ThHere thymus is a primary lymphoid organ derived from the endoderm of the 3rd pharyngeal pouch implicated in the maturationin, we summarize the main characteristics of thymocytes and, therefore, key for establishic epithelial cells emphasizing the immune surveillance. This organ does not contain self-renewing lymphoid precursor cellir origin, principal cell subsets and therefore, is colonized by lymphoid precursors coming either from the fetal liver through mesenchyme or from the adult bone marrow via the blood vessels [1]. Within the thymus, the developingfunctional properties, mainly related with positive and negative thymocytes move throughout a 3D thymic epithelial (TE) network, interacting with the thymic epithelial cells (TECs) of two histologically different compartments: the cortex and the medulla [2] selection, and Treg cell formation.
- Thymus
- Thymic epithelial cell
- T-cell maturation
- central tolerance
1. Introduction
The thymus is a primary lymphoid organ derived from the endoderm of the 3rd pharyngeal pouch implicated in the maturation of thymocytes and, therefore, key for establishing the immune surveillance. This organ does not contain self-renewing lymphoid precursor cells and therefore, is colonized by lymphoid precursors coming either from the fetal liver through mesenchyme or from the adult bone marrow via the blood vessels [1]. Within the thymus, the developing thymocytes move throughout a 3D thymic epithelial (TE) network, interacting with the thymic epithelial cells (TECs) of two histologically different compartments: the cortex and the medulla [2].
2. Thymic Epithelium Is Originated from Epithelial Progenitors (TEPCs) whose Nature Is Controversial
As mentioned above, TE derives from the endoderm of the 3rd pharyngeal pouch [1]. In this early thymic primordium, a common bipotent thymic progenitor cell was identified capable of giving rise to both committed cTEC precursors and mTE progenitor cells [3][4][5][6][7][8]. However, the phenotype of these cells is a matter of discussion and other studies have reported the existence of lineage-specific progenitor cells either for cTECs or mTECs [9][10]. In this respect, it is important to remark that embryonic thymic epithelial progenitor cells (TEPCs) express cell markers (i.e., β5t, CD205, IL7) specific of the cTEC lineage of adult thymus [11][12][13]. These results supported a model of serial progression for explaining the establishment of two major TEC subsets, cortical and medullary. Recently, single cell-RNA sequencing analysis [14] confirmed that most embryonic TECs show a strong cTEC footprint. According to this model, TEPCs would be committed to the cortical epithelial cell lineage by default, whereas the differentiation into mTECs would require the activation of a medullary specific transcriptional program together with the downregulation of cTEC committed genes [15].
On the other hand, embryonic and adult TEPCs seem to be different. Some studies have reported that adult thymi do not contain TEPCs with characteristics of cTECs [9][10] and others remark that bipotent TEPCs do exit in the adult thymus but exhibiting their own molecular characteristics [3][4][5][16]. In any case, adult TEPCs apparently lose their embryonic features, whereas committed progenitor cells either with cTEC or mTEC lineages appear [5][9][10][14][17].
23. Cortical Epithelium and Positive Selection
Reported cTEC specific cell markers are few (i.e., K8, Ly51, CD205) and, consequently, it is difficult to establish conclusively both the phenotype and function of cTEC subsets. The existence of bipotent TEPCs has been reported in a small subset of CD45−EpCAM+Ly51loUEA1−/loMHCIIloIntα6hiSca-1hi TECs [4]. This epithelial cell population is radioresistant and proliferates during thymic regeneration [18]. Previously, Shakib et al. [19] had described a CD205+CD40− cTEC progenitor population enriched in proliferating cells that expressed lower levels of cTEC-specific transcripts (i.e., β5t and cathepsin L), than the more mature cTECs. In addition, a small subset of CD205+ TECs has been reported as immature cTECs [11].
The cortical epithelium is involved in the maturation of DN thymocytes into DP cells through the production of IL7 [20][21] and the surface expression of several Notch ligands, largely Delta-like 4 (Dll4) [22][23], and presents MHC-peptides to DP thymocytes, an essential process for their positive selection. However, it is unclear which cTEC subset is specifically involved in these functions. In adult thymic cortex, cTEC subsets can be immunohistochemically distinguished according to their levels of Ly51 expression. Ly51hi cTECs occupy both the cortico-medullary border and the subcapsule, whereas Ly51lo cells occur throughout the cortex. By flow cytometry, Ly51hi cTECs were MHCIIhi and the Ly51lo cell subset correlated with the MHCIIlo ones [24][25][26]. In addition, adult Ly51hi cTECs express Dll4, whereas most of the Ly51lo cell subset is Dll4− [26][27][28][29]. Accordingly, Ly51hi cTEC might represent a niche for DN cells whereas Ly51lo ones would constitute the niche where DP thymocytes are selected [26].
