combined effect drastically increased the yield eight-fold [43]. Shoots generated from the sterilized seeds of A. annua on MS media containing 30 g/L sucrose and 10 g/L agar were maintained at 25 ± 2 °C for 16 h under cool-white light (35 mmol m−2s−1). These shoots were transferred to liquid MS media and subjected to elicitation with silver and copper nanoparticles. Studies confirmed that when nanoparticles were used at concentrations above 10 μM, they inhibited the shoot growth and reduced chlorophyll content. These heavy metal NPs induced oxidative stress liberating more H2O2, which augmented the production of secondary metabolites such as artemisinin as a defense strategy in shoots [44]. The hairy root culture of A. anuua was optimized for increased artemisinin production with gibberellic acid which had a profound effect on its growth. Artemisinin production increases as gibberellic acid induces branching of roots, induction of flowering and accumulation of artemisinin will be always high before flowering [9]. When the culture was elicited with methyl jasmonate (MeJA) and fungal elicitors (Alternaria alternate, Curvularia limata, Fusarium solani, and Piriformospora indica), farnesyl pyrophosphate and miconazole, the results obtained indicated the synergistic effect of abiotic (MeJA at 100 µM) and biotic (CH of P. indica at 3% v/v), elicitors improving the production of artemisinin [45]. The A. annua hairy root culture was subjected to chitosan, methyl jasmonate and yeast extract as the elicitors to stimulate the culture for a better yield of commercially important artemisinin production. Studies confirmed that all these elicitors when supplemented in MS media at concentrations of 200 µM (methyl jasmonate), 2.0 mg/L (Yeast extract) and 150 mg/L (chitosan) were found to be the optimal dose of elicitors for the highest yield of bioactive substances, recorded as 1.52 ± 0.32 and 0.95 ± 0.01 mg/mg dry wt, and 1.84 ± 0.02 mg/mg dry wt., respectively [46]. In order to know the effect and to optimize the concentration of elicitors on biomass and artemisinin production using a hairy root culture of A. annua, RSM studies were performed with 20 experiments designed with three elicitors—namely casein acid hydrolysate, sodium acetate and methyl jasmonate. The steep increase in the yield of artemisinin was recorded as 3.45 mg/g when cultured in MS media with methyl jasmonate (40 µg/L), casein acid hydrolysate (50 µg/L) and sodium acetate (500 µg/L), while in the control, the yield was only 0.4 mg/g [47]. The hairy root culture of A. dubia was transformed with A. rhizogenes LBA9402 that was elicited with gibberellic acid and salicylic acid for artemisinin production. Gibberellic acid exhibited a better elicitation potential at a low concentration of 0.01 mg/L with 80 ± 3 µg/g of DW, which was 93% more than the control. Salicylic acid could also stimulate the culture at a concentration of 13.8 mg/L yielding 79 ± 3 µg/g of artemisinin, which was 38% more than the control [48]. A callus that was initiated from the leaf explant of A. absinthium was cultured on MS media using the raft culture method to investigate the influence of growth factors and amino acids. This culture, when maintained with cysteine amino acid at a concentration of 12.5 mg/L, resulted in 2.8 µg/g of artemisinin, while when media were supplemented with growth factors such as BAP (2 mg/L) and NAA (2 mg/L), it improved the yield of artemisinin to 3.05 µg/g and 1.95 µg/g, respectively. Other combinations of amino acids and growth factors had the least effect on the culture regarding artemisinin production [49]. Table 13 depicts the data of comprehensive research on elicitation of cell and organ cultures using biotic and abiotic agents for artemisinin production.| Elicitor | Culture Type | Culture Conditions |
Yield of Artemisinin |
Reference |
|---|---|---|---|---|
| Biotic elicitors | ||||
| Cell wall’s oligosaccharide from Colletotrichum sp. B501 | Hairy root culture | MS medium + 20 mg/L elicitor | Increased by 68.29% | [28] |
| Cerebroside from fungal source | Hairy root culture | MS medium + 10–70 µg/mL cerebroside | Increased by 2.3 folds | [29] |
| Oligosaccharide from Fusarium oxysporum mycelium | Hairy root culture | MS medium + 0.3 mg total sugar/mL elicitor | Increased from 0.7 mg/g DW to 1.3 mg/g DW | [30] |
