Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Rita Xu and Version 1 by Hui-Jun Liao.
Dipeptidyl peptidase 4 (DPP4) inhibitors can treat type 2 diabetes by slowing GLP-1 degradation to increase insulin secretion. Studies have reported that
Pseudostellaria heterophylla
,
Linum usita-tissimum
(flaxseed), and
Drymaria diandra
, plants rich in Caryophyllaceae-type cyclopeptides and commonly used as herbal or dietary supplements, are effective in controlling blood sugar.
- Pseudostellaria heterophylla
- Linum usitatissimum
- Drymaria diandra
- cyclic peptides
- cyclopeptides
- DPP4
- diabetes
1. About Diabetes
Diabetes is a metabolic disorder characterized by increased blood glucose levels. With urbanization and social and cultural change in food behavior, the prevalence of diabetes is rapidly increasing. Diabetes and its extended vascular complications, diabetic nephropathy, retinopathy, etc., cause heavy physical and mental pressure to patients [1]. Diabetes is classically divided into three types: type 1, type 2, and gestational diabetes, with type 2 diabetes (T2D) accounting for the majority. Type 2 diabetes is caused by insulin resistance, which refers to impaired sensitivity to insulin-mediated glucose disposal or insufficient insulin secretion [2]. Type 2 diabetes can be prevented by maintaining a normal weight, regular exercise, and eating a healthy diet [3]. However, some will still face the need for medication. Metformin is the first-line hypoglycemic agent recommended when most patients are initially on drug control. In addition to metformin, new hypoglycemia treatments are constantly being proposed due to the high demand for T2D drugs and the emergence of resistance.
2. Mechanisms of Incretins (GLP-1 and GIP) in Glucose Homeostasis and Diabetes Treatment
Incretins are hormones secreted by endocrine cells of the intestinal epithelium with food to maintain blood sugar balance. The two most studied incretins are glucagon-like peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) [4]. Studies have found that GLP-1 and GIP can stimulate pancreatic β-cells to increase insulin synthesis and secretion to help stabilize blood sugar after meals [5]. More follow-up studies have indicated that GLP-1 has a greater impact on blood sugar than GIP. In addition to stimulating insulin secretion from β cells, GLP-1 can also inhibit the secretion of glucagon from α cells and increase the secretion of somatostatin by δ cells. Therefore, GLP-1 accounts for more than GIP in studies of diabetes based on the incretin system [6,7][6][7]. The bioactive forms of GLP-1 include GLP-1 (7−37) and GLP-1 (7−36) NH2. These active peptides maintain blood glucose homeostasis by activating the GLP-1 receptor (GLP-1R) on β cells, triggering a series of downstream reactions. The activation of GLP-1R by GLP-1 or GLP-1 analogs directly results in cAMP accumulation, followed by increased insulin secretion. Longer-term effects also include promoting β-cell proliferation and reducing β-cell apoptosis. This is of great significance in delaying the depletion of pancreatic islet cells in diabetic patients [6,7][6][7]. The effects of GLP-1 are also involved in extra-pancreatic events. The topic of greatest interest was the effect of GLP-1 on appetite and weight loss. The release of GLP-1 and related effects delay gastric emptying and bowel motility, and in addition, exert pressure on the hypothalamus to alter satiety; thereby, suppressing appetite and assisting in weight control [7].
Endogenous GLP-1 in the circulation will be immediately degraded by dipeptidyl peptidase 4 (DPP4) into inactive metabolites GLP-1 (9−37) and GLP-1 (9−36) NH2 (t 1/2 ~1–2 min). In T2D patients, reduced GLP-1 secretion or decreased GLP-1 response, also known as incretin deficiency, has been observed, resulting in poor postprandial blood glucose regulation [6]. DPP4 inhibitors are designed to interfere with the enzymatic activity of DPP4, reducing its rate of cleavage of GLP-1 to increase the concentration of active GLP-1 in plasma. Another incretin-based therapy for T2D is to apply GLP-1 analogs to mimic the effects of endogenous GLP-1. The hypoglycemic effects of incretin-based therapies for T2D have been proven [7]. Clinically available oral DPP4 inhibitors include Sitagliptin, Vildagliptin, Saxagliptin, Alogliptin, Linagliptin, etc. GLP-1 analogs such as Exenatide, Liraglutide, and Semaglutide (now also available in oral form) typically require administration by injection and are therefore less convenient than the orally available DPP4 inhibitors [7,8,9][7][8][9]. One of the advantages of incretin-based therapies is that since the effect of GLP-1 depends on blood glucose concentration, it rarely causes hypoglycemia. After the treatment reaches the tolerated dose of metformin and the patient’s blood sugar control is still unsatisfactory, DPP4 inhibitors can be used alone or in combination with metformin as second-line therapy [10]. According to clinical observations, DPP4 inhibitors have an apparent influence on the activity of DPP4, reducing the baseline by more than 50% [11]. When Sitagliptin (the first listed DPP4 inhibitor) monotherapy is used to treat adult T2D patients, the observed efficacy includes improvement in HbA1c, fasting plasma glucose (FPG), and 2-h postprandial glucose (PPG). In addition, most studies have reported its beneficial effects on the regulation of triglyceride (TG), HDL-c, and LDL-c [6,12][6][12].