During their journey throughout thymic cortex, developing thymocytes undergo selection twice. DN3 (CD4−CD8−CD44−CD25+) cells rearrange the TCRαβ alleles [30]; those successfully arranged TCRβ chains are associated with an invariant pre-Tα chain constituting a pre-TCR that apparently does not recognize any ligands [31], but presumably oligomerizes [32], and signals resulting in important proliferation and progression to the DP cell compartment through DN4 (CD4−CD8−CD44−CD25−) cells and an intermediate, immature SP (CD4+CD8− or CD4−CD8+) stage [30][33]. Then, DP thymocytes that have rearranged mature TCRαβ receptors are again selected by MHC-peptide complexes presented by cTECs, which have special mechanisms for processing these peptides. In the case of MHCI-peptide complexes, cTECs have thymic proteasomes that contain the β5t protein encoded by Psmb11 gene [34]. Apparently, the β5t protein of thymoproteasomes has lower chymotrypsin activity than the subunits of the immunoproteasomes consequently producing different peptides inside than outside the thymus. Intimate relationship between thymoproteosome activity and positive selection has been demonstrated in Psmb11−/− mice that exhibit significantly reduced proportions of CD4−CD8+ SP thymocytes and the remaining cells are functionally compromised showing an altered TCRαβ repertoire [35][36]. In addition, a single-nucleotide polymorphism that produces a thymoproteosome variant (G49s) is associated with the appearance of Sjögren’s syndrome and impairs positive selection of CD4−CD8+ thymocytes [37].
Similarly, the generation of a proper repertoire of selecting MHCII-associated self-peptides depends on the presence of both a thymus-specific serine protease, TSSP and the lysosomal protease, cathepsin L in cTECs. Accordingly, alterations in any of these proteases impair the positive selection of CD4+CD8− SP cells [38][39].
34. mTECs Constitute A Heterogeneous Thymic Cell Population Involved in Both Negative Selection and Treg Cell Generation
The medulla displays an important array of mTEC subsets whose functions in the thymocyte education has been extensively studied [40][41]. Currently, it is assumed the existence of immature mTECs, mature mTECs implicated in negative selection and the production of Treg cells, and a post mature stage whose functional significance is more controversial.
In addition, mTEPCs seem to be different in embryonic and adult mice. Whereas the former ones include Claudin (Cld)3/4hi progenitor cells, the expression of this cell marker disappears in the postnatal TEPCs, and Cld3/4hi cells seem to be restricted to mature Aire+ and terminally differentiated mTECs [42]. On the other hand, previous reports classified mTECs into immature MHCIIloCD80lo cells (mTEClo or mTEC-I) and mature MHCIIhiCD80hi cells (mTEChi or mTEC-II), but really mTEClo cells constitute a heterogeneous cell population that could contain both TEPCs and terminal post-Aire cells [17][43][44][45][46][47][48]. Therefore, these mTEClo cells presumably consist, at least, of two cell subsets: putative adult mTEPCs, that highly express podoplanin and ligands of CCR7, such as CCL21a, CCL21b, CCL21c [49][50] and predominantly occupy the cortico-medullary border [17][51], and a more mature CCL21-producing mTEC subpopulation involved in the recruitment of positively selected CCR7+ thymocytes into the medulla [17][50]. Furthermore, Aire+ terminally differentiated mTECs downregulate their specific cell markers (i.e., Aire, MHCII, and CD80) giving rise to Aire−/loMHCIIloCD80lo cells that could be therefore phenotypically considered immature mTECs.
Apart from the upregulation of other cell markers, such as MHCII and CD80 involved in the antigen presentation by mTECs, the most representative marker of the mature mTEC population is the transcription factor Aire. Mature mTEChi acquire Aire expression after RANK-RANKL signaling in immature mTECslo. Thus, mice deficient in RANK or RANKL lack Aire+ mTECshi in the embryonic thymus [52][53]. When the deletion of these molecules was tested in neonatal mice, the reduction of mTEChi cells was not complete [54]. In addition, the CD40-mediated effects on mTEC maturation are weaker that those exerted by RANK signals [53][55].
Firstly, Aire was reported as a defective gene product associated with an autoimmune syndrome called autoimmune polyglandular syndrome type 1 (APS1) [56][57]. The syndrome coursed with autoimmunity affecting diverse organs [58] and hypersensitivity to mucocutaneous candidiasis associated with autoimmune responses against Th17 cytokines, largely IFNα [59][60][61].
Aire promotes the expression of thousands of TSAs that govern the deletion of autoreactive T-cells [62][63][64][65] and the generation of self-reactive Treg cells [66][67][68]. However, Aire is only expressed in 30% of mTECs and controls the expression of approximately 40% of TSAs [69], but the total pool of TSAs expressed in mTECs is highly diverse covering most of the putative self-antigens [70][71]. These results suggest that other cells and/or molecules would be implicated in the central tolerance (see below). In fact, the expression of Aire-dependent TSAs is both stochastic and ordered: only a small fraction of mTECs express a specific TSAs and, at the same time, there is a general pattern of multiple co-expression of scarcely related types of TSAs within the individual mTECs [72][73][74].
Despite the obvious relevance of Aire expression for the establishment of central tolerance, numerous TSAs are expressed in Aire−/− mTECs [72][75]. As mentioned above, another transcription factor, Fezf2, has been recently reported for promoting the expression of a second group of TSAs distinct from the Aire-dependent TSAs [76]. Fezf2-deficient mice do not have defects in positive selection but show altered repertoire of TCR Vβ in both CD4+ and CD8+ T-cells. Additionally, specific elimination of Fezf2 gene in TECs results in autoimmunity with presence of tissue lymphocyte infiltrations and auto-antibodies [69]. Apparently, Aire and Fezf2 genes are controlled independently and do not regulate each other, although recent results indicate that RANK/RANKL signals are key for the appearance of both transcription factors [77]. In any case, only 60% of TSA genes are regulated by Aire and/or Fezf2 suggesting that there are still other unknown mechanisms for TSA production in the thymus. In this respect, the remaining source of TSAs could be peripheral DCs proposed to transport peripheral TSAs into the thymus and to induce T-cell deletion and Treg cell development [78][79][80][81][82][83]. In addition, mTECs transport self-antigens to intrathymic DCs in an Aire-dependent process [84][85].
