Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Tiziana Maria Sirangelo and Version 2 by Lindsay Dong.
Peach post-harvest ripening is a complex developmental process controlled by a plethora of genetic and epigenetic factors. It leads to protein, lipid and nucleic acid degradation, all resulting in cell death.gene expression; genomics; metabolomics; omics; Prunus persica; ripening; transcriptomicsgene expression; genomics; metabolomics; omics; Prunus persica; ripening; transcriptomics
- gene expression
- genomics
- metabolomics
- omics
- Prunus persica
- ripening
- transcriptomics
1. Introduction
Peach (Prunus persica (L.) Batsch) and its variant nectarine (p. persica (L.) Batsch var. nectarina) are fruits belonging to the Rosaceae family. Peach has become an important model plant due to its small (227.4 Mb) and publicly available doubled haploid genome, whose structure is similar to that of other important fruits such as apricot (p. armeniaca) [1]. Although it has excellent production potential, its consumption is still lower than other popular fresh fruits such as bananas and apples [2]. An effective strategy to promote its consumption could be to enhance its aroma, as well as its quality, which both tend to be affected during extended storage periods, along with its firmness and colour.
Peach ripening is a complex developmental process controlled by a plethora of genetic and epigenetic factors [3]. Its underlying mechanisms lead to structural changes of the fruit that acquires desirable organoleptic qualities and becomes edible. It is a coordinated process requiring a change in the expression levels of hundreds or even thousands of genes to modify a wide range of biochemical and physiological signals such as carbohydrate and organic acid metabolism, chlorophyll breakdown, anthocyanin accumulation, ethylene production, cell wall restructuring, small metabolite and volatile organic compound (VOC) biosynthesis [3].
VOCs are generated in the maturation process through the degradation of compounds such as fatty acids, proteins and carbohydrates and their production varies in different conditions and developmental stages [2]. VOC profiles change during ripening [4][5][4,5], during cold storage (CS) [6][7][6,7], as a result of post-harvest treatments [5], as well as amongst differing germplasm [8] and the compound distribution characterizes the different parts of the fruit [9].
Once fruits are removed from the plant and before they are consumed, a time interval known as post-harvest ripening can occur, whose effects depend on the specific fruit metabolism and maturation state at the time of harvest. Post-harvest ripening leads to protein, lipid and nucleic acid degradation, all ultimately resulting in cell death [10][18]. Furthermore, peach fruits are characterized by a rapid deterioration at room temperature [11][12][19,20]. The post-harvest degradative processes eventually impact fruit quality characteristics, like texture, taste and aroma. For this reason and in order to extend fruit shelf life, different post-harvest strategies, depending on the specific fruit, are applied. Cold storage (CS) treatments are commonly used for enhancing their shelf-life [13][21], even though CS can lead to chilling injury (CI), including changes in texture, and a decline in aroma quality [14][22]. The cold induces changes in gene expression, and an increased expression of the C-repeat Binding Factors (CBFs), related to plant cold acclimation and tolerance, has been reported in many species, including peach [15][16][23,24]. Post-harvest treatments such as controlled atmosphere (CA; a high CO2 concentration and low O2) are used under cold conditions to avoid this CI. A significant reduction of CI was also observed after pre-storage with a high-CO2 treatment [17][25]. In peaches, treatments with 1-methylcyclopropene (1-MCP) can also prevent CI [18][26]. Ultraviolet (UV) light irradiation and specifically UV-B and UV-C treatments, causing a reduction of ethylene levels and therefore a maturation delay, also have become common recently [19][20][27,28]. Many other post-harvest treatments have been applied to peach fruit and it has been found that nectarines have a better resilience to storage conditions than peaches [21][29].