| Mycelial extract of Colletotrichum sp. | Hairy root culture | MS medium + 0.4 mg total sugar/mL elicitor | Increased from 0.8 mg/g DW to 1 mg/g DW | [31] |
| Pencillium oxalium B4 | In vitro grown Rooted plantlets | MS medium + 5.0 mg/L BAP + 1.0 mg/L NAA + P. oxalium B4 (30 days exposure) | Increased by 43.5% | [33] |
| Abiotic elicitors | ||||
| Ag-SiO2 nanoparticles | Hairy root culture | MS medium + 900 mg/L nano elicitor | Increased by 3.9 folds | [34] |
| Chitosan nanoparticles | Cell suspension culture | MS medium + 0.5 mg/l NAA + 0.5 mg/L BAP + 15 mg/L Elicitor | NA | [35] |
| Cobalt nano particles | Callus culture | MS medium + 0.5 mg/L NAA + 0.5 mg/L BAP + 5 mg/L elicitor | Increased by 2.25 folds | [36] |
| Chitosan | Hairy root culture | MS medium + 150 mg/L chitosan | Increased by 6 folds | [46] |
| Oligogalacturonides | Hairy root culture | MS medium + 0.01 mg/L gibberellic acid +60 g/mL elicitor | Increased by 55.2% | [41] |
| Heptakis (2,6-di-O-methyl)-β-cyclodextrin (DIMEB) and methyl jasmonate | Cell suspension culture | MS medium + 2.0 mg/L 2,4-D + 0.15 mg/L BAP + 50 mM DIMEB + 100 µM Methyl jasmonate | Increased by 300 folds (27 umol/g DW) | [39] |
| Methyl jasmonate and mevalonic acid lactone | Cell suspension culture | MS medium + 0.1 mg/L NAA + 0.1 mg/L KN + 50 mg/L mevalonic acid lactone + methyl jasmonate | Increased by 5.93 times | [27] |
| Sorbitol and Coronatine | Cell suspension culture | MS medium + 0.1 mg/L NAA + 0.1 mg/L KN + 30 g/L Sorbitol + 0.05 µM Coronatine | Increased by 8 folds | [43] |
4. Bioreactor Scale Production of Artemisinin
Bioreactors are considered to be the heart of industrial biotechnology aiming at the mass production of commercial and pharmaceutical products for humankind. Construction and optimization of the various factors for optimal functioning of the bioreactor are challenging fields of research. The main factors considered in designing a plant bioreactor are multiplication rate, oxygen, and nutrient transfer, reduction in power supply, labor and space. These are generally automated machines that facilitate the controlled operation for a better yield of biomass and metabolites. However, there are limitations that can reduce the performance of bioreactors. The needs to be addressed are the accumulation of inhibitors, foaming, shear stress, contamination and hyperhydricity. By eliminating these limitations, bioreactors can be the ideal tool for mass production of metabolites in a short period of time at a large scale to satisfy the global demand [50]. The work by Patra et al. (2016) [51] focused on improving the growth and artemisinin production from hairy root culture with three different designs of bioreactors, namely bubble column, nutrient mist and modified nutrient mist bioreactors, which were operated in batch mode. Switching from a stirred tank bioreactor to a bubble column bioreactor showed better growth of the hairy root culture when all these bioreactor efficiencies for artemisinin production were evaluated based on the yield of artemisinin and biomass. Among the bioreactor tested, Modified NMB was better in both biomass and artemisinin yield followed by NMB and the column bioreactor. The main reason for better yield with Modified NMB is due to effective oxygen and nutrient supply. In total, 1.12 mg/g of artemisinin with biomass of 23.03 g/L was reported with Modified NMB followed by 0.22 mg/g of artemisinin and 8.52 g/L of biomass with NMB, and the least was with BCR, reporting 0.27 mg/g of artemisinin and biomass of 5.68 g/L [51]. Liu et al. (2003) [52] used a 25-day batch culture of shoots obtained from A. annua to optimize the yield of artemisinin for commercial applications. In order to examine the efficiency of the modified airlift bioreactor, a multi-plate radius-flow bioreactor and an ultrasonic nutrient mist bioreactor shoot culture were initiated and maintained in these bioreactors. The modified airlift bioreactor resulted in the lowest yield of artemisinin as shoots were hyperhydrated leading to vitrification. The mist bioreactor provided excellent support compared to the multiple plate radius bioreactor. Nutrients were provided in the gas phase as a mist, which ensures a good nutrient supply to cultures