Increasing evidence supports that mTEC differentiation continues beyond the Aire+ stages of thymic epithelium development, to give rise to the so-called tuft cells as well as to terminally differentiated epithelial cells that form a part of the named Hassall’s bodies (or corpuscles) [50][86][87]. These terminal epithelial cells express numerous markers of differentiated keratinocytes of the outer epidermal layers, such as keratin 1, keratin 10, involucrins, desmogleins, and Serine protease inhibitor Kazal type 5 (SPINK5) [15]. Whether Aire governs or not this final differentiation of mTECs is a matter of discussion [15]. In addition, the functions of these terminal mTECs are controversial. Although they have lost the MHCII expression making efficient antigen presentation to SP thymocytes difficult, they retain a large fraction of both Aire-dependent and Aire-independent TSA that could transfer to DCs. In addition, human Hassall’s bodies, but not murine ones [88], produce Thymus stromal lymphopoietin (TSLP) that induces Foxp3+ Treg cells [89].
Tuft cells are neonatal epithelial cells, firstly identified in mucosae [90][91] and later in the thymus [17][43][92][93]. Studies of lineage tracing demonstrate that more than half of thymic tuft cells derive from mature mTECshi but the rest do not seem to follow the Aire-expressing cell lineage [17][43]. Furthermore, the genetic signature of thymic tuft cells seems to be unique and, in any case, very different from that of Aire+ mTECs.
Mucosal tuft cells produce IL25 and express genes involved in the synthesis of acetylcholine, prostaglandins, or leukotrienes, and in the signaling of taste reception, such as Phospholypase C beta 2 (Plcb2), Transient receptor potential channel subfamily 5 (Trpm5), G protein subunit beta 3 (Gnb3), and G protein subunit gamma 13 (Gng13). Thymic tuft cells also express several members of the taste receptor 2 family as well as L1CAM, DLCK1, MHCII, and CD74, but not other genes characteristic of mucosal tuft cells (i.e., Galectins 2 and 4 (Lgals 2 and Lgals4), Mucin13, Fatty acid binding protein 1 (Fabp1), and Apolipoprotein A1 (Apoa1)) [15].
On the other hand, the functions of thymic tuft cells are largely unknown. Studies on tuft cell-deficient thymuses grafted in mice immunized with IL25 indicate that tuft cells are key to prevent the generation of anti-IL25 antibodies [43]. However, thymic tuft cells do not appear to be good APC because they weakly express MHCII. In addition, they could be related to other thymic cell subsets because tuft cell deficiency results in reduced numbers of both NKT2 cells and Eomes+CD8+ SP thymocytes [43]. Figure 1 summarizes all these described TECs.
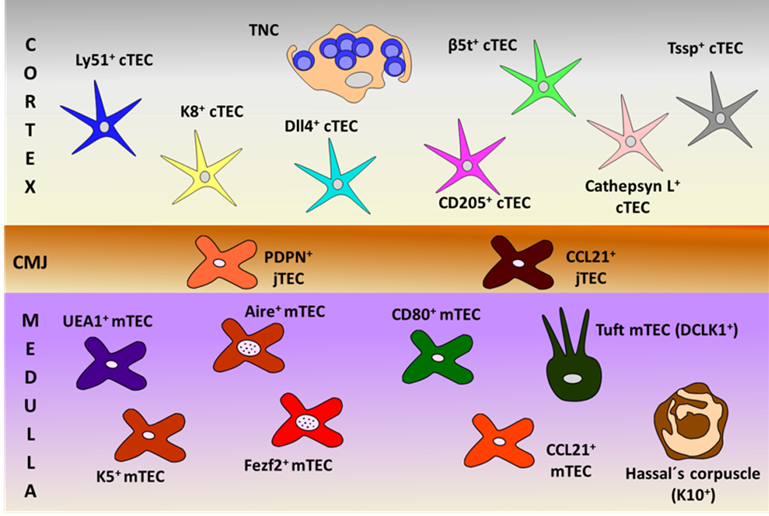
Figure 1. Schematic representation of thymic epithelial cell (TEC) subsets, defined by their specific cell markers in the different thymic compartments (cortex, cortico-medullary junction (CMJ) and medulla). PDPN: podoplanin; TNC: thymic nurse complexes.
References
- Munoz, J.J.; Zapata, A.G. Thymus ontogeny and development. In Thymus Transcriptome and Cell Biology; Passos, G.A., Ed.; Springer: Cham , Switzerland, 2019; pp. 19–34.
- Passos, G.A.; Genari, A.B.; Assis, A.F.; Monteleone-Cassiano, A.C.; Donadi, E.A.; Oliveira, E.H.; Duarte, M.J.; Machado, M.V.; Tanaka, P.P.; Mascarenhas, R. The thymus as a mirror of the body's gene expression. In Thymus Transcriptome and Cell Biology; Passos, G.A., Ed.; Springer: Cham , Switzerland, 2019; pp. 215–234.
- Ulyanchenko, S.; O’Neill, K.E.; Medley, T.; Farley, A.M.; Vaidya, H.J.; Cook, A.M.; Blair, N.F.; Blackburn, C.C. Identification of a Bipotent Epithelial Progenitor Population in the Adult Thymus. Cell Rep. 2016, 14, 2819–2832.
