Ferroptosis, an iron and reactive oxygen species (ROS)-dependent non-apoptotic type of regulated cell death, is characterized by a massive iron overload and peroxidation of polyunsaturated fatty acids (PUFAs), which finally results in cell death. Recent studies suggest that ferroptosis can influence carcinogenesis negatively and therefore may be used as a novel anti-cancer strategy. Hepatocellular carcinoma (HCC) is a deadly malignancy with poor chances of survival and is the second leading cause of cancer deaths worldwide. Diagnosis at an already late stage and general resistance to current therapies may be responsible for the dismal outcome. As the liver acts as a key factor in iron metabolism, ferroptosis is shown to play an important role in HCC carcinogenesis and, more importantly, may hold the potential to eradicate HCC.
- ferroptosis
- iron
- hepatocellular carcinoma
- drug development
- combinatory treatment
1. Introduction
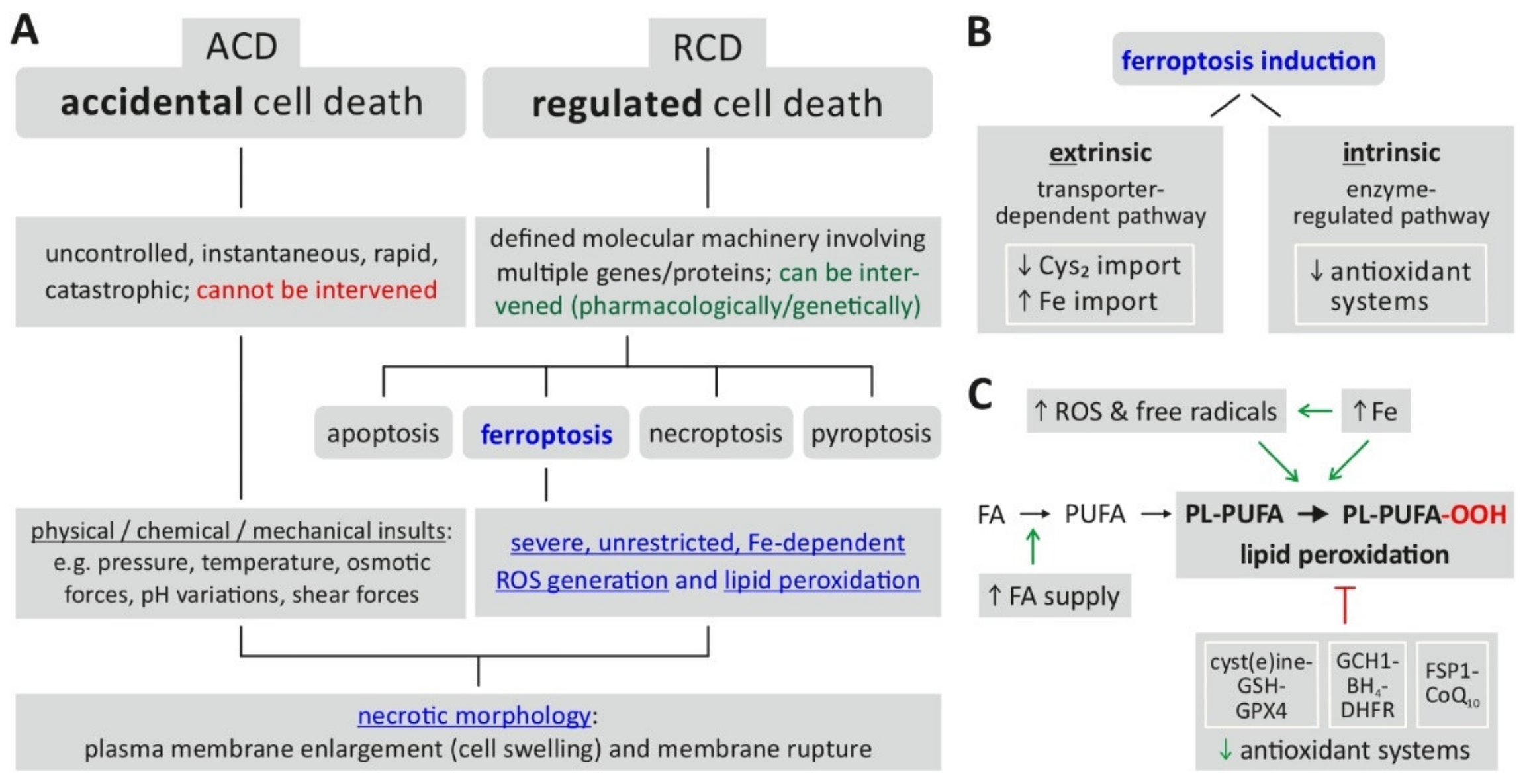
2. The Translation of Ferroptosis into Clinical HCC Practice
This chapter deals with the issue of how the basic knowledge of (pharmacologically) induced ferroptosis could be translated into “daily” clinical practice for HCC patients. In particular, possible therapeutic scenarios for HCC treatment based on the combination of ferroptosis induction with standard chemotherapy, targeted and immunotherapy as well as local ablative techniques and radiotherapy will be presented on the basis of “pre-clinical” experiments or first clinical trials.2.1. Ferroptosis Scoring System
| Year | Database(s) (Dataset) | Basic Cluster Description | Predictive and/or Prognostic Aspects of the Ferroptosis Cluster | Ref |
|---|---|---|---|---|
| 2021 | TCGA: LIHC GEO: GSE76427 |
Ferrcluster A: “Olfactory transduction” and “cardiac music contraction”. Ferrcluster B: “mTOR signaling pathway” and “neurotrophin signaling pathway”. Ferrcluster C: “adipokine signaling pathway”, “tyrosine metabolism” and “PPAR signaling pathway” |
Ferrcluster B: Overall survival ↓ High ferrscore group: Survival ↓, Programmed cell death 1 (PD-1) mRNA expression ↑, efficacy of PD-1 or PD-1 plus CTLA4 (cytotoxic T-lymphocyte associated protein 4) inhibitors ↓. |