that improved the yield of artemisinin by 3.3- and 1.4-fold, compared to the airlift and multiple plate radius reactor, respectively. The amount of artemisinin obtained was recorded as 48.2 mg/L after 25 days in the mist bioreactor [52]. The MNB (mist nutrient bioreactor) was found to be the most potential bioreactor for the mass production of artemisinin. Further operating conditions were standardized for improving the yield. A 25-day batch culture was initiated with a shoot culture of A. annua in MNB to ensure homogenous growth of the shoots in the bioreactor. The mist cycle was optimized to 3/90, in which the mist was on for 3 min and off for 90 min, which also resulted in the highest biomass and artemisinin yield, which were 13.4 and 46.9 mg/L [53]. Wyslouzil et al. (2001) [54] standardized the key operating parameters for the highest artemisinin production, using hairy root cultures of A. annua in MNB. Three key factors, namely mist cycle, carrier gas and nutrient composition, that can influence the kinetics of the root and the accumulation of artemisinin, were considered in this experiment. Setting the mist cycle can directly influence the root growth as well as the artemisinin production, as they can distribute the nutrients uniformly in the bioreactor and also mediate oxygen transfer. A 1 min on/15 min off mist cycle resulted in a good branching of the root with a fresh weight of 4.6 ± 1.0 mg, while 5 min on/15 min off caused browning of the root and necrosis with a fresh weight of 3.4 ± 1.4 mg. A 1 min on/60 min off mist cycle resulted in thinner roots with the least branching of roots with a fresh weight of 3.9 ± 0.6 mg. Furthermore, when 1% CO2 was used as the carrier gas, it had no impact on root growth but greatly reduced necrosis. Conditioned B5 media were found to be more effective in maintaining the roots in the log phase for a long time, and branching was also extensive [54]. Patra and Srivastava (2015) [55] used mathematical models to standardize the feed batch culture for the highest yield of artemisinin using a modified stirred tank bioreactor. Sucrose is a significant carbon source in plant tissue culture media and an osmoregulator that can greatly influence the in vitro propagation of roots in a bioreactor. The fed-batch culture can easily overcome the nutrient limiting factor for improving the yield of secondary metabolites. The application of the model design of the fed-batch culture can improve its performance. A modified Monod’s model was employed to optimize the addition of sucrose to the fed-batch culture of the hairy root culture of A. annua for the highest yield of artemisinin. Sucrose (37 g/L) was fed at a constant rate of 0.1 L/day for 10–15 days, which resulted in improved artemisinin accumulation of 0.77 mg/g, while another batch culture, which was maintained in a pseudo-steady state fed with (20.8 g/L) for 10–15 days, resulted in the highest artemisinin accumulation recorded as 0.99 mg/g [55]. Patra and Srivastava (2014) [56] set a batch culture of a hairy root from A. annua to examine and optimize the factors associated with the operation of a stirred tank bioreactor for the high accumulation of artemisinin in the culture. When the culture was set beyond 70 rpm, it resulted in sheer stress. Similarly, other parameters, such as temperature (25 °C), size of inoculum (1 g/L DW), age of inoculum (8 d) and medium to vessel volume ratio (0.18), were optimized for the best performance of the stirred tank bioreactor towards the mass production of artemisinin. A high biomass accumulation of 6.3 g/L dry weight (37.50 g fresh weight) and 0.32 mg/g was obtained after an incubation period of 25 days [56]. A batch culture was initiated in an internal loop bioreactor with an ultrasonic mist cycle for improved growth and artemisinin production from the adventurous roots of A. annua. The mist cycle was set in such a way that every 90 min interval, three mins of misting was applied for effective nutrient supply and oxygen transfer. After 25 days of incubation, with an airflow rate of 0.5 L/min, artemisinin yield was estimated as 46.9 mg DW/L of the culture media [57]. Kim et al. (2001) confirmed that the hairy root culture could yield three times more artemisinin in a nutrient mist bioreactor (NMB) than in a bubble bioreactor [58]. The hairy root culture is one of the recommended cultures for the mass production of plant secondary metabolites due to its genetic stability, good productivity and ease in genetic manipulations. In these studies, bubble bioreactors and modified bubble bioreactors were used to analyze the growth and artemisinin production of the hairy root culture. After 25 days of incubation, improved artemisinin was produced and a modified bubble bioreactor was observed [59]. Table 24 depicts the data of comprehensive research on bioreactors and culture conditions for the production of artemisinin.