2. The Peach Ripening Process
2.1. Compounds Involved
Although every fruit species and cultivar shows a specific response to hormones, it has long been recognised that in those belonging to the genus Prunus the integrated action of auxins, cytokinins (CKs) and gibberellins (GAs) plays a fundamental role in the regulation of several fruit traits. In addition, abscisic acid (ABA) and ethylene are also important during fruit ripening in all Prunus species [22][23][32,33]. Auxin effects are different in climacteric and non-climacteric fruits: in the former, auxin seems to accelerate the maturation process, while in the latter, auxin may negatively regulate it. A genomic approach has shown that many genes involved in auxin responses including auxin response factors (ARFs) and Aux/IAA increased in expression during peach ripening, highlighting how the hormone plays a key role in this process [24][34]. Other IAA-related genes seem to be up-regulated during the pre-climacteric/climacteric transition: particularly, two putative TIR1 genes, encoding auxin receptors, and one PIN1 gene, encoding a putative auxin efflux facilitator protein. PIN1 expression was found to be mainly induced by exogenous ethylene. This finding shows how in peach a dynamic cross-talk between auxin and ethylene is significant for ripening regulation. This evidence was further supported by changes in the expression of ACS1 and ACO1, two key ethylene biosynthesis genes, during treatment with both of these hormones. In fact, the expression of ACO1 is mostly up-regulated by ethylene, while the highest up-regulating effect on ACS1 was the result of auxin activity [24][25][34,35]. Endogenous levels of CKs play a crucial role during plant growth, through the stimulation of cell division, but they can also partially inhibit auxin responses and be responsible for an increase in fruit size [26][36]. GAs seem to enhance the fruit development process, promoting cell division and enlargement, and delaying peach ripening by decreasing ethylene production [27][37]. Ethylene signalling is associated with a complex transduction pathway involving genes related to traits associated with fruit maturation, such as aroma, colour, firmness and post-harvest shelf-life [23][33]. The levels of this hormone depend on three enzymatic reactions: L-methionine (Met) is converted into S-adenosyl-L-methionine (SAM) by the enzyme S-adenosyl-L-methionine synthetase (MAT), 1-aminocyclopropane-1-carboxylic acid (ACC) synthase (ACS) converts SAM to ACC; and, finally, ACC oxidase (ACO) oxidizes ACC to produce ethylene (Figure 1).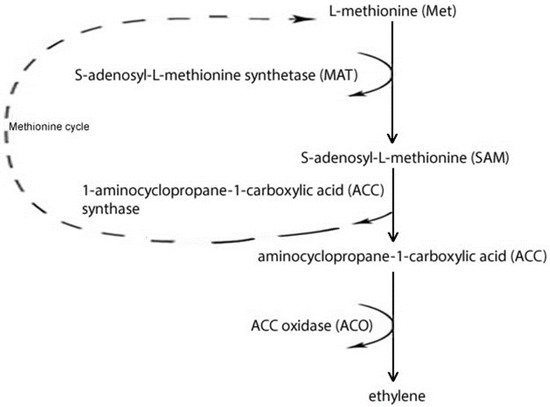
The hormone ABA interacts with ethylene, the cell wall and auxin-related genes in peach [29][41]. In climacteric fruits, it accumulates before ethylene production and reaches maximum levels in fully ripe fruits. Furthermore, ABA enhances anthocyanin and sugar accumulation, promotes softening and is associated with increased ethylene biosynthesis [29][41].
Fatty acids (FAs) are another compound category whose composition changes during peach fruit ripening and has an important influence on aroma production. FA biosynthesis relies on enzymes including lipoxygenases (LOXs), hydroperoxide lyases (HPLs), alcohol dehydrogenases (ADHs), alcohol acyltransferases (AATs) and fatty acid desaturases (FADs) [5]. LOXs catalyse oxidation of polyunsaturated fatty acids, like linoleic and linolenic acids, which accumulate during fruit ripening. Concentrations of linoleic acid were found to increase after peach harvest and then decrease during storage at 20 °C. Linolenic acid increased during the post-harvest ripening phase, especially after about one week. Conversely, palmitic acid showed a two-fold decrease over the same period, while the concentrations of other fatty acids, such as stearic and oleic acid showed no major changes [5].
LOX is responsible for catalysing the hydroperoxidation of polyunsaturated fatty acids to start the synthesis of oxylipins [30][42]. In particular, jasmonates (JAs) are a class of oxylipins that slow down the peach ripening process, often considered as a reference to investigate the effects of this hormone [31][43], and they are also responsible for an interference between ripening and stress defence genes [32][44]. Alterations in the ripening physiology may be related to effects of MJA (Methyl JAsmonate) treatment, often used to delay natural fruit decay [33][45], to prevent CI after harvesting [34][46] and inhibit pathogens [35][47]. Specifically, MJA causes a reduction of the expression levels of the genes responsible for the ripening process and that are found at high levels in this phase, with some also involved in ethylene biosynthesis (e.g., ACO1) [25][35].