3. The Structure of DPP4 and the Interaction of DPP4 Inhibitors with DPP4
Dipeptidyl peptidase 4 (DPP4), also known as CD26 (cluster of differentiation 26), is a type II transmembrane serine protease with 766 amino acids (110 kDa) anchored to the membrane, that selectively cleaves the Xaa-proline or Xaa-alanine dipeptides from the N-terminus of GLP-1 [11]. Transmembrane DPP4 enhances its enzymatic activity through dimerization. Matrix metalloproteinases (MMPs) cleave DPP4 on the membrane. Cleaved DPP4 lacks the cytoplasmic domain (aa 1-6), transmembrane domain (aa 7-28), and flexible stalk (aa 29-39) and becomes a circulating or soluble form (sDPP4, aa 39-766) [11,13,14,15][11][13][14][15]. sDPP4 is less studied relative to DPP4 on the membranes. The extracellular part of the monomer in dimeric DPP4, in addition to the flexible stalk, mainly includes a large cavity constructed between the eight-bladed β-propeller domain (aa 54-497) and α/β hydrolase domain (aa 39-51 and 506-766) with a set of entrances for GPL-1 and GIP, etc. to in and out [11,13,14,15][11][13][14][15]. The catalytic triad (composed of Ser630, Asp708, and His740) and its adjacent amino acids Glu205 and Glu206 (to ensure the anchoring of the N-terminus of the substrate) have a great influence on the enzymatic activity of DPP4. Moreover, Arg125 and Asn710 contribute to electrostatic adsorption; Tyr662 and Tyr666 form hydrophobic pockets; and Tyr547 is responsible for oxygen anion holes, constituting a series of important amino acids in the active site of DPP4 [11]. The area enclosed by the amino acid residues Glu205, Glu206, Tyr662, Ser630, Trp629, and Tyr547 is the site where Linagliptin (PDB: 2RGU) and most DPP4 inhibitors are located (Figure 1) [16].
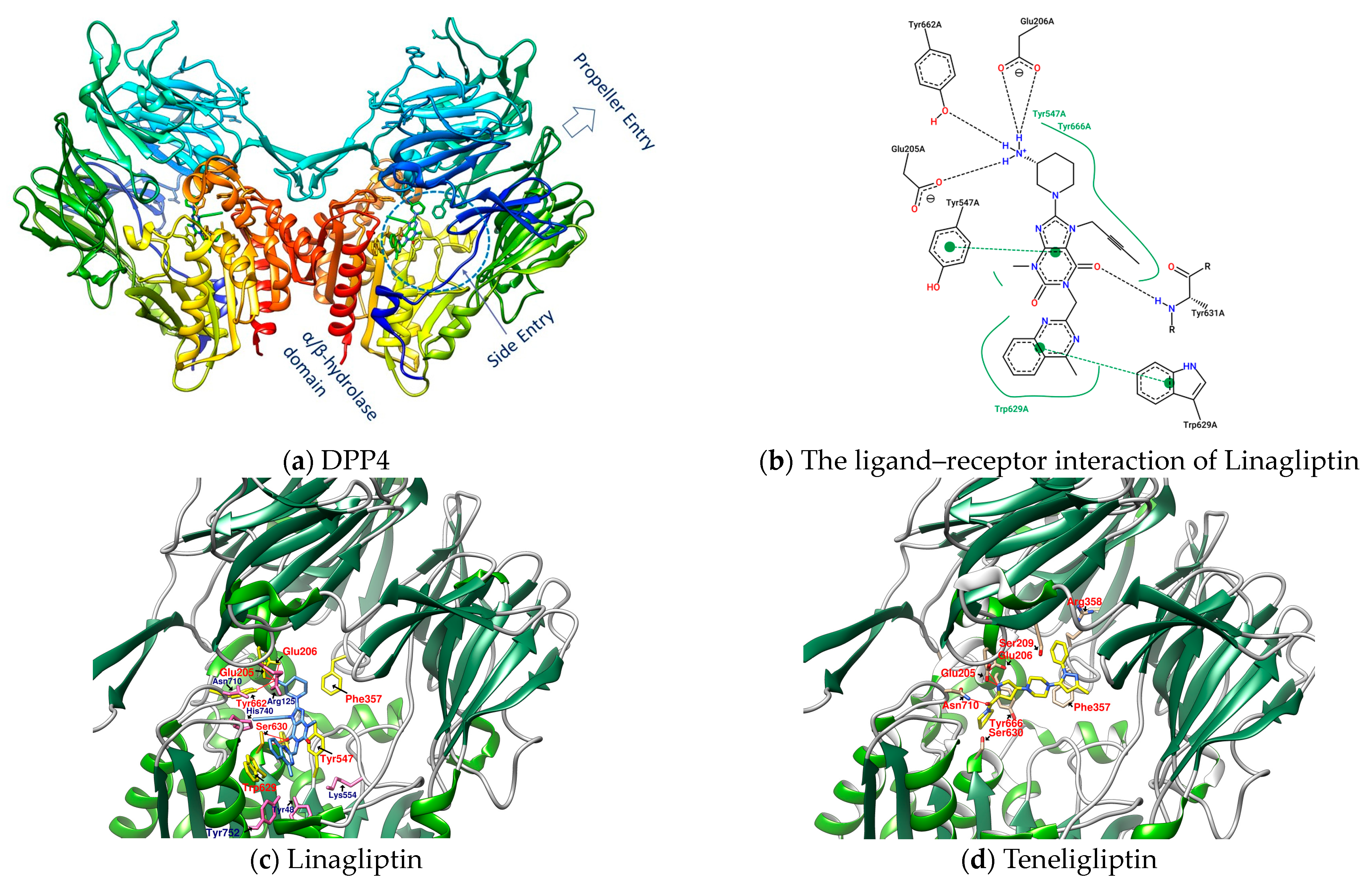
Figure 1. (a) DPP4 (PDB: 2RGU); (b) Linagliptin’s PoseView analysis; (c) Linagliptin (PDB: 2RGU) and (d) Teregliliptin (PDB: 3VJK) rendered by UCSF Chimera (red lines are hydrogen bonds).