| Culture Type | Type of Bioreactor | Culture Conditions | Yield | References |
|---|---|---|---|---|
| Hairy root culture | Modified nutrient mist bioreactor | ¼ MS + 10 µg/L GA3Batch culture having adequate oxygen supply and nutrient | 1.12 mg/g | [51] |
| Shoot culture | Mist bioreactor | MS + 0.05 mg/L NAA + 0.5 mg/L BAP; 25 days batch | 48.2 mg/L | [52] |
| Shoot culture | Mist nutrient bioreactor | MS + 0.05 mg/L NAA + 0.5 mg/L BAP; 25 days batch | 46.9 mg/L | [53] |
| Hairy root culture | Mist nutrient bioreactor | B5 medium; 1 min on/15 min off mist cycle | NA | [54] |
| Hairy root culture | Modified stirred tank bioreactor | ¼ MS + 10 µg/L GA3; Fed batch for 10–15 days | 0.99 mg/g DW | [55] |
| Hairy root culture | Stirred tank bioreactor | MS medium; 25-day batch. | 0.32 mg/g DW | [56] |
| Adventitious roots | Mist bioreactor | MS medium; 0.5 L/min air, 25-day culture | 46.9 mg DW/L | [57] |
5. Metabolic Engineering for Artemisinin Production
Conventional and in vitro modes of artemisinin production in plant systems alone can not meet the demand for artemisinin, as biosynthetic studies revealed that their production is restricted to specific tissues such as glandular trichomes. From recent studies, it is known that modern biotechnological applications such as metabolic engineering have helped in deriving the required metabolites from plants and microbes. It can be seen that terpenoids were produced by engineering mevalonate pathways in E. coli. Studies carried out by Martin et al. (2003) showed that the yield of precursors such as amorpha-diene was increased by engineering the amorpha-4,11-diene synthase gene from Saccharomyces cerevisiae [60]. Similarly, there are many studies carried out on plant model systems that helped in the enhancement of artemisinin. Previously, human therapeutic proteins were successfully produced in plant model systems, such as tobacco, using metabolic engineering techniques. The success rate was significant [61][62]. The expression of amorpha-4,11-diene is particularly significant in metabolic engineering for the manufacture of artemisinin and it is produced from the precursor farnesyl diphosphate. Wallaart et al. (2001) [63] successfully cloned the cDNA sequence for the expression of amorpha-4,11-diene synthase in E. coli and they could reach 50% identity. Later, upon introduction to the tobacco plant system, they found that it resulted in significant expression. This transgenic tobacco model system successfully helped in the production of amorpha-4,11-diene (0.2–1.7 ng/g FW) [63]. Amorpha-4,11-diene is expressed more than 1000-fold along with some other novel terpene molecules when cytosolic carbons and plastid’s isoprenoids are overexpressed and diverted among their compartments. Co-expressing plastid-targeted ADS and FPS helped in increasing the yield of amorpha-4,11-diene (25 µg/g FW) in tobacco [64]. Employing synthetic biology, Farhi et al. (2011) tried to create a mega vector with cytochrome P450, ADS, CYP71AV1, DBR2 and a de-regulated tHMGR gene from yeast. In spite of being strategically robust at the molecular level, the yield of artemisinin is comparatively lower (around 7 µg/g DW) [5]. Agroinfiltration of the constructs with amorpha-4,11-diene synthase, 3-hydroxy-3-methylglutaryl-CoA reductase and FPS regions helped in the expression of amorpha-4,11-diene in leaves of Nicotiana benthamiana [65].References
- Weathers, P.J.; Arsenault, P.R.; Covello, P.S.; McMickle, A.; Teoh, K.H.; Reed, D.W. Artemisinin production in Artemisia annua: Studies in planta and results of a novel delivery method for treating malaria and other neglected diseases. Phytochem. Rev. 2010, 10, 173–183.