- Wong, K.; Lister, N.L.; Barsanti, M.; Lim, J.M.; Hammett, M.V.; Khong, D.M.; Siatskas, C.; Gray, D.H.; Boyd, R.L.; Chidgey, A.P. Multilineage potential and self-renewal define an epithelial progenitor cell population in the adult thymus. Cell Rep. 2014, 8, 1198–209.
- Lepletier, A.; Hun, M.L.; Hammett, M.V.; Wong, K.; Naeem, H.; Hedger, M.; Loveland, K.; Chidgey, A.P. Interplay between Follistatin, Activin A, and BMP4 Signaling Regulates Postnatal Thymic Epithelial Progenitor Cell Differentiation during Aging. Cell Rep. 2019, 27, 3887–3901.e4.
- Gill, J.; Malin, M.; Hollander, G.A.; Boyd, R. Generation of a complete thymic microenvironment by MTS24(+) thymic epithelial cells. Nat. Immunol. 2002, 3 635–642.
- Rossi, S.W.; Jenkinson, W.E.; Anderson, G.; Jenkinson, E.J. Clonal analysis reveals a common progenitor for thymic cortical and medullary epithelium. Nature 2006, 441, 988–991.
- Bennett, A.R.; Farley, A.; Blair, N.F.; Gordon, J.; Sharp, L.; Blackburn, C.C. Identification and characterization of thymic epithelial progenitor cells. Immunity 2002, 16, 803–814.
- Mayer, C.E.; Zuklys, S.; Zhanybekova, S.; Ohigashi, I.; Teh, H.Y.; Sansom, S.N.; Shikama-Dorn, N.; Hafen, K.; Macaulay, I.C.; Deadman, M.E.; et al. Dynamic spatio-temporal contribution of single beta5t+ cortical epithelial precursors to the thymus medulla. Eur. J. Immunol. 2016, 46, 846–56.
- Ohigashi, I.; Zuklys, S.; Sakata, M.; Mayer, C.E.; Hamazaki, Y.; Minato, N.; Hollander, G.A.; Takahama, Y. Adult Thymic Medullary Epithelium Is Maintained and Regenerated by Lineage-Restricted Cells Rather Than Bipotent Progenitors. Cell Rep. 2015, 13, 1432–1443.
- Baik, S.; Jenkinson, E.J.; Lane, P.J.; Anderson, G.; Jenkinson, W.E. Generation of both cortical and Aire(+) medullary thymic epithelial compartments from CD205(+) progenitors. Eur. J. Immunol. 2013, 43, 589–594.
- Ohigashi, I.; Zuklys, S.; Sakata, M.; Mayer, C.E.; Zhanybekova, S.; Murata, S.; Tanaka, K.; Hollander, G.A.; Takahama, Y. Aire-expressing thymic medullary epithelial cells originate from beta5t-expressing progenitor cells. Proc. Natl. Acad. Sci. USA 2013, 110, 9885–9890.
- Ribeiro, A.R.; Rodrigues, P.M.; Meireles, C.; Di Santo, J.P.; Alves, N.L. Thymocyte selection regulates the homeostasis of IL-7-expressing thymic cortical epithelial cells in vivo. J. Immunol. 2013, 191, 1200–1209.
- Takahama, Y.; Ohigashi, I.; Baik, S.; Anderson, G. Generation of diversity in thymic epithelial cells. Nat. Rev. Immunol. 2017, 17, 295–305.
- Kadouri, N.; Nevo, S.; Goldfarb, Y.; Abramson, J. Thymic epithelial cell heterogeneity: TEC by TEC. Nat. Rev. Immunol. 2020, 20, 239–253.
- Meireles, C.; Ribeiro, A.R.; Pinto, R.D.; Leitao, C.; Rodrigues, P.M.; Alves, N.L. Thymic crosstalk restrains the pool of cortical thymic epithelial cells with progenitor properties. Eur. J. Immunol. 2017, 47, 958–969.
- Bornstein, C.; Nevo, S.; Giladi, A.; Kadouri, N.; Pouzolles, M.; Gerbe, F.; David, E.; Machado, A.; Chuprin, A.; Toth, B.; et al. Single-cell mapping of the thymic stroma identifies IL-25-producing tuft epithelial cells. Nature 2018, 559, 622–626.
- Dumont-Lagace, M.; Gerbe, H.; Daouda, T.; Laverdure, J.P.; Brochu, S.; Lemieux, S.; Gagnon, E.; Perreault, C. Detection of Quiescent Radioresistant Epithelial Progenitors in the Adult Thymus. Front. Immunol. 2017, 8, 1717.
- Shakib, S.; Desanti, G.E.; Jenkinson, W.E.; Parnell, S.M.; Jenkinson, E.J.; Anderson, G. Checkpoints in the development of thymic cortical epithelial cells. J. Immunol. 2009, 182, 130–137.
- Alves, N.L.; Richard-Le Goff, O.; Huntington, N.D.; Sousa, A.P.; Ribeiro, V.S.; Bordack, A.; Vives, F.L.; Peduto, L.; Chidgey, A.; Cumano, A.; et al. Characterization of the thymic IL-7 niche in vivo. Proc. Natl. Acad. Sci. USA 2009, 106, 1512–1517.
- Shitara, S.; Hara, T.; Liang, B.; Wagatsuma, K.; Zuklys, S.; Hollander, G.A.; Nakase, H.; Chiba, T.; Tani-ichi, S.; Ikuta, K. IL-7 produced by thymic epithelial cells plays a major role in the development of thymocytes and TCRgammadelta+ intraepithelial lymphocytes. J. Immunol. 2013, 190, 6173–6179.