[14] |
| 2021 | TCGA ICGC |
Ferroptosis-H and Ferroptosis-L: According to ferroptosis gene expression and methylation |
Ferroptosis-H: Overall and disease-specific survival ↓ | [15] |
| 2021 | GEO TCGA ICGC |
C1: Metabolism low, immunity high subtype. C2: Metabolism high, immunity low subtype. |
C1: Prognosis ↓ C1: Patients with clinical characteristics such as younger, female, advanced stage, higher grade, vascular invasion. |
[16] |
| 2020 | GEO: GSE14520/GPL3921 TCGA |
Low and high group: Comprehensive index of ferroptosis and immune status (CIFI). | High CIFI: Prognosis ↓ | [17] |
| 2021 | TCGA | Low-risk and high-risk groups: 2 ferroptosis-related mRNAs and ferroptosis-related lncRNAs | Higher risk group: Prognosis ↓ Higher risk group: Differences of tumor microenvironment, immune cell infiltration as well as tumor-related pathways |
[18] |
| 2021 a | TCGA-CHOL GEO: GSE107943 EMBL-EBI: E-MTAB-6389 |
Low and high group: Ferroptosis-related weighted coexpression gene network and model construction. | Higher risk group: Prognosis ↓ | [19] |
2.2. Nanoparticles and Exosomes
2.3. Long Noncoding RNAs/miRNA
3. Conclusions
References
- Liang, C.; Zhang, X.; Yang, M.; Dong, X. Recent Progress in Ferroptosis Inducers for Cancer Therapy. Adv. Mater. 2019, 31, e1904197.
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364.
- Hassannia, B.; Vandenabeele, P.; Berghe, T.V. Targeting Ferroptosis to Iron Out Cancer. Cancer Cell 2019, 35, 830–849.
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072.
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541.
- Chen, X.; Comish, P.B.; Tang, D.; Kang, R. Characteristics and Biomarkers of Ferroptosis. Front. Cell Dev. Biol. 2021, 9, 637162.
- Dolma, S.; Lessnick, S.L.; Hahn, W.C.; Stockwell, B.R. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells. Cancer Cell 2003, 3, 285–296.
- Yang, W.S.; Stockwell, B.R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem. Biol. 2008, 15, 234–245.
- Tang, D.; Kroemer, G. Ferroptosis. Curr. Biol. 2020, 30, R1292–R1297.
- Conrad, M.; Angeli, J.P.F.; Vandenabeele, P.; Stockwell, B.R. Regulated necrosis: Disease relevance and therapeutic opportunities. Nat. Rev. Drug Discov. 2016, 15, 348–366.