Some inhibitors may also expand the discussion to the outer region formed by Arg125, Asn710, His740, Tyr752, Tyr48, and Lys554. There is also a design strategy for DPP4 inhibitors, in which the inhibitor approaches Lys554 in the S1′ pocket to form a salt bridge and establishes a hydrophobic interaction with Tyr547 to achieve the effect instead of binding to the amino acid residues in the catalytic center [17]. Additionally, it was observed that the (1-phenylpyrazol-5-yl) piperazine moiety of Teneligliptin (PDB: 3VJK) extends to Ser209, Phe357, and Arg358 closer to the β-propeller domain [18]. Vildagliptin (PDB: 6B1E) and Saxagliptin (PDB: 3BJM) are two cyanopyrrolidine-bearing compounds with smaller molecules than Linagliptin. The main structure of their crystals in DPP4 only occupies the more concentrated area between “Glu205, Glu206, and Ser 630” and forms a covalent bond with Ser 630 (Figure 2) [19].
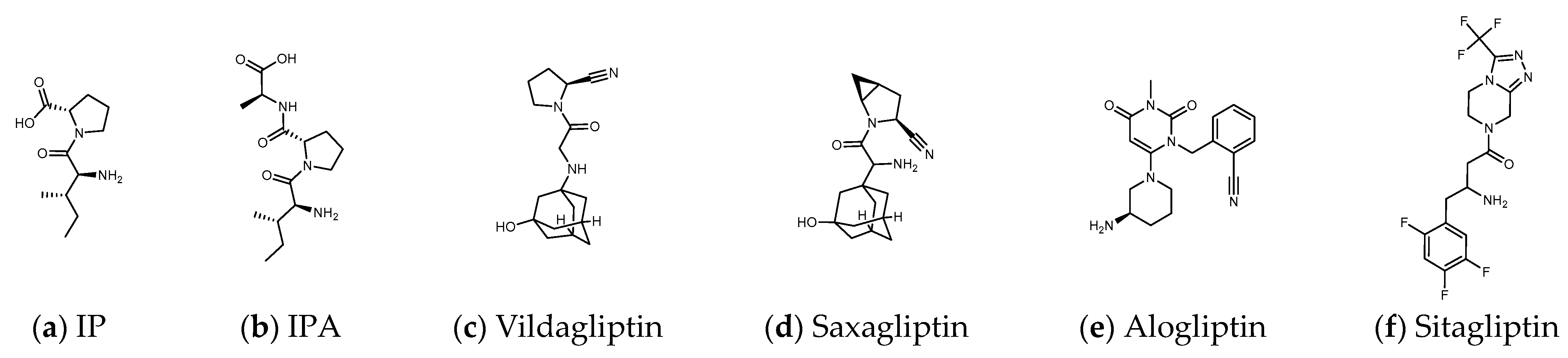
Figure 2. (a,b) Peptide DPP4 inhibitors. (c–f) Clinical DPP4 drugs. Compared with Sitagliptin, the molecular size and proline-contained structure of the dipeptides IP and IPA are closer to Vildagliptin and Saxagliptin. However, Vildagliptin and Saxagliptin are cyanopyrrolidine-bearing compounds that can form a covalent bond with DPP4. In order to establish more hydrogen bonds between the peptide and DPP4, it may be necessary to increase the length of the sequence.
4. Natural Products with Relevant Reports on Lowering Blood Sugar and Their Mechanisms
Many herbs have been shown to have the effect of regulating blood sugar and have become a choice of dietary supplements for patients with T2D. Studies have found that DPP4, PTP1B, α-glucosidase, AMPK, PPARγ, etc. are the targets that natural products may be involved in hypoglycemia [20]. These hypoglycemic natural substances include a large number of phenols, lignans, terpenes, alkaloids, protein hydrolysates, etc., but cyclic peptides (cyclopeptides) are still rare. Due to the highly charged nature of the catalytic domain of PTP1B, the oral drug design of PTP1B inhibitors remains a very challenging task [21]. Inhibition of α-glucosidase reduces the intestinal absorption of glucose and slows postprandial blood glucose rise, which is the possible hypoglycemic mechanism of many natural products [22]. However, if drug intake increases pancreatic β-cell density and improves fasting blood glucose, the pharmacological mechanism may go beyond the inhibition of α-glucosidase. DPP4 inhibition is another hypoglycemic target that may be involved. The triterpenoids quinovic acid-3β-O-β-d-glycopyranoside, lupeol, and phytosterol stigmasterol isolated from natural anti-diabetic plant Fagonia cretica L. and Hedera nepalensis K. Koch have been demonstrated to have inhibitory effects on DPP4 [23]. Flavonoids and phenols such as luteolin, apigenin, quercetin, isoquercetin, rosmarinic acid, naringin, and eriocitrin also revealed the efficacy in inhibiting the activity of DPP4 [24]. Curcumin is evaluated as an α-glucosidase and DPP4 inhibitor, and is recommended for the management of diet-induced hyperglycemia [25]. In addition, many protein hydrolysates from natural resources show potential as DPP4 inhibitors [26,27][26][27] such as LKPTPEGDL and LKPTPEGDLEIL from pepsin-treated bovine whey proteins [28]; LPQNIPPL from gouda-type cheese [29]; PPPP, GP, PP, MP, VA, MA, KA, LA, FA, AP, FP, PA, LP, VP, LL, VV, HA, IPA, and IPI from the hydrolysis of amaranth proteins [30]; and LP and IP from defatted rice bran (Figure 2) [31]. The molecular weights of these peptides vary widely. Although the spacing of Phe357, Ser209, Glu205, and Ser630 to Lys554 in DPP4 is sufficient to accommodate large molecules, small molecules can also occupy the vicinity of the catalytic center and hinder enzyme activity. This may explain why LKPTPEGDLEIL and IP both show the bioactivity of DPP4 inhibition in related studies.
