Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Bettina Toth and Version 2 by Conner Chen.
As chemotherapy—in contrast to radiation—is mainly applied to all cancer patients, a myriad of studies already exists, which indicate that commonly used agents cause premature ovarian insufficiency (POI) by inducing the death or accelerated loss of primordial follicles as well as damage to blood vessels, stromal cells, and immune components.
- low-dose radiation
- ovarian damage
1. Introduction
The overall five-year relative survival rate of all childhood cancers has significantly improved over the last thirty years from 58% to 83%, giving rise to a special population: adult survivors of childhood cancer after cytotoxic chemo- and radiotherapy (RT) with the desire to have children regardless of their medical history [1][2][1,2]. Fertile women have lower chances to become a mother after surviving cancer treatments [3].
As chemotherapy—in contrast to radiation—is mainly applied to all cancer patients, a myriad of studies already exists, which indicate that commonly used agents cause premature ovarian insufficiency (POI) by inducing the death or accelerated loss of primordial follicles as well as damage to blood vessels, stromal cells, and immune components [4][5][6][7][8][9][10][11][12][4,5,6,7,8,9,10,11,12]. However, data on (low-dose) radiation in fertile women are urgently needed.
Radiotherapy accelerates oocyte depletion by different mechanisms such as apoptosis or oxidative stress, leading to acute ovarian failure (AOF), POI, and menopause. Radiotherapy is a cornerstone of state-of-the-art cancer therapy in children, women, and men. At present, around 50–60% of all patients with long-term cancer (45–50% curative success for all cancer types) receive radiation therapy alone or in combined treatment schedules [13]. The direct and undisputed beneficial effects of targeted radiation on tumor cells result from preventing further cell proliferation or inducing cell death. Irrespective of this, the surrounding organs can be directly damaged by scattered radiation [13][14][13,14].
The human ovary contains a finite pool of primordial follicles (PMF), which comprise the ovarian reserve. Its maximum is already reached as a fetus at five months of gestation, which then declines with increasing age and culminates in the menopause at an average age of 50–52 years [14][15][14,15]. This fixed number of oocytes is non-renewable and must provide for the entire reproduction cycle throughout adult life. The maximum reserve during fetal life is followed by atresia and the loss of more than half of the originally developed germ cell pool at the time of birth. Only oocytes that are enclosed by a sufficient number of epithelial and stromal cells are able to survive. Therefore, the interaction between oocytes and stromal cells is of utmost importance even during prenatal development [16][17][18][16,17,18]. Different cell populations of the ovarian cortex can be defined by gene expression analyses: stroma (83%); oocytes (0.2%); perivascular (10%); endothelial (5%); granulosa (1.2%); and theca and immune cells (0.4%) [19][20][19,20].
In the last decade, maintaining the gonadal function and preserving fertility after successful cancer treatment have become critical and have increased the concerns of young fertile patients. The options depend upon the age of the patient, their physical state, the administered agent, and the start of the cancer treatment. Regarding radiation of the pelvis, ovaries can be transposed outside the radiation field (ovarian transposition). This surgical technique has been performed since 1952 with low success rates and a high risk of POI (33–100%) also due to scattered radiation [21][22][23][21,22,23]. Patients undergoing brachytherapy seem to benefit most with ovarian survival rates from 77.8–100% [24]. However, an increased risk of ovarian cysts after ovarian transposition has been reported, ranging between 0 and 34% [25][26][25,26]. Together with other complications such as abdominal pain, hematomas, tubal ligation, ischemia, or small bowel obstructions, reoperation was necessary in 34.7% of patients with complications [24]. Therefore, ovarian transposition may be offered to women with planned pelvic radiation without chemotherapy as being recommended by international guidelines [27][28][29][30][27,28,29,30]. Of note, supporting evidence is weak; therefore, ovarian transposition may not act as a safe procedure for fertility preservation with special regard to the above-mentioned complications. Another technique—which can only be offered after puberty—is the cryopreservation of oocytes before the legal age and the cryopreservation of (non) fertilized oocytes thereafter. In order to cryopreserve, women need to undergo controlled hormonal stimulation before the oocytes are transvaginally retrieved [31]. Therefore, this technique can only be offered if the cancer treatment can be postponed for at least 2–3 weeks.