- Koch, U.; Fiorini, E.; Benedito, R.; Besseyrias, V.; Schuster-Gossler, K.; Pierres, M.; Manley, N.R.; Duarte, A.; Macdonald, H.R.; Radtke, F. Delta-like 4 is the essential, nonredundant ligand for Notch1 during thymic T cell lineage commitment. J. Exp. Med. 2008, 205, 2515–2523.
- Hozumi, K.; Mailhos, C.; Negishi, N.; Hirano, K.; Yahata, T.; Ando, K.; Zuklys, S.; Hollander, G.A.; Shima, D.T.; Habu, S. Delta-like 4 is indispensable in thymic environment specific for T cell development. J. Exp. Med. 2008, 205 2507-2513.
- Gray, D.H.; Seach, N.; Ueno, T.; Milton, M.K.; Liston, A.; Lew, A.M.; Goodnow, C.C.; Boyd, R.L. Developmental kinetics, turnover, and stimulatory capacity of thymic epithelial cells. Blood 2006, 108, 3777–1785.
- Yang, S.J.; Ahn, S.; Park, C.S.; Holmes, K.L.; Westrup, J.; Chang, C.H.; Kim, M.G. The quantitative assessment of MHC II on thymic epithelium: Implications in cortical thymocyte development. Int. Immunol. 2006, 18, 729–739.
- Cejalvo, T.; Munoz, J.J.; Tobajas, E.; Fanlo, L.; Alfaro, D.; Garcia-Ceca, J.; Zapata, A. Ephrin-B-dependent thymic epithelial cell-thymocyte interactions are necessary for correct T cell differentiation and thymus histology organization: Relevance for thymic cortex development. J. Immunol. 2013, 190, 2670–2681.
- Billiard, F.; Kirshner, J.R.; Tait, M.; Danave, A.; Taheri, S.; Zhang, W.; Waite, J.C.; Olson, K.; Chen, G.; Coetzee, S.; et al. Ongoing Dll4-Notch signaling is required for T-cell homeostasis in the adult thymus. Eur. J. Immunol. 2011, 41, 2207–2216.
- Fiorini, E.; Ferrero, I.; Merck, E.; Favre, S.; Pierres, M.; Luther, S.A.; MacDonald, H.R. Cutting edge: Thymic crosstalk regulates delta-like 4 expression on cortical epithelial cells. J. Immunol. 2008, 181, 8199–8203.
- Tussiwand, R.; Engdahl, C.; Gehre, N.; Bosco, N.; Ceredig, R.; Rolink, A.G. The preTCR-dependent DN3 to DP transition requires Notch signaling, is improved by CXCL12 signaling and is inhibited by IL-7 signaling. Eur. J. Immunol. 2011, 41, 3371–3380.
- Carpenter, A.C.; Bosselut, R. Decision checkpoints in the thymus. Nat. Immunol. 2010, 11, 666–673.
- Kim, S.T.; Takeuchi, K.; Sun, Z.Y.; Touma, M.; Castro, C.E.; Fahmy, A.; Lang, M.J.; Wagner, G.; Reinherz, E.L. The alphabeta T cell receptor is an anisotropic mechanosensor. J. Biol. Chem. 2009, 284, 31028–31037.
- Yamasaki, S.; Ishikawa, E.; Sakuma, M.; Ogata, K.; Sakata-Sogawa, K.; Hiroshima, M.; Wiest, D.L.; Tokunaga, M.; Saito, T. Mechanistic basis of pre-T cell receptor-mediated autonomous signaling critical for thymocyte development. Nat. Immunol. 2006, 7, 67–75.
- Egawa, T.; Tillman, R.E.; Naoe, Y.; Taniuchi, I.; Littman, D.R. The role of the Runx transcription factors in thymocyte differentiation and in homeostasis of naive T cells. J. Exp. Med. 2007, 204, 1945–1957.
- Murata, S.; Sasaki, K.; Kishimoto, T.; Niwa, S.; Hayashi, H.; Takahama, Y.; Tanaka, K. Regulation of CD8+ T cell development by thymus-specific proteasomes. Science 2007, 316, 1349–1353.
- Nitta, T.; Murata, S.; Sasaki, K.; Fujii, H.; Ripen, A.M.; Ishimaru, N.; Koyasu, S.; Tanaka, K.; Takahama, Y. Thymoproteasome shapes immunocompetent repertoire of CD8+ T cells. Immunity 2010, 32, 29–40.
- Takada, K.; Van Laethem, F.; Xing, Y.; Akane, K.; Suzuki, H.; Murata, S.; Tanaka, K.; Jameson, S.C.; Singer, A.; Takahama, Y. TCR affinity for thymoproteasome-dependent positively selecting peptides conditions antigen responsiveness in CD8(+) T cells. Nat. Immunol. 2015, 16, 1069–1076.
- Cheng, M.; Anderson, M.S. Thymic tolerance as a key brake on autoimmunity. Nat. Immunol. 2018, 19, 659–664.
- Nakagawa, T.; Roth, W.; Wong, P.; Nelson, A.; Farr, A.; Deussing, J.; Villadangos, J.A.; Ploegh, H.; Peters, C.; Rudensky, A.Y. Cathepsin L: Critical role in Ii degradation and CD4 T cell selection in the thymus. Science 1998, 280, 450–453.
