Marginal zone (MZ) B-cells are innate-like, and possess a polyreactive B-cell receptor (BCR) and several pattern recognition receptors (PRR). They are known to generate low-affinity first-line antibody responses against invading pathogens such as encapsulated bacteria. Unfortunately, deregulations affecting MZ B-cell populations have been reported in the context of Human Immunodeficiency Virus (HIV) and other chronic inflammatory conditions.
- marginal zone (MZ) B-cells
1. Introduction
Marginal zone (MZ) B-cells are innate-like, and possess a polyreactive B-cell receptor (BCR) and several pattern recognition receptors (PRR) [1][2]. They are known to generate low-affinity first-line antibody responses against invading pathogens such as encapsulated bacteria [3]. Unfortunately, deregulations affecting MZ B-cell populations have been reported in the context of Human Immunodeficiency Virus (HIV) and other chronic inflammatory conditions [2][4][5]. In here, the MZ B-cell ontogeny and antibody responses will only be briefly discuss, as these topics have been reviewed elsewhere and are beyond the scope [1][2][3][6]. Examining the regulatory capacities of MZ and other B-cell populations sharing similar features will be concentrated. The importance of the B-cell activating factor (BAFF) and its analog A Proliferation-Inducing Ligand (APRIL) in shaping the MZ B-cell pool and Breg profile will be discussed. The deregulation of MZ B-cell populations and development of MZ lymphomas (MZL) in the context of HIV and other inflammatory diseases will also be addressed. Lastly, the possible therapeutic avenues that could be deployed to restore MZ B-cell immune competence will also be talked about.
2. Ontogeny of MZ B-Cells
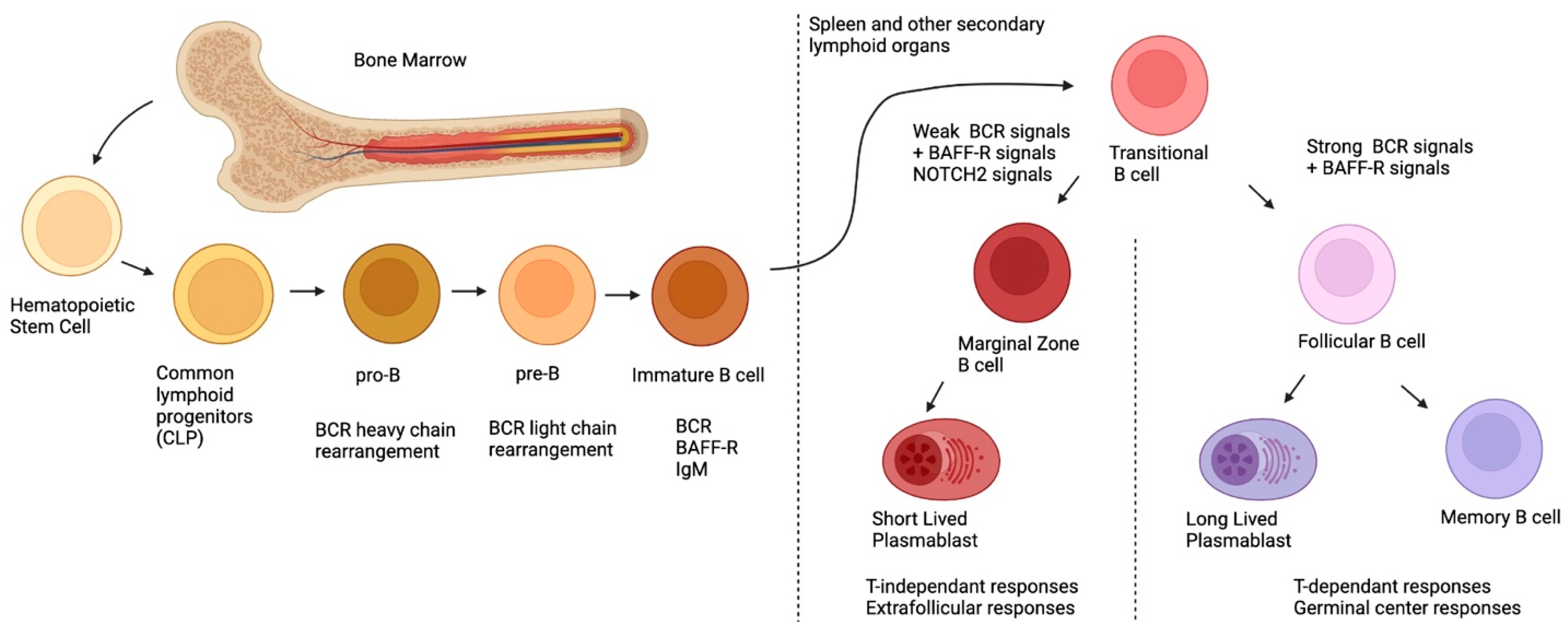
3. MZ B-Cells and Their Antibody Responses
4. MZ B-Cell Populations and Their Regulatory Potential
| Species | Population | Phenotype | Mechanism of Suppression | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mouse | B10 | CD19 + CD5 + CD1d | hi | IL-10 | [51][52] | ||||||
| MZ B-cells | IgM | hi | IgD | lo | CD21 | hi | CD23-CD1d | hi | IL-10 | [53] | |
| T2-MZP | B220 + CD21 | hi | CD1d | hi | IgM | hi | CD23+ | IL-10 | [54] | ||
| B1a | CD90-CD5+ | IL-10 | [55] | ||||||||
| Plasma cells | CD19 + CD138 + IgM+ | IL10, IL-35 | [56] | ||||||||
| Plasmablasts | CD138 + CD44 | hi | IL-10 | [57] | |||||||
| Tim-1 + B-cells | CD19 + Tim-1+ | IL-10 | [58] | ||||||||
| IL-35-Bregs | CD5 + CD1d | hi | FcγIib | hi | IL-35 | [59] | |||||
| GITRL + B-cells | - | GITRL | [60] | ||||||||
| Killer B-cells | CD19 + CD5 + FasL+ | FasL, TGF-β | [61][62] | ||||||||
| PD-L1 | hi | B-cells | CD19 + PD-L1 | hi | PD-L1 | [63] | |||||
| - | B220 + CD39 + CD73+ | ADO, CD39 + CD73 + Extracellular vesicules | |||||||||