Flavonoids such as anthocyanins and flavonols, play a crucial role in fruit colour and health benefits. They vary during peach fruit growth and ripening and R2R3 MYB transcription factors regulate their biosynthesis [36][48].
In peaches, violaxanthin is the predominant xanthophyll both in the skin and in the flesh. It can be found in amounts up to three times higher than other carotenoids [37][51]. Carotenoid levels are highly influenced by post-harvest treatment, including temperature and CA composition [38][52]. In fruits stored at 20 °C, β-criptoxanthin was found to increase, while violaxanthin and β-carotene levels decreased. Refrigeration at lower temperatures resulted in a slowed down carotenoid biosynthesis [39][53].
Overall, the phenolic profile resulting from ripening changes significantly among cultivars and with maturation stage at harvest, even though no general rules can be established [40][41][42][54,56,57]. However, peach cultivars are generally characterized by a higher antioxidant content compared to nectarines, with a similar trend observed for late-harvested versus the early-harvested cultivars [43][58].
2.2. Volatilome and Profiling Techniques
VOCs are a major fraction of the plant metabolome, and being main contributors to fruit aroma and flavour, and thus to fruit quality, have proved to be of particular biological interest. Two lactones, γ- and δ-decalactone are considered to be the most relevant compounds to the final peach aroma [44][45][46][59,60,61]. Particularly, γ-decalactone is the major lactone compound of peach VOCs and it is widely used as a flavour and fragrance agent in the food industry [2][47][48][49][50][51][2,62,63,64,65,66]. Another lactone showing a peach-like odour is γ -jasmolactone, first detected in peach juice [52][67]. A few other VOCs, including δ-octalactone, δ-dodecalactone and 6-pentyl-α-pyrone, are also major contributors of the overall peach aroma [49][64]. C6 aldehydes and alcohols, such as n-hexanal, (E)-2-hexenal and (E)-2-hexenol are also partly responsible for peach aroma, but not to the same extent. Similarly, esters, such as (Z)-3-hexenyl acetate, partially contribute to the flavour [47][62]. Most typical peach VOCs are produced during fatty acid biosynthesis and an involvement of LOX, ADHs and FAs has been proposed [53][68]. All lactones derive from saturated fatty acids, through several steps, including dehydrogenation, epoxidation, hydration, hydroxylation, shortening by β-oxidation and then internal esterification with hydroxyacetyl-coenzyme A [54][55][69,70]. Amongst the VOCs, only a small sub-set impacts the final aroma which increases during fruit ripening. However, VOC production in peach is not a static process, since their level changes dramatically during this stage and flavour compound patterns are different among tissues, species and cultivars, depending on post-harvest conditions [4][5][56][57][4,5,71,72]. In particular, it has been demonstrated that the aromatic profile and organoleptic properties are influenced by both pre-harvest factors, such as growing conditions and maturity stage of the fruit, as well as post-harvest factors, including storage temperature, controlled atmosphere composition, ethylene modulation and the presence of wax coatings [58][59][60][16,73,74]. Overall, some broad trends in VOC changes have been observed. For example, aldehydes tend to decline, and esters to increase in peach fruit over their shelf-life [61][75]. Linalool is a terpene which is reported to be present in peaches at harvest and to rapidly decrease under cold storage [60][74]. CS induces the production of esters and lactones, two compounds typically related with advanced ripening stages [5], that increase during storage at 20 °C after cold treatment [61][62][75,76].3. Peach Gene Expression and Correlation with VOCs