- Teoh, K.H.; Polichuk, D.R.; Reed, D.W.; Nowak, G.; Covello, P.S. Artemisia annua L. (Asteraceae) trichome-specific cDNAs reveal CYP71AV1, a cytochrome P450 with a key role in the biosynthesis of the antimalarial sesquiterpene lactone artemisinin. FEBS Lett. 2006, 580, 1411–1416.
- Weathers, P.J.; Elkholy, S.F.; Wobbe, K.K. Artemisinin: The biosynthetic pathway and its regulation in Artemisia annua, a terpenoid-rich species. Vitr. Cell. Dev. Biol. -Plant 2006, 42, 309–317.
- Ikram, N.K.B.K.; Simonsen, H.T. A Review of biotechnological artemisinin production in plants. Front. Plant Sci. 2017, 8, 1966.
- Farhi, M.; Marhevka, E.; Ben-Ari, J.; Algamas-Dimantov, A.; Liang, Z.; Zeevi, V.; Edelbaum, O.; Spitzer-Rimon, B.; Abeliovich, H.; Schwartz, B.; et al. Generation of the potent anti-malarial drug artemisinin in tobacco. Nat. Biotechnol. 2011, 29, 1072–1074.
- Xie, D.-Y.; Ma, D.-M.; Judd, R.; Jones, A.L. Artemisinin biosynthesis in Artemisia annua and metabolic engineering: Questions, challenges, and perspectives. Phytochem. Rev. 2016, 15, 1093–1114.
- Blando, F.; Rizzello, F.; Durante, M.; De Paolis, A.; Caretto, S.; Mita, G. In Vitro adventitious regeneration of Artemisia annua L. influencing artemisinin metabolism. Horticulturae 2021, 7, 438.
- Liu, C.; Zhao, Y.; Wang, Y. Artemisinin: Current state and perspectives for biotechnological production of an antimalarial drug. Appl. Microbiol. Biotechnol. 2006, 72, 11–20.
- Liu, C.; Wang, Y.; Ouyang, F.; Ye, H.; Li, G. Production of artemisinin by hairy root cultures of Artemisia annua L in bioreactor. Biotechnol. Lett. 1998, 20, 265–268.
- Weathers, P.J.; Cheetham, R.D.; Follansbee, E.; Teoh, K. Artemisinin production by transformed roots of Artemisia annua. Biotechnol. Lett. 1994, 16, 1281–1286.
- Paniego, N.; Giulietti, A.M. Artemisinin production by Artemisia annua L.-transformed organ cultures. Enzym. Microb. Technol. 1996, 18, 526–530.
- Paniego, N.B.; Giulietti, A.M. Artemisia annua L.: Dedifferentiated and differentiated cultures. Plant Cell Tissue Organ Cult. 1994, 36, 163–168.
- Woerdenbag, H.; Lüers, J.F.J.; Van Uden, W.; Pras, N.; Malingré, T.M.; Alfermann, A.W. Production of the new antimalarial drug artemisinin in shoot cultures of Artemisia annua L. Plant Cell Tissue Organ Cult. 1993, 32, 247–257.
- Gulati, A.; Bharel, S.; Jain, S.K.; Abdin, M.Z.; Srivastava, P.S. In Vitro micropropagation and flowering in Artemisia annua. J. Plant Biochem. Biotechnol. 1996, 5, 31–35.
- Ferreira, J.F.; Janick, J. Immunoquantitative analysis of artemisinin from Artemisia annua using polyclonal antibodies. Phytochemistry 1996, 41, 97–104.