| hi | |||||||||||
| CD27+ | |||||||||||
| IL-10 | [ | 69 | ] | ||||||||
| Br1 | CD25 | hi | CD71hi CD73 | lo | IL-10 | [70] | |||||
| TIM1 + B-cells | CD19 + TIM1+ | IL-10 | [71] | ||||||||
| Plasmablast | CD19 | lo | CD27 | hi | CD38 | hi | IL-10 | [72][73] | |||
| IgA + B-cells | CD19 + IgA+ | IL-10, PD-L1 | [74] | ||||||||
| Exhausted B-cells | CD19 + CD95+ | CD95 | [75] | ||||||||
| Killer B-cells | CD19 + CD38 + IgM + FasL+ | FasL | [76] | ||||||||
| PD-L1 B-cells | CD19 + PD-L1+ | PD-L1 | [62] | ||||||||
| CD39 | high | CD19 + CD39 | high | CD73+ | ADO | [77] | |||||
| iBreg | - | TGF-β, IDO | [78] |
| mRNA Expression | Confirmed Protein Expression | |||||||
|---|---|---|---|---|---|---|---|---|
| NR4A1, NR4A2, NR4A3, CD83 CD39, CD73, TGF-β, IL-10, PD-L1, IL-10R, IL-27β, IL-12 p35, HLA-G | NR4A1, NR4A3, CD83, CD39, CD73, PD-L1, IL-10 | |||||||
| [ | ||||||||
| 64 | ||||||||
| ] | ||||||||
| [ | ||||||||
| 65 | ||||||||
| ] | ||||||||
| GIFT-15 B-cells | ||||||||
| B220 + CD21 + CD22 + CD23 + CD24 + CD1d + CD138 + IgM + IgD+ | IL-10 | [ | 66 | ] | ||||
| Human | MZp | CD19 + CD1c + CD21 | lo | IgM | hi | CD27 + CD10+ | CD83, PD-L1, IL-10 | [5][67] |
| Transitional B-cells | CD19 + CD24 | hi | CD38 | hi | IL-10 | [68] | ||
| Memory B-cells | CD19 + CD24 |
4.1. Importance of NR4As
4.2. Importance of CD39 and CD73
References
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: Virtues of innate-like antibody-producing lymphocytes. Nat. Rev. Immunol. 2013, 13, 118–132.
- Palm, E.A.-K.; Kleinau, S. Marginal zone B cells: From housekeeping function to autoimmunity? J. Autoimmun. 2021, 119, 102627.
- Weill, J.-C.; Weller, S.; Reynaud, C.-A. Human Marginal Zone B Cells. Annu. Rev. Immunol. 2009, 27, 267–285.
- Fontaine, J.; Chagnon-Choquet, J.; Valcke, H.S.; Poudrier, J.; Roger, M.; the Montreal Primary HIV Infection and Long-Term Non-Progressor Study Groups. High expression levels of B lymphocyte stimulator (BLyS) by dendritic cells correlate with HIV-related B-cell disease progression in humans. Blood 2011, 117, 145–155.
- Chagnon-Choquet, J.; Fontaine, J.; Poudrier, J.; Roger, M.; for the Montreal Primary HIV Infection and Slow Progressor Study Groups. IL-10 and Lymphotoxin-α Expression Profiles within Marginal Zone-Like B-Cell Populations Are Associated with Control of HIV-1 Disease Progression. PLOS ONE 2014, 9, e101949.
- Martin, F.; Kearney, J.F. Marginal-zone B cells. Nat. Rev. Immunol. 2002, 2, 323–335.
- Jackson, R.T.; Ling, R.E.; Roy, A. The Origin of B-cells: Human Fetal B Cell Development and Implications for the Patho-genesis of Childhood Acute Lymphoblastic Leukemia. Front. Immunol. 2021, 12.
- Hardy, R.R.; Hayakawa, K. B CELL DEVELOPMENT PATHWAYS. Annu. Rev. Immunol. 2001, 19, 595–621.