FADs are undoubtedly key enzymes influencing VOC production, but surprisingly their specific role in plant VOC synthesis has not yet been completely elucidated. In peach, ppFAD1 was reported to be involved in the formation of a precursor of lactones/esters [63][127]. Real-time quantitative polymerase chain reaction (qPCR) analysis showed that two ω-3 FAD genes, PpFAD3-1 and PpFAD3-2, may be important in peach VOC biosynthesis since in ripe fruit PpFAD3-1 was high while expression of PpFAD3-2 was low. Instead, high PpFAD3-2 and low PpFAD3-1 transcript levels characterized young fruit [63][127]. The FAD gene family expression during peach ripening and in particular the transcript abundance of PpFAD1 seems to increase in the first days after harvest [5]. In contrast PpFAD2 is found at low levels for one or two days, and then increases together with ethylene and linolenic acid during post-harvest ripening [5]. FADs and their related genes also play a significant role in the changes in lipid membrane fluidity, which is typical of cold-responsive fruits [64][128]. The transcriptional regulation of these genes, in peach, has been associated with metabolic changes occurring during CI [11][19]. Both in several peach cultivars and nectarines, PpFAD4 gene expression was found to decrease under CS, showing, however, very distinctive differences among peach varieties before and after CS [7]. In peach PpLOX1 and PpLOX4, PpLOX2 and PpLOX3, are associated with the synthesis of lactones and of C6 aldehydes [5], and recently differences in expression of these genes amongst cultivars and in response to storage conditions have been shown [7]. Specifically, PpLOX1 expression increased following cold storage in three different nectarine cultivars, whereas in peach cultivars its gene expression fluctuated. Conversely, at the same temperature, PpLOX2, PpLOX3 and PpLOX4 showed a down-regulation with no significant differences among cultivars [7]. The expression of members of the epoxide hydrolase gene family, PpEPH2 and PpEPH3, were found to be involved in the formation of γ-decalactone [51][55][66,70] and to be down regulated during CS in different cultivars [7]. However, the alcohol acyltransferase PpAAT1, which catalyses the biosynthesis of this lactone [65][66][129,130] appeared significantly up regulated after cold exposure in different nectarine cultivars [7]. PpTPS1 terpene synthase, whose expression was found to decline significantly in different peach cultivars after CS [7] is localized in plastids and its expression during cold storage was correlated with the linalool production, while the isoform PpTPS2 was shown to be responsible for (E,E)-α-farnesene (a common biotic-stress-induced plant volatile) biosynthesis in the cytoplasm [67][132]. Under UV-B light treatments, RNA-Seq showed altered transcript levels for these two terpene synthases in peach [67][132], with a decrease of 86% of PpTPS1 and an 80-fold increase of PpTPS2. The reduction of the volatile linalool suggests that the levels of compounds contributing to flavour in peach fruits can be regulated by this ultraviolet treatment. In relation to the production of alcohol VOCs, in CS post-harvest peaches, PpADH2 gene expression seems to depend on the specific cultivar more than on the treatment [7], thus an examination of the other genes related to ester biosynthesis should be evaluated across different cultivars to assess whether they also show cultivar-specific responses. Therefore, VOC profile, gene expression and changes in their response to CS appear to be cultivar specific, and to obtain a complete picture there is the need to test these parameters across a wider range of cultivars.4. Multi-Omics Approaches and the Peach Post-Harvest Ripening Process
NGS technologies have revolutionized plant biology and have been extended widely to non-model systems with very low costs [68][30]. At the same time, combined approaches, based on the relationship amongst genomics, transcriptomics and metabolomics methods, have been developed to exploit these inter-related datasets. Multi-omics applications have been carried out for a wide range of different fruit species and have provided a powerful tool for identifying correlations between different biological components controlling plant functions and metabolic pathways. For instance, a combined analysis of metabolites and transcripts revealed the metabolic shifts underlying tomato fruit development and new associations between specific transcripts and metabolites were identified [69][133]. Similar approaches have been successfully applied to study candidate genes involved in tomato fruit ripening [70][71][134,135] and in other fruit e.g., grape berry development [72][73][74][136,137,138]. In peach, combined omics approaches were applied to identify relationships between fruit VOCs and QTLs for a better understanding of the gene regulation mechanisms behind the biosynthesis of the compounds. For