- Martinez, B.C.; Staba, E.J. The Production of artemisinin in Artemisia annua L. tissue cultures. Adv. Cell Culture 1988, 6, 69–87.
- Liu, C.; Guo, C.; Wang, Y.; Ouyang, F. Factors influencing artemisinin production from shoot cultures of Artemisia annua L. World J. Microbiol. Biotechnol. 2003, 19, 535–538.
- Ferreira, J.F.; Janick, J. Roots as an enhancing factor for the production of artemisinin in shoot cultures of Artemisia annua. Plant Cell Tissue Organ Cult. 1996, 44, 211–217.
- Ahlawat, S. Influence of Agrobacterium rhizogenes on induction of hairy roots for enhanced production of artemisinin in Artemisia annua L. plants. Afr. J. Biotechnol. 2012, 11, 35.
- Gantait, S.; Mukherjee, E. Hairy root culture technology: Applications, constraints and prospect. Appl. Microbiol. Biotechnol. 2020, 105, 35–53.
- Beigmohammadi, D.M.; Seyyedi, M.; Rostampour, S.; Mohammadi, E.; Sharafi, A. In vitro regeneration and genetically transformed culture of Artemisia diffusa. J. Appl. Biotechnol. Rep. 2021, 8, 312–319.
- Timina, O.; Timin, O.; Stepanova, A.; Shchuka, T. Getting bearded roots in Artemisia annua L. In Proceedings of the 90 Years—From Plant to Medicine: Achievements and Prospects, Moscow, Russia, 10 June 2021. (in Russian).
- Dilshad, E.; Cusido, R.M.; Estrada, K.R.; Bonfill, M.; Mirza, B. Genetic transformation of Artemisia carvifolia Buch with rol genes enhances artemisinin accumulation. PLoS ONE 2015, 10, e0140266.
- Brown, G. Production of anti-malarial and anti-migraine drugs in tissue culture of Artemisia annua and Tanacetum parthenium. Acta Hortic. 1993, 269–276.
- Woerdenbag, H.J.; Pras, N.; Van Uden, W.; De Boer, A.; Batterman, S.; Visser, J.F.; Malingré, T.M. High peroxidase activity in cell cultures of Artemisia annua with minute artemisinin contents. Nat. Prod. Lett. 1992, 1, 121–128.
- Liu, K.-S.; Yang, S.-L.; Roberts, M.; Elford, B.; Phillipson, J. Antimalarial activity of Artemisia annua flavonoids from whole plants and cell cultures. Plant Cell Rep. 1992, 11, 637–640.
- Baldi, A.; Dixit, V. Yield enhancement strategies for artemisinin production by suspension cultures of Artemisia annua. Bioresour. Technol. 2008, 99, 4609–4614.
- Wen, W.J.; Hao, X.Z.; Xiang, T.R. Elicitation on artemisinin biosynthesis in Artemisia annua hairy roots by the oligosaccharide extract from the Endophytic colletotrichum sp. B501. J. Integr. Plant Biol. 2002, 44, 1233.
- Wang, J.W.; Zheng, L.P.; Zhang, B.; Zou, T. Stimulation of artemisinin synthesis by combined cerebroside and nitric oxide elicitation in Artemisia annua hairy roots. Appl. Microbiol. Biotechnol. 2009, 85, 285–292.
- Zheng, L.-P.; Guo, Y.-T.; Wang, J.-W.; Tan, R.-X. Nitric Oxide potentiates Oligosaccharide-induced artemisinin production in Artemisia annua hairy roots. J. Integr. Plant Biol. 2008, 50, 49–55.
- Wang, J.W.; Zhang, Z.; Tan, R.X. Stimulation of artemisinin production in Artemisia annua hairy roots by the elicitor from the endophytic Colletotrichum sp. Biotechnol. Lett. 2001, 23, 857–860.
- Sharma, G.; Agrawal, V. Marked enhancement in the artemisinin content and biomass productivity in Artemisia annua L. shoots co-cultivated with Piriformospora indica. World J. Microbiol. Biotechnol. 2013, 29, 1133–1138.