- Vanden Berghe, T.; Linkermann, A.; Jouan-Lanhouet, S.; Walczak, H.; Vandenabeele, P. Regulated necrosis: The expanding network of non-apoptotic cell death pathways. Nat. Rev. Mol. Cell Biol. 2014, 15, 135–147.
- Chen, X.; Kang, R.; Kroemer, G.; Tang, D. Broadening horizons: The role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 2021, 18, 280–296.
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282.
- Gao, L.; Xue, J.; Liu, X.; Cao, L.; Wang, R.; Lei, L. A scoring model based on ferroptosis genes for prognosis and immunotherapy response prediction and tumor microenvironment evaluation in liver hepatocellular carcinoma. Aging 2021, 13, 24866–24881.
- Deng, T.; Hu, B.; Jin, C.; Tong, Y.; Zhao, J.; Shi, Z.; Zhang, T.; Deng, L.; Sun, Z.; Chen, G.; et al. A novel ferroptosis phenotype-related clinical-molecular prognostic signature for hepatocellular carcinoma. J. Cell. Mol. Med. 2021, 25, 6618–6633.
- Liu, Z.; Wang, L.; Liu, L.; Lu, T.; Jiao, D.; Sun, Y.; Han, X. The Identification and Validation of Two Heterogenous Subtypes and a Risk Signature Based on Ferroptosis in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 619242.
- Liu, Y.; Zhang, X.; Zhang, J.; Tan, J.; Li, J.; Song, Z. Development and Validation of a Combined Ferroptosis and Immune Prognostic Classifier for Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2020, 8, 596679.
- Chen, Z.-A.; Tian, H.; Yao, D.-M.; Zhang, Y.; Feng, Z.-J.; Yang, C.-J. Identification of a Ferroptosis-Related Signature Model Including mRNAs and lncRNAs for Predicting Prognosis and Immune Activity in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 738477.
- Zhang, Z.-J.; Huang, Y.-P.; Li, X.-X.; Liu, Z.-T.; Liu, K.; Deng, X.-F.; Xiong, L.; Zou, H.; Wen, Y. A Novel Ferroptosis-Related 4-Gene Prognostic Signature for Cholangiocarcinoma and Photodynamic Therapy. Front. Oncol. 2021, 11, 747445.
- Ocker, M.; Mayr, C.; Kiesslich, T.; Stintzing, S.; Neureiter, D. Immunmodulatory Treatment Strategies of Hepatocellular Carcinoma: From Checkpoint Inhibitors Now to an Integrated Approach in the Future. Cancers 2021, 13, 1558.
- Feng, J.; Lu, P.-Z.; Zhu, G.-Z.; Hooi, S.C.; Wu, Y.; Huang, X.-W.; Dai, H.-Q.; Chen, P.-H.; Li, Z.-J.; Su, W.-J.; et al. ACSL4 is a predictive biomarker of sorafenib sensitivity in hepatocellular carcinoma. Acta Pharmacol. Sin. 2020, 42, 160–170.
- Neureiter, D.; Stintzing, S.; Kiesslich, T.; Ocker, M. Hepatocellular carcinoma: Therapeutic advances in signaling, epigenetic and immune targets. World J. Gastroenterol. 2019, 25, 3136–3150.
- Zhou, Q.-M.; Lu, Y.-F.; Zhou, J.-P.; Yang, X.-Y.; Wang, X.-J.; Yu, J.-N.; Du, Y.-Z.; Yu, R.-S. Self-amplification of oxidative stress with tumour microenvironment-activatable iron-doped nanoplatform for targeting hepatocellular carcinoma synergistic cascade therapy and diagnosis. J. Nanobiotechnol. 2021, 19, 361.
- Tang, H.; Chen, D.; Lia, C.; Zhengc, C.; Wud, X.; Zhanga, Y.; Songc, Q.; Feic, W. Dual GSH-exhausting sorafenib loaded manganese-silica nanodrugs for inducing the ferroptosis of hepatocellular carcinoma cells. Int. J. Pharm. 2019, 572, 118782.
- Xu, Q.; Zhan, G.; Zhang, Z.; Yong, T.; Yang, X.; Gan, L. Manganese porphyrin-based metal-organic framework for synergistic sonodynamic therapy and ferroptosis in hypoxic tumors. Theranostics 2021, 11, 1937–1952.