instance, thanks to the availability of an annotated peach genome, QTLs were detected for 23 VOCs and associations between candidate genes and QTLs were established [75][139]. Furthermore, QTLs associated with characters of agronomic interest both to pre-harvest and post-harvest ripening across several Prunus species have been assembled through genotyping with many DNA markers distributed across the entire genome [76][140]. For peach this has been particularly challenging due to the restricted genetic diversity of cultivated peaches, but nevertheless hundreds of QTLs have been identified and related to fruit VOCs. Other combined genomics and metabolomics approaches have investigated pre- and post-harvest ripening processes and confirmed specific loci that control peach aroma [77][141]. By using GC-MS, compounds associated with aroma were also analysed and a correlation-based analysis of these datasets was developed, revealing that the peach volatilome is organized into modules composed of compounds from the same biological pathway or having similar chemical structures. A QTL approach was applied to a very heterogenous peach pedigree, always using the recently available reference peach genome [78][142]. Thanks to use of this genome, the intrachromosomal positions of several QTLs showed differences compared with those previously reported in peach and the mapping quality was generally enhanced. The results of this study provided new insights for a model study for pedigree-based analysis in several peach breeding programs. The validation of genes localized in QTLs through gene expression analysis (RNA-Seq and qPCR) has been recently tested as an approach in peach, in a study aimed at identifying candidate genes involved in fruit softening rate [79][143]. The results, suggesting that auxin may be important in rapid fruit softening, helped to improve our understanding of the genetic mechanisms involved in this process both in peaches and nectarines, and could lead to the identification of molecular markers associated with softening rate. Further multi-omics studies, based on transcriptomic and metabolomic analyses, identified new candidate genes impacting aroma volatiles in pre-harvest and post-harvest conditions in two peach cultivars [55][70]. In this investigation, datasets from microarrays and qRT-PCR analyses were combined with VOCs detected using HS-SPME-GC-MS. The combined dataset was analysed through a correlation-based approach (using CNA) to identify the genes showing a correlation with the major aromatic compounds, including lactones, esters and phenolics. The results showed a core set of genes, including alcohol acyl transferase, fatty acid desaturases and transcription factor genes, that are highly related with peach fruit VOCs and could be useful for future biotechnological activities.Abbreviations
| Abbreviations | Definition |
|---|---|
| 1-MCP | 1-Methylcyclopropene |
| AATs | Alcohol acyltransferases |
| ABA | Abscisic Acid |
| ACC | Aminocyclopropane-1-carboxylic acid |
| ADHs | Alcohol Dehydrogenases |
| AVG | Aminoethoxyvinylglycine |
| BAC | Bacterial Artificial Chromosome |
| CA | Controlled Atmosphere |
| CBF | C-Repeat Binding Factor |
| cDNA | complementary DNA |
| CI | Chilling Injury |
| CKs | Cytokinins |
| CNA | Correlation Networks Analysis |
| CRISP/Cas9 | Clustered Regularly Interspaced Short Palindromic Repeats/CRISP associated protein 9 |
| CS | Cold Storage |
| DEG | Differentially Expressed Gene |
| EST | Expressed Sequence Tag |
| GAs | Gibberellins |
| GC-FPD | Gas Chromatography-Flame Photometric Detection |
| GC-MS | Gas Chromatography–Mass Spectrometry Techniques |
| GC-O | Gas Chromatography-Olfactometry |
| GWAS | Genome-Wide Association Study |
| HPLs | Hydroperoxide Lyases |
| HS-SPME | Headspace Solid-Phase Microextraction |
| Jas | Jasmonates |
| LC-MS | Liquid Chromatography–Mass Spectrometry Techniques |
| LOXs | Lipoxygenases |
| MET | L-methionine |
| MJA | Methyl Jasmonate |
| NGS | Next Genetation Sequencing |
| NMR | Nuclear Magnetic Resonance |
| QTL | Quantitative Trait Loci |
| qRT-PCR | Real Time or Quantitative PCR–Polymerase Chain Reaction |
| SAM | S-Adenosyl Methionine |
| SMRT | Single Molecule Real-Time |
| SNPs | Single Nucleotide Polymorphisms |
| SVs | Structural Variants |
| TD | Thermal Desorption |
| UV | Ultraviolet |
| VOC | Volatile Organic Compounds |