5. Pseudostellaria Heterophylla, a Reported Natural Product with Hypoglycemic Effect
Pseudostellaria heterophylla (Heterophylly Falsesatarwort Root, Taizishen, or P. hetero-phylla) is rich in cyclic peptides (cyclopeptides) and is reported to be a medicinal plant with hypoglycemic effect. P. heterophylla, belonging to the Caryophyllaceae family, is known as the “ginseng of the lungs” (similar to ginseng that is good for the lungs). According to the herbal pharmacopoeia record, it is suitable for improving dry cough, loss of appetite, fatigue, mental exhaustion, and physical weakness after illness. It is also used as a nutritional supplement for children with a weak physique. In modern times, P. heterophylla is one of the important materials used in clinical Chinese compound prescriptions for improving hyperglycemia. It is rich in polysaccharides, saponins, and cyclopeptides, among which Heterophyllin B (HB) is one of its quality indicators [32,33,34][32][33][34]. Studies have already shown that P. heterophylla’s polysaccharides and saponins have the effect of lowering blood sugar [35,36][35][36]. However, a review of the hypoglycemic effect of its cyclopeptides is lacking. Heterophyllin A (HA) and HB were found in 1991 as the first cyclopeptides identified from P. heterophylla and encouraged a large number of studies on cyclopeptides in the next 30 years [37]. In the classification of cyclopeptides by NH Tan et al., the cyclopeptides from P. heterophylla including Heterophyllin A, B, C, J, and Pseudostellarin A~H were entered into the category of “Caryophyllaceae-type cyclic peptides” (CTCs) (CTCs: homo-mono-cyclopeptides formed with the peptide bonds, which include cyclic dipeptides to dodecapeptides) [38]. In 2013, PG Arnison et al.’s recommendations for a universal nomenclature suggested that plant N–C cyclic peptides lacking disulfide bonds and significantly biased toward hydrophobic amino acids could be classified as “Orbitides”. The cyclic peptides listed in CTCs and Orbitides are approximately the same, involving at least nine independent plant families. In addition to Caryophyllaceae, it also includes Annonaceae, Linaceae, and Rutaceae, etc. Some non-Caryophyllaceae-derived cyclic peptides used to be called Orbitides such as those derived from flaxseed. Studies have shown that Orbitides have many biological activities including cytotoxicity, antiplatelet, antimalarial, immune regulation, immune suppression, etc. [39]. Recently, Feng Lu et al. found that P. heterophylla’s cyclopeptides can ameliorate COPD (chronic obstructive pulmonary disease) and reduces lung inflammation via the TLR4/MyD88 pathway; moreover, the 28-days animal test of 500 mg/kg purified extract (by oral administration) showed no toxicity [40]. Recently, in related studies on DPP4 as a therapeutic target for lung diseases, it was found that DPP4 may be involved in the pathophysiology of COPD [41]. Moreover, DPP4 inhibition by Sitagliptin was found to be able to attenuate LPS-induced lung injury in mice [42]. Is it possible that the effect of P. heterophylla’s cyclopeptides on COPD is related to the inhibition of DPP4? In addition to polysaccharides and saponins, does the indicator compound HB participate in the hypoglycemic mechanism? It can be observed that the diameter of HB (octacyclic peptide) is close to the length of Linagliptin (Figure 3). Could parts of the HB ring be used to match or be close to the region in DPP4 where Linagliptin acts? Since natural ligands of DPP4 such as GLP-1 and GIP are peptides, there has been much discussion of whether protein hydrolysates are involved in DPP4 inhibition. Is it possible that the cyclopeptides of P. heterophylla are involved in the DPP4 inhibition similar to some protein hydrolysates?
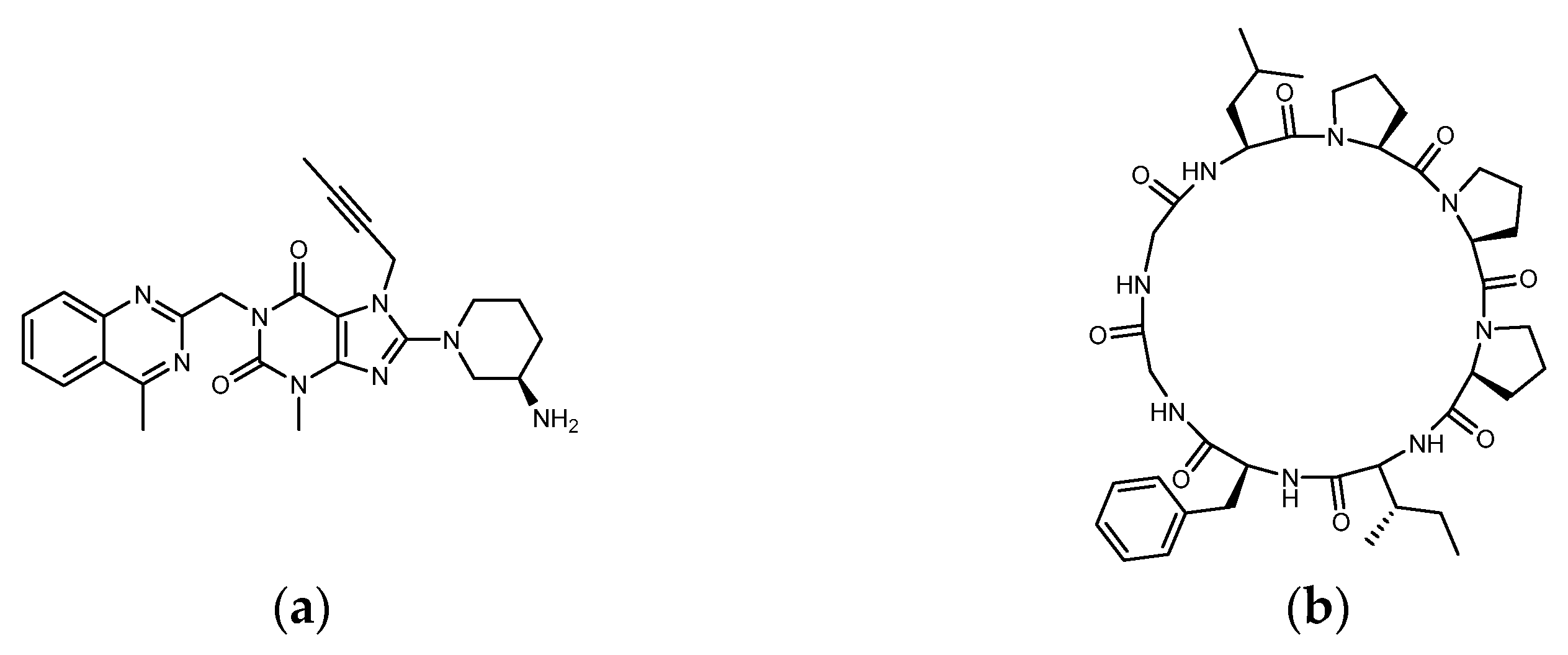
Figure 3. Structure of (a) Linagliptin and (b) Heterophyllin B.