- Zheng, L.P.; Tian, H.; Yuan, Y.F.; Wang, J.W. The influence of endophytic Penicillium oxalicum B4 on growth and artemisinin biosynthesis of in vitro propagated plantlets of Artemisia annua L. Plant Growth Regul. 2016, 80, 93–102.
- Zhang, B.; Zheng, L.P.; Yi Li, W.; Wen Wang, J. Stimulation of artemisinin production in Artemisia annua hairy roots by Ag-SiO2 core-shell nanoparticles. Curr. Nanosci. 2013, 9, 363–370.
- Ghassemi, B.; Nayeri, F.D.; Hosseini, R. The effects of chitosan nanoparticles on genes expression of artemisinin synthase in suspension culture of Artemisia annua L.: A comparative study. Int. J. Adv. Biol. Biomed. Research 2021, 9, 190–203.
- Ghasemi, B.; Hosseini, R.; Nayeri, F.D. Effects of cobalt nanoparticles on artemisinin production and gene expression in Artemisia annua. Turk. J. Bot. 2015, 39, 769–777.
- Yee, M.K.; Ping, W.Y. Successes of artemisinin elicitation in low-artemisinin producing Artemisia annua cell cultures constrained by repression of biosynthetic genes. bioRxiv 2019, 740167.
- Mannan, A.; Liu, C.; Arsenault, P.R.; Towler, M.J.; Vail, D.R.; Lorence, A.; Weathers, P.J. DMSO triggers the generation of ROS leading to an increase in artemisinin and dihydroartemisinic acid in Artemisia annua shoot cultures. Plant Cell Rep. 2009, 29, 143–152.
- Durante, M.; Caretto, S.; Quarta, A.; De Paolis, A.; Nisi, R.; Mita, G. β-Cyclodextrins enhance artemisinin production in Artemisia annua suspension cell cultures. Appl. Microbiol. Biotechnol. 2011, 90, 1905–1913.
- Pontiggia, D.; Ciarcianelli, J.; Salvi, G.; Cervone, F.; De Lorenzo, G.; Mattei, B. Sensitive detection and measurement of oligogalacturonides in Arabidopsis. Front. Plant Sci. 2015, 6, 258.
- Zhang, B.; Zou, T.; Lu, Y.H.; Wang, J.W. Stimulation of artemisinin biosynthesis in Artemisia annua hairy roots by oligogalacturonides. Afr. J. Biotechnol. 2010, 9, 3437–3442.
- Patil, A.; Suryavanshi, P.; Fulzele, D. In vitro regeneration of gamma irradiated callus of Artemisia annua and evaluation of increase artemisinin content by HPLC analysis. J. Anal. Pharm. Res. 2018, 7, 1.
- Salehi, M.; Karimzadeh, G.; Naghavi, M.R. Synergistic effect of coronatine and sorbitol on artemisinin production in cell suspension culture of Artemisia annua L. cv. Anamed. Plant Cell Tissue Organ Cult. 2019, 137, 587–597.
- SaghirzadehDarki, B.; Shabani, L.; Pourvaez, R.; Ghannadian, M. Effects of CuSO4 and AgNO3 on artemisinin and phenolic compound in shoot cultures of Artemisia annua L. J. Plant Physiol. 2010, 31, 1–8.
- Ahlawat, S.; Saxena, P.; Alam, P.; Wajid, S.; Abdin, M.Z. Modulation of artemisinin biosynthesis by elicitors, inhibitor, and precursor in hairy root cultures of Artemisia annua L. J. Plant Interact. 2014, 9, 811–824.
- Putalun, W.; Luealon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y. Improvement of artemisinin production by chitosan in hairy root cultures of Artemisia annua L. Biotechnol. Lett. 2007, 29, 1143–1146.
- Patra, N.; Srivastava, A.K.; Sharma, S. Study of various factors for enhancement of artemisinin in Artemisia annua hairy roots. Int. J. Chem. Eng. Appl. 2013, 157–160.
- Ali, M.; Kiani, B.H.; Mannan, A.; Ismail, T.; Mirza, B. Enhanced production of artemisinin by hairy root cultures of Artemisia dubia. J. Med. Plants Res. 2012, 6.
