1. The Circadian Clock Circuitry
In mammals, the system driving the generation of internal timing and ultimately the regulation of physiology and behaviour is formed by a hierarchical network of biological oscillators with two hubs in the hypothalamic suprachiasmatic nuclei (SCN)
[1][12]. The SCN clock (main pacemaker) is necessary for the maintenance of 24-h body rhythms
[2][3][13,14] and can restore
[4][15] or de novo establish circadian rhythms in gene expression when transplanted in genetically arrhythmic mice
[5][6][16,17]. The circadian circuitry encompasses molecular clocks of the main pacemaker and additional cellular clocks in peripheral tissues
[7][8][9][18,19,20]. These are driven by the SCN through neural and humoral outputs
[10][21] and, together with the main pacemaker, manage circadian timing of physiological processes and behaviour
[11][12][2,22]. One of the main external synchronizers (or zeitgeber) is light, a strong central pacemaker-tuning cue
[13][23]. Light information is perceived by retinal ganglion cells expressing the photopigment melanopsin and passed on to the SCN through retino-hypothalamic tract fibres
[14][15][16][17][24,25,26,27] and to neighbouring and downstream oscillators via neurotransmitters such as glutamate, PACAP, VIP, GRP
[15][18][19][25,28,29], VP
[20][21][30,31], and GABA
[22][32]. Even though the oscillators in the SCN are capable of autonomous ticking, the oscillations become progressively dampened in peripheral oscillators in the absence of SCN-synchronizing signals
[23][33].
2. The Molecular Mechanisms of Biological Ticking
The molecular clockworks generate rhythmic oscillations through “transcription-translation feedback loops” (TTFL), i.e., core-clock transcription factors positively turn on the expression of genes encoding circadian proteins that subsequently inhibit transcription via negative-feedback loops, with a time delay indispensable for the proper functioning of the biological clock
[24][34]. The TTFL is manoeuvred by several core-clock mRNAs and proteins, including activators (BMAL1, CLOCK, RORα, RORβ, and RORγ) and repressors (PER1, PER2, PER3, CRY1, CRY2, REV-ERBα, and REV-ERBβ), kinases (CKIα, CKIδ, and CKIε), and phosphatases (PP1 and PP5) that control protein stability and subcellular localization. CLOCK (or its paralogue NPAS2) and BMAL1 (alias ARNTL), as basic helix-loop-helix-PAS (bHLH–PAS) transcription factors
[24][34] heterodimerize and bind to E-boxes in the promoter regions of the genes encoding the period (PER1, PER2, and PER3) and cryptochrome (CRY1 and CRY2) proteins, which upon heterodimerization in the cytoplasm translocate back into the nucleus and repress CLOCK:BMAL1-mediated transcription. Then, ubiquitination-dependent degradation of PER/CRY complexes takes place and a new round of CLOCK:BMAL1-mediated transcription restarts
[24][34].
3. Viruses and Circadian Clock Circuits
Synchronization with the external environment and anticipation of predictable environmental changes confer competitive advantages to living beings, especially in the wild. Accordingly, in humans experiencing dysregulation of circadian timing, for example, due to shift work or social jet lag, there is increased incidence of various degenerative, inflammatory, metabolic, and neoplastic diseases
[25][26][35,36]. Besides human health and wellbeing maintenance, a detailed knowledge of the molecular mechanisms operating biological clocks has implications also for the efficacy of treatment strategies for pathological states.
Mounting evidence shows that time-of-day-related treatment schedules and chrono-modulated drug delivery significantly affect both effectiveness and side effects of pharmacological therapy
[27][28][29][37,38,39]. Diseases caused by viruses, rickettsiae, bacteria, fungi, or parasites continue to represent a primary cause of morbidity. Infections by viruses rework the biological processes of infected cells to facilitate replication and spread, and thus the molecular interplay between the mechanisms of the biological clock, immune system, and virus infection can influence disease outcomes (
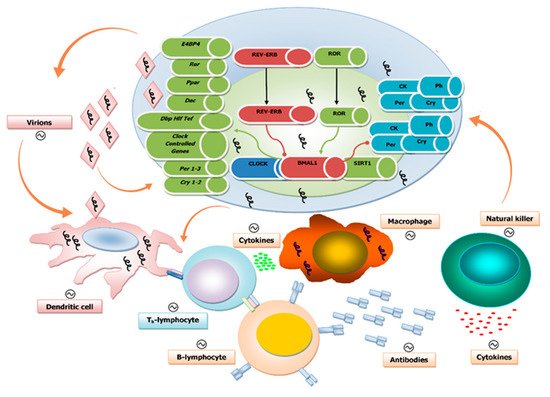
Figure 1).