Many natural proline-rich cyclopeptides from marine organisms have been found to be very similar in appearance to plant-derived Orbitides, but their structure may be interspersed with non-peptide elements [43]. There have been many physiological and pharmacological studies on marine cyclopeptides including anti-fertility, anti-cancer, anti-viral, etc. Many sponge-derived cyclopeptides such as the “Phakellistatin 1–19” series are well-known cytostatic compounds for the development of anticancer agents. In the “structure–activity relationship” study of Phakellistatins, it was found that because proline residues can reduce the flexibility of the backbone, the proline-rich cyclopeptides can enhance the selectivity and affinity of their receptors [43,44,45][43][44][45]. The health-oriented hydrolyzed dairy products also emphasize the role of their proline-rich peptides. The proline-rich polypeptide complex Colostrinin™, isolated from ovine colostrum, has immunoregulatory properties and shows beneficial effects on neurodegenerative diseases [46]. In addition, the aforementioned protein hydrolysates PP, PPPP, IP, and LPQNIPPL were reported to have an effect on DPP4 inhibition. The peptide sequence IFGGLPPP of Heterophyllin B is also enriched in proline. With a large number of plant-derived cyclopeptides, proline-rich may be an option to narrow down the search when screening for specific cyclopeptides of interest. Among the existing peptide drugs, cyclopeptides account for the majority due to their higher lipophilicity, higher membrane permeability, in vivo stability, and higher specificity for target receptors [47,48][47][48]. Plant cyclopeptides are currently receiving attention in many aspects such as anti-tumor, immune regulation, sedation, antibacterial, antiviral, and so on [38,39][38][39]. However, there are relatively few reports on the use of cyclopeptides for lowering blood sugar. Therapeutic drugs or dietary supplements for patients with type 2 diabetes need to consider the safety of long-term use, so it is necessary to avoid toxic species. Many marine cyclopeptides and disulfide-rich cyclotides are cytotoxic and are not suitable for development as dietary supplements. Besides, the possible changes in the configuration of cyclic peptides composed of disulfide bonds are much more complex relative to “Caryophyllaceae-Type Cyclopeptides”. Is there a chance to find other natural substances with hypoglycemic reports in the group containing CTCs, in addition to P. heterophylla? In this context, Linum usitatissimum (flaxseed) and Drymaria diandra, which are rich in CTCs, have been examined and compared with P. heterophylla under the hypoglycemic theme. They have many pharmacological effects including lowering blood sugar, and are documented for nutrition or daily health care. In addition, many of their cyclopeptides contain two proline residues [38].
6. Linum Usitatissimum, Which Is Rich in Cyclic Peptides and Has Hypoglycemic Effect Reported
Linum usitatissimum (flax, flaxseed, or linseed) contains α-linoleic acid, lignans, and cyclic peptides, etc., and is mainly used as flaxseed oil and dietary flaxseed meal. Many flaxseed studies take the term “Orbitides” to refer to its cyclopeptides. As of 2019, 39 flaxseed cyclopeptides have been isolated from flaxseed oil with a high content (more than 100 mg/100 g) [49]. According to the open-label study by Mani, U.V. et al., by supplementing 10 g of flax seed powder (FS) daily for 1 month and keeping drug intake unchanged, reductions in fasting blood glucose (FBG), glycated hemoglobin, total cholesterol, and triglyceride values were observed in the experimental group [50]. The hypoglycemic effects of flaxseed lignans have been reported in the nutrition literature. A study by A Pan et al. showed that flaxseed-derived lignan supplement improved HbA1c, but no significant difference was observed in fasting plasma glucose (FPG), insulin concentration, insulin resistance, and lipid profile [51]. There were differences in the results of hypoglycemic observations on flax lignans and FS, although the background of the experiment was not exactly the same. This led to consideration of whether there are ingredients other than lignans in flaxseed that are involved in the blood sugar-lowering mechanism and may have a broader impact. A de novo peptide sequencing study by the CycloNovo analysis method revealed that many flaxseed cyclopeptides in the human gut such as Cyclolinopeptide A, B, D, E, H, etc. [52]. The biological activity of the cyclopeptide further provides support for thinking about whether flaxseed Orbitides can reduce the enzyme activity of DPP4 and participate in the hypoglycemic mechanism.