- Zia, M.; Mannan, A.; Chaudhary, M.F. Effect of growth regulators and amino acids on artemisinin production in the callus of Artemisia absinthium. Pak. J. Bot. 2007, 39, 799–805.
- Yancheva, S.; Georgieva, L.; Badjakov, I.; Dincheva, I.; Georgieva, M.; Georgiev, V.; Kondakova, V. Application of bioreactor technology in plant propagation and secondary metabolite production. J. Cent. Eur. Agric. 2019, 20, 321–340.
- Patra, N.; Srivastava, A.K. Artemisinin production by plant hairy root cultures in gas- and liquid-phase bioreactors. Plant Cell Rep. 2015, 35, 143–153.
- Liu, C.-Z.; Guo, C.; Wang, Y.-C.; Ouyang, F. Comparison of various bioreactors on growth and artemisinin biosynthesis of Artemisia annua L. shoot cultures. Process. Biochem. 2003, 39, 45–49.
- Liu, C.-Z.; Wang, Y.-C.; Guo, C.; Ouyang, F.; Ye, H.-C.; Li, G.-F. Production of artemisinin by shoot cultures of Artemisia annua L. in a modified inner-loop mist bioreactor. Plant Sci. 1998, 135, 211–217.
- Wyslouzil, B.E.; Waterbury, R.G.; Weathers, P.J. The growth of single roots of Artemisia annua in nutrient mist reactors. Biotechnol. Bioeng. 2000, 70, 143–150.
- Patra, N.; Srivastava, A.K. Use of model-based nutrient feeding for improved production of artemisinin by hairy roots of Artemisia annua in a modified stirred tank bioreactor. Appl. Biochem. Biotechnol. 2015, 177, 373–388.
- Patra, N.; Srivastava, A.K. Mass scale artemisinin production in a stirred tank bioreactor using hairy roots of Artemisia annua. Int. J. Biosci. Biochem. Bioinform. 2014, 467–474.
- Liu, C.Z.; Wang, Y.C.; Kang, X.Z.; Fan, O.Y.; Ye, H.C.; Li, G.F. Artemisinin production by adventitious shoots of Artemisia annua in a novel mist bioreactor. Acta Bot. Sin. 1999, 41, 524–527.
- Kim, Y.; Wyslouzil, B.E.; Weathers, P.J. A comparative study of mist and bubble column reactors in the in vitro production of artemisinin. Plant Cell Rep. 2001, 20, 451–455.
- Liu, C.; Wang, Y.; Ouyang, F.; Ye, H.; Li, G. Production of artemisinin by hairy root cultures of Artemisia annua L. Biotechnol. Lett. 1997, 19, 927–929.
- Martin, V.J.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802.
- Tremblay, R.; Wang, D.; Jevnikar, A.M.; Ma, S. Tobacco, a highly efficient green bioreactor for production of therapeutic proteins. Biotechnol. Adv. 2010, 28, 214–221.
- Staub, J.M.; Garcia, B.; Graves, J.; Hajdukiewicz, P.T.J.; Hunter, P.; Nehra, N.; Paradkar, V.; Schlittler, M.; Carroll, J.A.; Spatola, L.; et al. High-yield production of a human therapeutic protein in tobacco chloroplasts. Nat. Biotechnol. 2000, 18, 333–338.
- Wallaart, T.E.; Bouwmeester, H.J.; Hille, J.; Poppinga, L.; Maijers, N.C.A. Amorpha-4,11-diene synthase: Cloning and functional expression of a key enzyme in the biosynthetic pathway of the novel antimalarial drug artemisinin. Planta 2001, 212, 460–465.
- Wu, S.; Schalk, M.; Clark, A.; Miles, R.B.; Coates, R.; Chappell, J. Redirection of cytosolic or plastidic isoprenoid precursors elevates terpene production in plants. Nat. Biotechnol. 2006, 24, 1441–1447.
- Van Herpen, T.W.J.M.; Cankar, K.; Nogueira, M.; Bosch, D.; Bouwmeester, H.J.; Beekwilder, J. Nicotiana benthamiana as a production platform for artemisinin precursors. PLoS ONE 2010, 5, e14222.