The cryopreservation of ovarian tissue was first performed by Prof. Dr. Donnez in 1997. After successful cancer treatments, the tissue can be thawed and transplanted onto the remaining ovary or into a peritoneal tissue pouch near the ovaries. As the first successful ovarian tissue transplantation was performed in 2004, this technique is now routinely performed. The resumption of cyclic hormone production can be achieved in up to 63% of cases, and live birth rates (LBR) are described as 23% per transplantation [32][33][32,33]. To date, more than 170 live births after the transplantation of frozen–thawed ovarian cortical pieces have been reported, and the transplanted ovarian tissue remains active from six months to three years [32]. However, the cryopreservation of ovarian tissue is not established in children and only parts of the ovary (40–50%) can be cryopreserved whereas the major part stays within the pelvis and is exposed to radio-chemotherapy.
2. The Human Ovary and Folliculogenesis
Folliculogenesis
Human folliculogenesis has been described in detail in the literature [34][35][45,46]. Meiosis of the oogonia starts in early fetal life and the final PMF pool is formed around 24 gestational weeks in humans and around birth in rodents (Figure 12). Once meiosis is initiated, mitosis ends; therefore, the individual PMF pool is already fixed before birth. Of note, POI occurs in 1% of women under the age of 45 years without the influence of gonadotoxic treatment due to a low PMF pool [36][47]. After menarche, the follicles either start to grow or become atretic. The activation of PMF includes different pathways such as PI3K/PTEN7Akt and Hippo [37][38][39][40][48,49,50,51]. However, the detailed mechanisms of the regulation of the PMF pool over the reproductive female lifespan remain unknown. PMFs are mainly located in the ovarian cortex, which represents the poorest vascularized zone in the human ovary. Both primordial and early growing follicles are dependent on stromal vessels as they do not rely on an independent vascular network [41][52]. PMFs consist of an oocyte surrounded by granulosa cells [42][53]. The high proliferation rate of granulosa cells explains their sensitivity to chemotherapy and radiation. The secondary follicle is characterized by the formation of a zona pellucida, stromal cells, and an undifferentiated theca layer. After differentiation into a theca interna and a theca externa layer, the stage of a pre-antral follicle is reached followed by an early antral follicle with fluid-filled cavities and a large antral follicle with a visible large antrum and a marginal oocyte [35][42][46,53].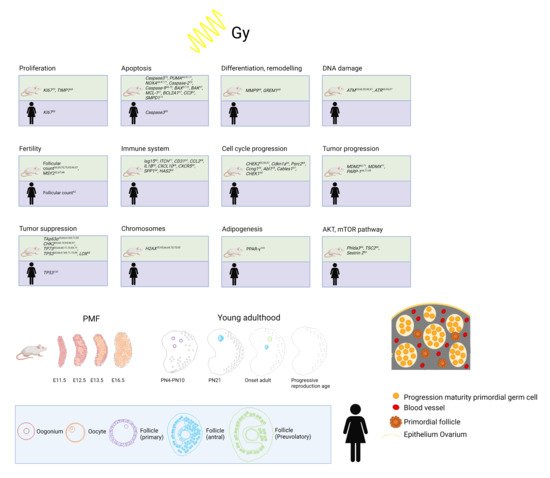
Figure 12. Top: Overview of already established markers concerning ovarian damage including proliferation, apoptosis, differentiation and remodeling, DNA damage, fertility, immune system, cell cycle progression, tumor progression, tumor suppression, chromosomes, adipogenesis, and AKT/mTOR pathway after radiation in human and mouse models. Bottom: Development from PMF pool to young adulthood in mice and humans and human folliculogenesis and detail in PMF pool in humans at birth. Adapted from [54,55,56,57,58,59,60,61,62].