7. Drymaria Diandra, Which Is Rich in Cyclic Peptides and Has Hypoglycemic Effect Reported
Drymaria diandra (D. diandra, Drymaria cordata Willd, or D. cordata), also known as tropical chickweed, belongs to the family Caryophyllaceae. It grows quickly in some humid and warm places in Africa, Asia, and the Americas and used as a folk medicine for anti-inflammatory, antibacterial, antipyretic, analgesic, and acute hepatitis [53,54,55][53][54][55]. Similar to P. heterophylla, D. diandra has also been reported as an antitussive effect. People in some areas use it when they have a cold or cough [56]. The main pharmacological ingredients of D. diandra are cyclopeptides, flavonoids, and alkaloids. D. diandra leaves can be sun-dried and boiled into herbal tea. In addition, it is said that the fresh leaves can be ground lightly and applied to the wound or diluted with honey water to treat fever [55,57,58][55][57][58]. Compared with the obvious symptoms of inflammation and fever, diabetes is a new concept for traditional medicine. Even in modern times, a considerable proportion of people do not know that they have diabetes. Therefore, it is not easy to find the “hypoglycemic” term in the local herbal pharmacopoeia. The ethnic groups of the Sikkim in India use D. diandra for various diseases including diabetes. D. diandra has become one of the few regional herbal medicines that can enter the field of contemporary diabetes research. S Patra et al. tried to treat diabetic rats with a D. diandra (D. cordata) methanol extract (DCME) to observe changes in various physiological indicators. DCME is still safe at an oral dose of 2000 mg/kg. Compared with the diabetes group, HbA1c, FBG, and lipid profiles in the DCME group were reduced, and the β cell density was improved in a dose-dependent manner. Their studies speculated that the α-glucosidase inhibitory activity of DCME and the antioxidant properties of flavonoids and alkaloids are responsible for the improvement of type 2 diabetes [59]. Since experimental values overlap with the effect of DPP4 inhibition, and the cyclopeptides “Diandrine A–D” are present in the methanol extract according to the early identification by PW Hsieh et al., is it possible that cyclopeptides from D. diandra are also involved in the hypoglycemic mechanism based on incretin [57]?
8. Can Linear Precursors of Heterophyllin B “IFGGLPPP” Participate in DPP4 Inhibition?
Numerous protein hydrolysates have shown the effect of DPP4 inhibition. However, the process of identifying functional peptide sequences from hydrolysis, isolation, purification, and bioassay to mass spectrometry analysis is difficult and time-consuming. Two linear precursors of Heterophyllin B (HB), “GGLPPPIF”, and “IFGGLPPP” have been previously reported. The sequence of IFGGLPPP was verified by precursor gene (prePhHB) screening and in vitro and in vivo experiments, which were suggested to be more likely to be the precursor peptide of HB, later [33]. Is it possible that the linear peptide IFGGLPPP has a favorable binding affinity for DPP4? Additionally, can IFGGLPPP be modified to obtain more samples for comparative studies? For example, inserting other fragments such as PPPP, FP, WP, and PY into existing linear peptides, or changing the sequence of local fragments. DPP4 cleaves dipeptides such as Xaa-Proline or Xaa-Alanine (also includes Xaa-Gly, Xaa-Ser, Xaa-Val, etc., but mainly Xaa-Pro) from the N-terminus of the polypeptide, where Xaa stands for any amino acid. The sequence to be cleaved by DPP4 can be any amino acid except Proline at the third position [11]. Cleavage by DPP4 should be avoided when designing or developing linear peptides as DPP4 inhibitors. Since the samples to be explored contain a large number of cyclopeptides from P. heterophylla, flaxseed, and D. diandra as well as a series of IFGGLPPP-derived peptides, molecular modeling provides a feasible method for preliminary screening of potential cases. In the future, molecules with good DPP4 binding affinity screened by docking can be isolated, purified, or synthesized for more in vitro or in vivo studies.
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843.
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59.
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From Pre-Diabetes to Diabetes: Diagnosis, Treatments and Translational Research. Medicina 2019, 55, 546.
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157.
- Kim, W.; Egan, J.M. The role of incretins in glucose homeostasis and diabetes treatment. Pharm. Rev. 2008, 60, 470–512.
- Godinho, R.; Mega, C.; Teixeira-de-Lemos, E.; Carvalho, E.; Teixeira, F.; Fernandes, R.; Reis, F. The Place of Dipeptidyl Peptidase-4 Inhibitors in Type 2 Diabetes Therapeutics: A “Me Too” or “the Special One” Antidiabetic Class? J. Diabetes Res. 2015, 2015, 806979.
- Müller, T.D.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.J.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130.
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80.
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes–state-of-the-art. Mol. Metab. 2021, 46, 101102.
- Gallwitz, B. Clinical use of DPP-4 inhibitors. Front. Endocrinol. 2019, 10, 389.
- Klemann, C.; Wagner, L.; Stephan, M.; von Hörsten, S. Cut to the chase: A review of CD26/dipeptidyl peptidase-4′s (DPP4) entanglement in the immune system. Clin. Exp. Immunol. 2016, 185, 1–21.
- Scott, L.J. Sitagliptin: A Review in Type 2 Diabetes. Drugs 2017, 77, 209–224.
- Engel, M.; Hoffmann, T.; Wagner, L.; Wermann, M.; Heiser, U.; Kiefersauer, R.; Huber, R.; Bode, W.; Demuth, H.U.; Brandstetter, H. The crystal structure of dipeptidyl peptidase IV (CD26) reveals its functional regulation and enzymatic mechanism. Proc. Natl. Acad. Sci. USA 2003, 100, 5063–5068.
- Röhrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in diabetes. Front. Immunol. 2015, 6, 386.
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019.