- Gommeaux, J.; Gregoire, C.; Nguessan, P.; Richelme, M.; Malissen, M.; Guerder, S.; Malissen, B.; Carrier, A. Thymus-specific serine protease regulates positive selection of a subset of CD4+ thymocytes. Eur. J. Immunol. 2009, 39, 956–964.
- Wang, H.X.; Pan, W.; Zheng, L.; Zhong, X.P.; Tan, L.; Liang, Z.; He, J.; Feng, P.; Zhao, Y.; Qiu, Y.R. Thymic epithelial cells contribute to thymopoiesis and T cell development. Front. Immunol. 2019, 10, 3099.
- Dhalla, F.; Baran-Gale, J.; Maio, S.; Chappell, L.; Hollander, G.A.; Ponting, C.P. Biologically indeterminate yet ordered promiscuous gene expression in single medullary thymic epithelial cells. EMBO J. 2020, 39, e101828.
- Hamazaki, Y.; Fujita, H.; Kobayashi, T.; Choi, Y.; Scott, H.S.; Matsumoto, M.; Minato, N. Medullary thymic epithelial cells expressing Aire represent a unique lineage derived from cells expressing claudin. Nat. Immunol. 2007, 8, 304–311.
- Miller, C.N.; Proekt, I.; von Moltke, J.; Wells, K.L.; Rajpurkar, A.R.; Wang, H.; Rattay, K.; Khan, I.S.; Metzger, T.C.; Pollack, J.L.; et al. Thymic tuft cells promote an IL-4-enriched medulla and shape thymocyte development. Nature 2018, 559, 627–631.
- White, A.J.; Withers, D.R.; Parnell, S.M.; Scott, H.S.; Finke, D.; Lane, P.J.; Jenkinson, E.J.; Anderson, G. Sequential phases in the development of Aire-expressing medullary thymic epithelial cells involve distinct cellular input. Eur. J. Immunol. 2008, 38, 942–947.
- Gray, D.; Abramson, J.; Benoist, C.; Mathis, D. Proliferative arrest and rapid turnover of thymic epithelial cells expressing Aire. J. Exp. Med. 2007, 204, 2521–2528.
- Michel, C.; Miller, C.N.; Kuchler, R.; Brors, B.; Anderson, M.S.; Kyewski, B.; Pinto, S. Revisiting the Road Map of Medullary Thymic Epithelial Cell Differentiation. J. Immunol. 2017, 199, 3488–3503.
- Metzger, T.C.; Khan, I.S.; Gardner, J.M.; Mouchess, M.L.; Johannes, K.P.; Krawisz, A.K.; Skrzypczynska, K.M.; Anderson, M.S. Lineage tracing and cell ablation identify a post-Aire-expressing thymic epithelial cell population. Cell Rep. 2013, 5, 166–179.
- Nishikawa, Y.; Nishijima, H.; Matsumoto, M.; Morimoto, J.; Hirota, F.; Takahashi, S.; Luche, H.; Fehling, H.J.; Mouri, Y.; Matsumoto, M. Temporal lineage tracing of Aire-expressing cells reveals a requirement for Aire in their maturation program. J. Immunol. 2014, 192, 2585–2592.
- Onder, L.; Nindl, V.; Scandella, E.; Chai, Q.; Cheng, H.W.; Caviezel-Firner, S.; Novkovic, M.; Bomze, D.; Maier, R.; Mair, F.; et al. Alternative NF-kappaB signaling regulates mTEC differentiation from podoplanin-expressing precursors in the cortico-medullary junction. Eur. J. Immunol. 2015, 45, 2218–2231.
- Wang, X.; Laan, M.; Bichele, R.; Kisand, K.; Scott, H.S.; Peterson, P. Post-Aire maturation of thymic medullary epithelial cells involves selective expression of keratinocyte-specific autoantigens. Front. Immunol. 2012, 3, 19.
- Miragaia, R.J.; Zhang, X.; Gomes, T.; Svensson, V.; Ilicic, T.; Henriksson, J.; Kar, G.; Lonnberg, T. Single-cell RNA-sequencing resolves self-antigen expression during mTEC development. Sci. Rep. 2018, 8, 685.
- Rossi, S.W.; Kim, M.Y.; Leibbrandt, A.; Parnell, S.M.; Jenkinson, W.E.; Glanville, S.H.; McConnell, F.M.; Scott, H.S.; Penninger, J.M.; Jenkinson, E.J.; et al. RANK signals from CD4(+)3(−) inducer cells regulate development of Aire-expressing epithelial cells in the thymic medulla. J. Exp. Med. 2007, 204, 1267–1272.
- Akiyama, T.; Shimo, Y.; Yanai, H.; Qin, J.; Ohshima, D.; Maruyama, Y.; Asaumi, Y.; Kitazawa, J.; Takayanagi, H.; Penninger, J.M.; et al. The tumor necrosis factor family receptors RANK and CD40 cooperatively establish the thymic medullary microenvironment and self-tolerance. Immunity 2008, 29, 423–437.
- Hikosaka, Y.; Nitta, T.; Ohigashi, I.; Yano, K.; Ishimaru, N.; Hayashi, Y.; Matsumoto, M.; Matsuo, K.; Penninger, J.M.; Takayanagi, H.; et al. The cytokine RANKL produced by positively selected thymocytes fosters medullary thymic epithelial cells that express autoimmune regulator. Immunity 2008, 29, 438–450.
- Irla, M.; Hugues, S.; Gill, J.; Nitta, T.; Hikosaka, Y.; Williams, I.R.; Hubert, F.X.; Scott, H.S.; Takahama, Y.; Hollander, G.A.; et al. Autoantigen-specific interactions with CD4+ thymocytes control mature medullary thymic epithelial cell cellularity. Immunity 2008, 29, 451–463.