- Tian, H.; Zhao, S.; Nice, E.C.; Huang, C.; He, W.; Zou, B.; Lin, J. A cascaded copper-based nanocatalyst by modulating glutathione and cyclooxygenase-2 for hepatocellular carcinoma therapy. J. Colloid Interface Sci. 2021, 607, 1516–1526.
- Ou, W.; Mulik, R.S.; Anwar, A.; McDonald, J.G.; He, X.; Corbin, I.R. Low-density lipoprotein docosahexaenoic acid nanoparticles induce ferroptotic cell death in hepatocellular carcinoma. Free Radic. Biol. Med. 2017, 112, 597–607.
- Du, J.; Wan, Z.; Wang, C.; Lu, F.; Wei, M.; Wang, D.; Hao, Q. Designer exosomes for targeted and efficient ferroptosis induction in cancer via chemo-photodynamic therapy. Theranostics 2021, 11, 8185–8196.
- Huang, A.; Li, T.; Xie, X.; Xia, J. Computational Identification of Immune- and Ferroptosis-Related LncRNA Signature for Prognosis of Hepatocellular Carcinoma. Front. Mol. Biosci. 2021, 8, 759173.
- Bai, T.; Liang, R.; Zhu, R.; Wang, W.; Zhou, L.; Sun, Y. MicroRNA-214-3p enhances erastin-induced ferroptosis by targeting ATF4 in hepatoma cells. J. Cell. Physiol. 2020, 235, 5637–5648.
- Wang, L.; Ge, X.; Zhang, Z.; Ye, Y.; Zhou, Z.; Li, M.; Yan, H.; Wu, L.; Bai, Q.; Li, J.; et al. Identification of a Ferroptosis-Related Long Noncoding RNA Prognostic Signature and Its Predictive Ability to Immunotherapy in Hepatocellular Carcinoma. Front. Genet. 2021, 12, 682082.
- Wu, H.; Liu, T.; Qi, J.; Qin, C.; Zhu, Q. Four Autophagy-Related lncRNAs Predict the Prognosis of HCC through Coexpression and ceRNA Mechanism. BioMed Res. Int. 2020, 2020, 3801748.
- Hou, W.; Xie, Y.; Song, X.; Sun, X.; Lotze, M.T.; Zeh, H.J., 3rd; Kang, R.; Tang, D. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy 2016, 12, 1425–1428.
- Lyu, N.; Zeng, Y.; Kong, Y.; Chen, Q.; Deng, H.; Ou, S.; Bai, Y.; Tang, H.; Wang, X.; Zhao, M. Ferroptosis is involved in the progression of hepatocellular carcinoma through the circ0097009/miR-1261/SLC7A11 axis. Ann. Transl. Med. 2021, 9, 675.
- Liu, Z.; Wang, Q.; Wang, X.; Xu, Z.; Wei, X.; Li, J. Circular RNA cIARS regulates ferroptosis in HCC cells through interacting with RNA binding protein ALKBH5. Cell Death Discov. 2020, 6, 3801748.
- Zhang, Z.; Yao, Z.; Wang, L.; Ding, H.; Shao, J.; Chen, A.; Zhang, F.; Zheng, S. Activation of ferritinophagy is required for the RNA-binding protein ELAVL1/HuR to regulate ferroptosis in hepatic stellate cells. Autophagy 2018, 14, 2083–2103.
- Li, B.; Yang, L.; Peng, X.; Fan, Q.; Wei, S.; Yang, S.; Li, X.; Jin, H.; Wu, B.; Huang, M.; et al. Emerging mechanisms and applications of ferroptosis in the treatment of resistant cancers. Biomed. Pharmacother. 2020, 130, 110710.
- Lee, J.-Y.; Kim, W.; Bae, K.-H.; Lee, S.; Lee, E.-W. Lipid Metabolism and Ferroptosis. Biology 2021, 10, 184.
- Jia, M.; Zhang, H.; Qin, Q.; Hou, Y.; Zhang, X.; Chen, D.; Zhang, H.; Chen, Y. Ferroptosis as a new therapeutic opportunity for nonviral liver disease. Eur. J. Pharmacol. 2021, 908, 174319.
- Zhang, X.; Xing, X.; Liu, H.; Feng, J.; Tian, M.; Chang, S.; Liu, P.; Zhang, H. Ionizing radiation induces ferroptosis in granulocyte-macrophage hematopoietic progenitor cells of murine bone marrow. Int. J. Radiat. Biol. 2020, 96, 584–595.