Figure 1. Schematic illustration of the interplay between the biological clock, virus replication, and the immune system at the cellular level: The components of the molecular clockwork are depicted within the cell, with green arrows indicating activation and red arrows indicating inhibition. The presence of viral genomes is indicated by squiggles. Below the cell are elements of the immune response. The orange arrows indicate interactions among virions, biological clocks, and immune competent cells. The interaction of virions and their nucleic acid core with these two players impacts viral replication and rhythmic patterns of host–hosted molecular trade off. The immune system with its innate and adaptive arms provides shielding against viral infections with a number of molecular factors and effectors, such as dendritic cells, T and B cells, macrophages, and natural killer cells. These are engaged to hold up and hinder virus replication and dissemination through the secretion of cytokines and the production of specific antibodies. The components and, ultimately, the complex function of the immune system are rhythmically driven by the biological clock and, in turn, influence the function of the molecular clockwork. Viral particles impact the interplay between immune and circadian systems.
4. Circadian Regulation of Both Innate and Adaptive Immune Systems
The circadian regulation of the immune system response allows for its time-dependent success against different pathogens in a rhythmic fashion
[30][31][32][33][34][40,41,42,43,44]. Immune cells of the innate and adaptive components of the immune system have molecular clockworks managing their rhythmic processes on a 24-h timescale
[35][36][37][38][39][45,46,47,48,49], such as lymphocyte migration through lymph nodes and lymph in mice
[40][50].
The rhythmic function of the immune system influences the appearance of pathological conditions, for instance, inflammatory lung disease and asthma, which show circadian fluctuations in symptoms and occurrence. In a recent study using both acute and chronic models of virus-induced airway disease (based on the Sendai virus (SeV)), disturbance of the 24-h periodicity negatively affected development, progression, and exacerbation of asthma
[41][51]. In this study,
BMAL1 silencing lead to more pronounced asthma-typical airway changes, which suggests a role for
BMAL1 in the regulation of lung-specific antiviral responses and the subsequent development of asthmatic symptoms. This study reinforces the previous findings from Majumdar et al., who reported a role for BMAL1 as a regulator of innate immunity and showed that BMAL1-deficent cells are more susceptible to infection of the RNA viruses RSV (respiratory syncytial virus) and PIV3 (parainfluenza virus type 3)
[42][52].
The efficacy of vaccination is also thought to be under circadian control as suggested by results obtained in mice and human studies. Silver et al. have shown in immunized mice that vaccination at the time of higher TLR9 (Toll-like receptor 9) expression led to enhanced immune response, likely due to the circadian expression pattern of TLR9
[34][44]. Interestingly, in the same study,
PER2-deficient mice did not show the observed time variation of immune response. Likewise, the antibody response to viral challenge in humans was also reported to be time dependent: immunization with hepatitis A and influenza virus vaccines in the morning led to higher antibody response as compared to afternoon vaccination
[43][44][45][53,54,55].
5. Influence of the Biological Clock on Virus Replication Cycle and Disease
Cellular and tissue functions exhibit rhythmic fluctuations over the course of 24 h that impact the susceptibility to or progression of the viral infection. Several core-clock elements have been reported to modulate viral infection. BMAL1 and PER2 are regulators of antiviral immunity, as described in the previous section, and additional roles for PER1 and REV-ERBα, as well as melatonin have been highlighted, as described in the following. The seasonal variation of BMAL1 expression, reaching its lowest values in winter, seems to contribute to the high occurrence of respiratory viral diseases in this season since low
BMAL1 expression enhances viral disease
[46][47][56,57]. An effect of the circadian clock in viral infection was also reported for wild-type mice infected with murine herpesvirus 4 at different times of the day. The mice were kept under controlled temperature and lighting environment (12-h light/12-h darkness), and the levels of infection and spread of the virus were evaluated
[48][58]. In mice infected at the beginning of the day, corresponding to the beginning of the resting phase of these nocturnal animals, the viral replication was increased tenfold compared to the values measured in mice infected during the night period, corresponding to the activity phase. In mice lacking
BMAL1, the levels of virus replication were independent of the time of day at which the viral infection was performed
[48][58]. The effect of time of infection on virus replication was independent of the immune system, as also confirmed on single cell cultures (embryonic fibroblasts), in which several time-dependent processes (intracellular trafficking, biosynthetic processes, protein synthesis, and chromatin assembly) were involved. The obliteration of cellular circadian rhythms increased the replication of herpes and influenza A viruses, which lost rhythmic fluctuation and remained persistently high
[48][58]. The time of day of herpes simplex virus type 2 (HSV-2) infection impacted the outcome of the pathology in wild-type mice kept under 12-h light and 12-h darkness (LD 12:12), probably due to the circadian rhythmic expression of the HSV-2 receptor in the skin Nectin1. This effect was confirmed in immunodeficient RAG2-knockout mice
[49][59].