- Eckhardt, M.; Langkopf, E.; Mark, M.; Tadayyon, M.; Thomas, L.; Nar, H.; Pfrengle, W.; Guth, B.; Lotz, R.; Sieger, P.; et al. 8-(3-(R)-aminopiperidin-1-yl)-7-but-2-ynyl-3-methyl-1-(4-methyl-quinazolin-2-ylmethyl)-3,7-dihydropurine-2,6-dione (BI 1356), a highly potent, selective, long-acting, and orally bioavailable DPP-4 inhibitor for the treatment of type 2 diabetes. J. Med. Chem. 2007, 50, 6450–6453.
- Maezaki, H.; Tawada, M.; Yamashita, T.; Banno, Y.; Miyamoto, Y.; Yamamoto, Y.; Ikedo, K.; Kosaka, T.; Tsubotani, S.; Tani, A.; et al. Design of potent dipeptidyl peptidase IV (DPP-4) inhibitors by employing a strategy to form a salt bridge with Lys554. Bioorg. Med. Chem. Lett. 2017, 27, 3565–3571.
- Yoshida, T.; Akahoshi, F.; Sakashita, H.; Kitajima, H.; Nakamura, M.; Sonda, S.; Takeuchi, M.; Tanaka, Y.; Ueda, N.; Sekiguchi, S. Discovery and preclinical profile of teneligliptin (3- pyrrolidin-2-ylcarbonyl] thiazolidine): A highly potent, selective, long-lasting and orally active dipeptidyl peptidase IV inhibitor for the treatment of type 2 diabetes. Bioorganic Med. Chem. 2012, 20, 5705–5719.
- Berger, J.P.; SinhaRoy, R.; Pocai, A.; Kelly, T.M.; Scapin, G.; Gao, Y.D.; Pryor, K.A.D.; Wu, J.K.; Eiermann, G.J.; Xu, S.S.; et al. A comparative study of the binding properties, dipeptidyl peptidase-4 (DPP-4) inhibitory activity and glucose-lowering efficacy of the DPP-4 inhibitors alogliptin, linagliptin, saxagliptin, sitagliptin and vildagliptin in mice. Endocrinol. Diabetes Metab. 2018, 1, e00002.
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994.
- Jiang, C.S.; Liang, L.F.; Guo, Y.W. Natural products possessing protein tyrosine phosphatase 1B (PTP1B) inhibitory activity found in the last decades. Acta Pharm. Sin. 2012, 33, 1217–1245.
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738.
- Saleem, S.; Jafri, L.; Haq, I.U.; Chang, L.C.; Calderwood, D.; Green, B.D.; Mirza, B. Plants Fagonia cretica L. and Hedera nepalensis K. Koch contain natural compounds with potent dipeptidyl peptidase-4 (DPP-4) inhibitory activity. J. Ethnopharmacol. 2014, 156, 26–32.
- Gao, Y.; Zhang, Y.; Zhu, J.; Li, B.; Li, Z.; Zhu, W.; Shi, J.; Jia, Q.; Li, Y. Recent progress in natural products as DPP-4 inhibitors. Future Med. Chem. 2015, 7, 1079–1089.
- Cao, W.; Chen, X.; Chin, Y.; Zheng, J.; Lim, P.E.; Xue, C.; Tang, Q. Identification of curcumin as a potential α-glucosidase and dipeptidyl-peptidase 4 inhibitor: Molecular docking study, in vitro and in vivo biological evaluation. J. Food Biochem. 2021, 46, e13686.
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422.
- Oseguera-Toledo, M.E.; de Mejía, E.G.; Reynoso-Camacho, R.; Cardador-Martínez, A.; Amaya-Llano, S.L. Proteins and bioactive peptides. Nutrafoods 2014, 13, 147–157.
- Lacroix, I.M.; Li-Chan, E.C. Isolation and characterization of peptides with dipeptidyl peptidase-IV inhibitory activity from pepsin-treated bovine whey proteins. Peptides 2014, 54, 39–48.
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda.a-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30.
- Velarde-Salcedo, A.J.; Barrera-Pacheco, A.; Lara-González, S.; Montero-Morán, G.M.; Díaz-Gois, A.; González de Mejia, E.; Barba de la Rosa, A.P. In vitro inhibition of dipeptidyl peptidase IV by peptides derived from the hydrolysis of amaranth (Amaranthus hypochondriacus L.) proteins. Food Chem. 2013, 136, 758–764.
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802.
- Hu, D.J.; Shakerian, F.; Zhao, J.; Li, S.P. Chemistry, pharmacology and analysis of Pseudostellaria heterophylla: A mini-review. Chin. Med. 2019, 14, 21.
- Zheng, W.; Zhou, T.; Li, J.; Jiang, W.; Zhang, J.; Xiao, C.; Wei, D.; Yang, C.; Xu, R.; Gong, A.; et al. The Biosynthesis of Heterophyllin B in Pseudostellaria heterophylla From prePhHB-Encoded Precursor. Front. Plant. Sci. 2019, 10, 1259.
- Su, K.; Zhu, F.; Guo, L.; Zhu, Y.; Li, W.; Xiong, X. Retrospective study on Professor Zhongying Zhou’s experience in Traditional Chinese Medicine treatment on diabetic nephropathy. J. Tradit. Chin. Med. 2013, 33, 262–267.
- Hu, J.; Pang, W.; Chen, J.; Bai, S.; Zheng, Z.; Wu, X. Hypoglycemic effect of polysaccharides with different molecular weight of Pseudostellaria heterophylla. BMC Complement. Altern. Med. 2013, 13, 267.
- Cui, J.; Li, Y.W.; Jia, N.; Song, X.M.; Duan, J.L.; Weng, Y.; Guan, Y.; Zhou, D.; Wang, L.; Xi, M.M.; et al. Insulin-secretagogue activity of eleven plant extracts and twelve pure compounds isolated from Aralia taibaiensis. Life Sci. 2013, 92, 131–136.