- Finnish-German, A.C. An autoimmune disease, APECED, caused by mutations in a novel gene featuring two PHD-type zinc-finger domains. Nat. Genet. 1997, 17, 399–403.
- Nagamine, K.; Peterson, P.; Scott, H.S.; Kudoh, J.; Minoshima, S.; Heino, M.; Krohn, K.J.; Lalioti, M.D.; Mullis, P.E.; Antonarakis, S.E.; et al. Positional cloning of the APECED gene. Nat. Genet. 1997, 17, 393–398.
- Perheentupa, J. Autoimmune polyendocrinopathy-candidiasis-ectodermal dystrophy. J. Clin. Endocrinol. Metab. 2006, 91, 2843–2850.
- Kisand, K.; Boe Wolff, A.S.; Podkrajsek, K.T.; Tserel, L.; Link, M.; Kisand, K.V.; Ersvaer, E.; Perheentupa, J.; Erichsen, M.M.; Bratanic, N.; et al. Chronic mucocutaneous candidiasis in APECED or thymoma patients correlates with autoimmunity to Th17-associated cytokines. J. Exp. Med. 2010, 207, 299–308.
- Puel, A.; Doffinger, R.; Natividad, A.; Chrabieh, M.; Barcenas-Morales, G.; Picard, C.; Cobat, A.; Ouachee-Chardin, M.; Toulon, A.; Bustamante, J.; et al. Autoantibodies against IL-17A, IL-17F, and IL-22 in patients with chronic mucocutaneous candidiasis and autoimmune polyendocrine syndrome type I. J. Exp. Med. 2010, 207, 291–297.
- Meloni, A.; Furcas, M.; Cetani, F.; Marcocci, C.; Falorni, A.; Perniola, R.; Pura, M.; Boe Wolff, A.S.; Husebye, E.S.; Lilic, D.; et al. Autoantibodies against type I interferons as an additional diagnostic criterion for autoimmune polyendocrine syndrome type I. J. Clin. Endocrinol. Metab. 2008, 93, 4389–4397.
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039.
- Liston, A.; Lesage, S.; Wilson, J.; Peltonen, L.; Goodnow, C.C. Aire regulates negative selection of organ-specific T cells. Nat. Immunol. 2003, 4, 350–354.
- Anderson, M.S.; Venanzi, E.S.; Chen, Z.; Berzins, S.P.; Benoist, C.; Mathis, D. The cellular mechanism of Aire control of T cell tolerance. Immunity 2005, 23, 227–239.
- Taniguchi, R.T.; DeVoss, J.J.; Moon, J.J.; Sidney, J.; Sette, A.; Jenkins, M.K.; Anderson, M.S. Detection of an autoreactive T-cell population within the polyclonal repertoire that undergoes distinct autoimmune regulator (Aire)-mediated selection. Proc. Natl. Acad. Sci. USA 2012, 109, 7847–7852.
- Malchow, S.; Leventhal, D.S.; Nishi, S.; Fischer, B.I.; Shen, L.; Paner, G.P.; Amit, A.S.; Kang, C.; Geddes, J.E.; Allison, J.P.; et al. Aire-dependent thymic development of tumor-associated regulatory T cells. Science 2013, 339, 1219–1224.
- Perry, J.S.A.; Lio, C.J.; Kau, A.L.; Nutsch, K.; Yang, Z.; Gordon, J.I.; Murphy, K.M.; Hsieh, C.S. Distinct contributions of Aire and antigen-presenting-cell subsets to the generation of self-tolerance in the thymus. Immunity 2014, 41, 414–426.
- Yang, S.; Fujikado, N.; Kolodin, D.; Benoist, C.; Mathis, D. Immune tolerance. Regulatory T cells generated early in life play a distinct role in maintaining self-tolerance. Science 2015, 348, 589–594.
- Takaba, H.; Takayanagi, H. The Mechanisms of T cell selection in the thymus. Trends Immunol. 2017, 38, 805–816.
- Kyewski, B.; Derbinski, J. Self-representation in the thymus: An extended view. Nat. Rev. Immunol. 2004, 4, 688–698.
- Passos, G.A.; Speck-Hernandez, C.A.; Assis, A.F.; Mendes-da-Cruz, D.A. Update on Aire and thymic negative selection. Immunology 2018, 153, 10–20.
- Sansom, S.N.; Shikama-Dorn, N.; Zhanybekova, S.; Nusspaumer, G.; Macaulay, I.C.; Deadman, M.E.; Heger, A.; Ponting, C.P.; Hollander, G.A. Population and single-cell genomics reveal the Aire dependency, relief from Polycomb silencing, and distribution of self-antigen expression in thymic epithelia. Genome Res. 2014, 24, 1918–1931.
- Meredith, M.; Zemmour, D.; Mathis, D.; Benoist, C. Aire controls gene expression in the thymic epithelium with ordered stochasticity. Nat. Immunol. 2015, 16, 942–949.
- Brennecke, P.; Reyes, A.; Pinto, S.; Rattay, K.; Nguyen, M.; Kuchler, R.; Huber, W.; Kyewski, B.; Steinmetz, L.M. Single-cell transcriptome analysis reveals coordinated ectopic gene-expression patterns in medullary thymic epithelial cells. Nat. Immunol 2015, 16, 933–941.