6. Disruption of the Circadian Clock by Viruses
Viral infection may also alter circadian gene expression as reported in different studies. Simian immunodeficiency virus (SIV) infection in monkeys caused changes in amplitude and values of body temperature and locomotor activity circadian rhythmicity after the acute retroviral syndrome stage. These changes were not linked to modifications detected in the acute febrile response triggered by virus inoculation. On the other hand, hypothalamic microglia infiltration and macrophage accumulation were shown to occur in animals sacrificed upon appearance of strong circadian anomalies, hinting at analogously significant physiological and psycho-cognitive consequences in human subjects with HIV infection
[50][68]. Moreover, in vitro experiments performed in a cell line (Bel-7404-HBx) stably transfected with the hepatitis B virus X protein (HBx, involved in hepatocellular carcinogenesis) showed alterations in the mRNA expression levels for several core-clock genes, with upregulation of
CLOCK,
PER1, and
PER2 and downregulation of
BMAL1,
PER3,
CRY1,
CRY2, and
CKIɛ [51][69].
7. The Biological Clock and Influenza Virus Infection
A worldwide problem is represented by seasonal epidemics of influenza. Influenza is a disease with global impact that causes enormous morbidity and mortality on an annual basis. The severity of the infection depends on both the virus strain and a number of host factors, primarily age and the presence of comorbid conditions. The mortality and usage of healthcare resources associated with influenza is focused in the elderly and in those with a coexisting disease such as chronic obstructive pulmonary disease (COPD). In patients with COPD, the morbidity and mortality caused by an influenza virus infection are considerably greater.
In a COPD/emphysema mouse model, influenza A virus (IAV) infection reworked lung clock gene expression and decreased the amplitude in the rhythms of locomotor activity. This effect was more prominent in C57BL/6J mice that were chronically exposed to cigarette smoke and was accompanied by decreased body weight and augmented mortality; likewise, Bmal1 knockout mice infected with IAV showed amplified lung inflammatory and pro-fibrotic responses.
8. Epigenetic Mechanisms Underlying the Interaction Between Viral Infection and the Circadian Clock Machinery
Epigenetic regulatory mechanism, which includes DNA methylation and histone posttranslational modifications (PTM), leads to chromatin remodelling and regulated gene expression. Chromatin remodelling represents a critical process through which inputs, such as light or food, are transduced by the cell to generate permissive and silencing histone modifications to influence gene transcription and ultimately signalling pathway activity. CLOCK-BMAL1-dependent activation of clock-controlled genes is connected to circadian changes in histone PTM at their own promoters. Numerous chromatin remodelers, such as the deacetylases Sirtuin 1 (SIRT1, a NAD+-dependent histone deacetylase) and histone deacetylase 3 (HDAC3), are recruited to the promoter region of the clock-controlled genes in a circadian manner
[24][52][34,73]. Importantly, the core element of the clockwork machinery, the transcription factor CLOCK, displays histone acetyltransferase activity as well. In the absence of these chromatin modifiers, the rhythmic expression of the clock-controlled genes is abrogated
[52][73]. One of the most common epigenetic modifications is the methylation of DNA (addition of a methyl group to the cytosine residue in CpG dinucleotides mediated by the family of DNA methyltransferase enzymes DNMT1, DNMT3a, and DNMT3b). Overall DNA methylation with focal hypermethylation on the promoter of tumour suppressor genes, causing transcriptional silencing, is a mechanism of carcinogenesis
[53][54][55][74,75,76]. These epigenetic mechanisms are critical for the functioning of the biological clock
[52][73] and are disrupted by viral infection
[53][54][55][74,75,76].