- Zhou, J.; Tan, N. Application of a new TLC chemical method for detection of cyclopeptides in plants. Chin. Sci. Bull. 2000, 45, 1825–1831.
- Tan, N.-H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895.
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160.
- Lu, F.; Yang, H.; Lin, S.D.; Zhao, L.; Jiang, C.; Chen, Z.B.; Liu, Y.Y.; Kan, Y.J.; Hu, J.; Pang, W.S. Cyclic Peptide Extracts Derived From Pseudostellaria heterophylla Ameliorates COPD via Regulation of the TLR4/MyD88 Pathway Proteins. Front. Pharm. 2020, 11, 850.
- Zou, H.; Zhu, N.; Li, S. The emerging role of dipeptidyl-peptidase-4 as a therapeutic target in lung disease. Expert Opin. Ther. Targets 2020, 24, 147–153.
- Kawasaki, T.; Chen, W.; Htwe, Y.M.; Tatsumi, K.; Dudek, S.M. DPP4 inhibition by sitagliptin attenuates LPS-induced lung injury in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 315, L834–L845.
- Fang, W.Y.; Dahiya, R.; Qin, H.L.; Mourya, R.; Maharaj, S. Natural Proline-Rich Cyclopolypeptides from Marine Organisms: Chemistry, Synthetic Methodologies and Biological Status. Mar. Drugs 2016, 14, 194.
- Meli, A.; Tedesco, C.; Della Sala, G.; Schettini, R.; Albericio, F.; De Riccardis, F.; Izzo, I. Phakellistatins: An Underwater Unsolved Puzzle. Mar. Drugs 2017, 15, 78.
- Zhao, L.; Wu, J.; Bao, Y.; Jiang, S.; Wang, Z.; Jin, Y.; Qu, F. The conventional turns rather than irregular γ-/β-turn secondary structures accounting for the antitumor activities of cyclic peptide Phakellistatin 6 analogs. Tetrahedron 2020, 76, 130881.
- Janusz, M.; Zabłocka, A. Colostral proline-rich polypeptides--immunoregulatory properties and prospects of therapeutic use in Alzheimer’s disease. Curr. Alzheimer Res. 2010, 7, 323–333.
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29.
- Henninot, A.; Collins, J.C.; Nuss, J.M. The Current State of Peptide Drug Discovery: Back to the Future? J. Med. Chem. 2018, 61, 1382–1414.
- Shim, Y.Y.; Song, Z.; Jadhav, P.D.; Reaney, M.J. Orbitides from flaxseed (Linum usitatissimum L.): A comprehensive review. Trends Food Sci. Technol. 2019, 93, 197–211.
- Mani, U.V.; Mani, I.; Biswas, M.; Kumar, S.N. An open-label study on the effect of flax seed powder (Linum usitatissimum) supplementation in the management of diabetes mellitus. J. Diet. Suppl. 2011, 8, 257–265.
- Pan, A.; Sun, J.; Chen, Y.; Ye, X.; Li, H.; Yu, Z.; Wang, Y.; Gu, W.; Zhang, X.; Chen, X.; et al. Effects of a flaxseed-derived lignan supplement in type 2 diabetic patients: A randomized, double-blind, cross-over trial. PLoS ONE 2007, 2, e1148.
- Behsaz, B.; Mohimani, H.; Gurevich, A.; Prjibelski, A.; Fisher, M.; Vargas, F.; Smarr, L.; Dorrestein, P.C.; Mylne, J.S.; Pevzner, P.A. De Novo Peptide Sequencing Reveals Many Cyclopeptides in the Human Gut and Other Environments. Cell Syst. 2020, 10, 99–108.e105.
- Nono, N.R.; Nzowa, K.L.; Barboni, L.; Tapondjou, A.L. Drymaria cordata (Linn.) Willd (Caryophyllaceae): Ethnobotany, Pharmacology and Phytochemistry. Adv. Biol. Chem. 2014, 2014.
- Barua, C.C.; Roy, J.D.; Buragohain, B.; Barua, A.G.; Borah, P.; Lahkar, M. Analgesic and anti-nociceptive activity of hydroethanolic extract of Drymaria cordata Willd. Indian J. Pharm. 2011, 43, 121–125.
- Ding, Z.; Zhou, J.; Tan, N.; Teng, R. Two new cyclic peptides from Drymaria diandra. Planta Med. 2000, 66, 386–388.
- Mukherjee, P.K.; Saha, K.; Bhattacharya, S.; Giri, S.; Pal, M.; Saha, B. Studies on antitussive activity of Drymaria cordata Willd. (Caryophyllaceae). J. Ethnopharmacol. 1997, 56, 77–80.
- Hsieh, P.W.; Chang, F.R.; Wu, C.C.; Wu, K.Y.; Li, C.M.; Wang, W.Y.; Gu, L.C.; Wu, Y.C. Selective Inhibition of Collagen-Induced Platelet Aggregation by a Cyclic Peptide from Drymaria diandra. Helv. Chim. Acta 2004, 87, 57–66.
- Hsieh, P.W.; Chang, F.R.; Lee, K.H.; Hwang, T.L.; Chang, S.M.; Wu, Y.C. A new anti-HIV alkaloid, drymaritin, and a new C-glycoside flavonoid, diandraflavone, from Drymaria diandra. J. Nat. Prod. 2004, 67, 1175–1177.
- Patra, S.; Bhattacharya, S.; Bala, A.; Haldar, P.K. Antidiabetic effect of Drymaria cordata leaf against streptozotocin-nicotinamide-induced diabetic albino rats. J. Adv. Pharm. Technol. Res. 2020, 11, 44–52.
More