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an immunological self shadow within the thymus by the aire protein. Science 2002, 298, 1395–1401.
- Takaba, H.; Morishita, Y.; Tomofuji, Y.; Danks, L.; Nitta, T.; Komatsu, N.; Kodama, T.; Takayanagi, H. Fezf2 Orchestrates a Thymic Program of Self-Antigen Expression for Immune Tolerance. Cell 2015, 163, 975–987.
- Cosway, E.J.; Lucas, B.; James, K.D.; Parnell, S.M.; Carvalho-Gaspar, M.; White, A.J.; Tumanov, A.V.; Jenkinson, W.E.; Anderson, G. Redefining thymus medulla specialization for central tolerance. J. Exp. Med. 2017, 214, 3183–3195.
- Gallegos, A.M.; Bevan, M.J. Central tolerance to tissue-specific antigens mediated by direct and indirect antigen presentation. J. Exp. Med. 2004, 200, 1039–1049.
- Bonasio, R.; Scimone, M.L.; Schaerli, P.; Grabie, N.; Lichtman, A.H.; von Andrian, U.H. Clonal deletion of thymocytes by circulating dendritic cells homing to the thymus. Nat. Immunol. 2006, 7, 1092–1100.
- Hadeiba, H.; Butcher, E.C. Thymus-homing dendritic cells in central tolerance. Eur. J. Immunol. 2013, 43, 1425–1459.
- Leventhal, D.S.; Gilmore, D.C.; Berger, J.M.; Nishi, S.; Lee, V.; Malchow, S.; Kline, D.E.; Kline, J.; Vander Griend, D.J.; Huang, H.; et al. Dendritic Cells Coordinate the Development and Homeostasis of Organ-Specific Regulatory T Cells. Immunity 2016, 44, 847–859.
- Mbongue, J.; Nicholas, D.; Firek, A.; Langridge, W. The role of dendritic cells in tissue-specific autoimmunity. J. Immunol. Res. 2014, 2014, 857143.
- Proietto, A.I.; van Dommelen, S.; Zhou, P.; Rizzitelli, A.; D'Amico, A.; Steptoe, R.J.; Naik, S.H.; Lahoud, M.H.; Liu, Y.; Zheng, P.; et al. Dendritic cells in the thymus contribute to T-regulatory cell induction. Proc. Natl. Acad. Sci. USA 2008, 105, 19869–19874.
- Hubert, F.X.; Kinkel, S.A.; Davey, G.M.; Phipson, B.; Mueller, S.N.; Liston, A.; Proietto, A.I.; Cannon, P.Z.; Forehan, S.; Smyth, G.K.; et al. Aire regulates the transfer of antigen from mTECs to dendritic cells for induction of thymic tolerance. Blood 2011, 118, 2462–2472.
- Mouri, Y.; Ueda, Y.; Yamano, T.; Matsumoto, M.; Tsuneyama, K.; Kinashi, T.; Matsumoto, M. Mode of Tolerance Induction and Requirement for Aire Are Governed by the Cell Types That Express Self-Antigen and Those That Present Antigen. J. Immunol. 2017, 199, 3959–3971.
- Yano, M.; Kuroda, N.; Han, H.; Meguro-Horike, M.; Nishikawa, Y.; Kiyonari, H.; Maemura, K.; Yanagawa, Y.; Obata, K.; Takahashi, S.; et al. Aire controls the differentiation program of thymic epithelial cells in the medulla for the establishment of self-tolerance. J. Exp. Med. 2008, 205, 2827–2838.
- Nishikawa, Y.; Hirota, F.; Yano, M.; Kitajima, H.; Miyazaki, J.; Kawamoto, H.; Mouri, Y.; Matsumoto, M. Biphasic Aire expression in early embryos and in medullary thymic epithelial cells before end-stage terminal differentiation. J. Exp. Med. 2010, 207, 963–971.
- Odaka, C.; Hauri-Hohl, M.; Takizawa, K.; Nishikawa, Y.; Yano, M.; Matsumoto, M.; Boyd, R.; Hollander, G.A. TGF-beta type II receptor expression in thymic epithelial cells inhibits the development of Hassall's corpuscles in mice. Int. Immunol. 2013, 25, 633–642.
- Watanabe, N.; Wang, Y.H.; Lee, H.K.; Ito, T.; Wang, Y.H.; Cao, W.; Liu, Y.J. Hassall's corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature 2005, 436, 1181–1185.
- Banerjee, A.; McKinley, E.T.; von Moltke, J.; Coffey, R.J.; Lau, K.S. Interpreting heterogeneity in intestinal tuft cell structure and function. J. Clin. Investig. 2018, 128, 1711–1719.
- Gerbe, F.; Legraverend, C.; Jay, P. The intestinal epithelium tuft cells: Specification and function. Cell. Mol. Life Sci. 2012, 69, 2907–2917.
- Panneck, A.R.; Rafiq, A.; Schutz, B.; Soultanova, A.; Deckmann, K.; Chubanov, V.; Gudermann, T.; Weihe, E.; Krasteva-Christ, G.; Grau, V.; et al. Cholinergic epithelial cell with chemosensory traits in murine thymic medulla. Cell Tissue Res. 2014, 358, 737–748.
- Soultanova, A.; Voigt, A.; Chubanov, V.; Gudermann, T.; Meyerhof, W.; Boehm, U.; Kummer, W. Cholinergic chemosensory cells of the thymic medulla express the bitter receptor Tas2r131. Int. Immunopharmacol. 2015, 29, 143–147.