- Nishana, M.; Raghavan, S.C. Role of recombination activating genes in the generation of antigen receptor diversity and beyond. Immunology 2012, 137, 271–281.
- Pieper, K.; Grimbacher, B.; Eibel, H. B-cell biology and development. J. Allergy Clin. Immunol. 2013, 131, 959–971.
- Zhou, Y.; Zhang, Y.; Han, J.; Yang, M.; Zhu, J.; Jin, T. Transitional B cells involved in autoimmunity and their impact on neuroimmunological diseases. J. Transl. Med. 2020, 18, 131.
- Pillai, S.; Cariappa, A. The follicular versus marginal zone B lymphocyte cell fate decision. Nat. Rev. Immunol. 2009, 9, 767–777.
- Tull, T.J.; Pitcher, M.J.; Guesdon, W.; Siu, J.H.; Lebrero-Fernández, C.; Zhao, Y.; Petrov, N.; Heck, S.; Ellis, R.; Dhami, P.; et al. Human marginal zone B cell development from early T2 progenitors. J. Exp. Med. 2021, 218, e20202001.
- Tiegs, S.L.; Russell, D.M.; Nemazee, D. Receptor editing in self-reactive bone marrow B cells. The Journal of Experimental Medicine. 1993. 177: 1009-1020. J. Immunol. 2011, 186, 1313–1324.
- Spencer, J.; Finn, T.; Pulford, K.A.; Mason, D.Y.; Isaacson, P.G. The human gut contains a novel population of B lymphocytes which resemble marginal zone cells. Clin. Exp. Immunol. 1985, 62, 607–612.
- Thomas, L.; Rothstein, D.O.; Griffin, N.E.; Holodick, T.D. Human B-1 cells take the stage. Ann. N. Y. Acad. Sci. 2013, 1285, 97–114.
- Baumgarth, N. A Hard(y) Look at B-1 Cell Development and Function. J. Immunol. 2017, 199, 3387–3394.
- Lechner, M.; Engleitner, T.; Babushku, T.; Schmidt-Supprian, M.; Rad, R.; Strobl, L.J.; Zimber-Strobl, U. Notch2-mediated plasticity between marginal zone and follicular B cells. Nat. Commun. 2021, 12, 1111.
- Cariappa, A.; Tang, M.; Parng, C.; Nebelitskiy, E.; Carroll, M.; Georgopoulos, K.; Pillai, S. The Follicular versus Marginal Zone B Lymphocyte Cell Fate Decision Is Regulated by Aiolos, Btk, and CD21. Immunity 2001, 14, 603–615.
- Hammad, H.; Vanderkerken, M.; Pouliot, P.; Deswarte, K.; Toussaint, W.; Vergote, K.; Vandersarren, L.; Janssens, S.; Ramou, I.; Savvides, S.N.; et al. Transitional B cells commit to marginal zone B cell fate by Taok3-mediated surface expression of ADAM10. Nat. Immunol. 2017, 18, 313–320.
- Moran, S.T.; Cariappa, A.; Liu, H.; Muir, B.; Sgroi, D.; Boboila, C.; Pillai, S. Synergism between NF-κB1/p50 and Notch2 during the Development of Marginal Zone B Lymphocytes. J. Immunol. 2007, 179, 195–200.
- Jia, W.; Poe, J.C.; Su, H.; Anand, S.; Matsushima, G.K.; Rathmell, J.C.; Maillard, I.; Radojcic, V.; Imai, K.; Reyes, N.J.; et al. BAFF promotes heightened BCR responsiveness and manifestations of chronic GVHD after allogeneic stem cell transplantation. Blood 2021, 137, 2544–2557.
- Dono, M.; Zupo, S.; Leanza, N.; Melioli, G.; Fogli, M.; Melagrana, A.; Chiorazzi, N.; Ferrarini, M. Heterogeneity of Tonsillar Subepithelial B Lymphocytes, the Splenic Marginal Zone Equivalents. J. Immunol. 2000, 164, 5596–5604.
- Weller, S. Human blood IgM memory B cells are circulating splenic marginal zone B cells harboring a prediversified immunoglobulin repertoire. Blood 2004, 104, 3647–3654.
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616.
- Hendricks, J.; Bos, N.A.; Kroese, F.G. Heterogeneity of Memory Marginal Zone B Cells. Crit. Rev. Immunol. 2018, 38, 145–158.
- Berkowska, M.A.; Driessen, G.J.A.; Bikos, V.; Grosserichter-Wagener, C.; Stamatopoulos, K.; Cerutti, A.; He, B.; Biermann, K.; Lange, J.F.; van der Burg, M.; et al. Human memory B cells originate from three distinct germinal center-dependent and -independent maturation pathways. Blood 2011, 118, 2150–2158.
- Seifert, M.; Küppers, R. Human memory B cells. Leukemia 2016, 30, 2283–2292.
- Janeway, C.A.Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216.
- Bendelac, A.; Bonneville, M.; Kearney, J.F. Autoreactivity by design: Innate B and T lymphocytes. Nat. Rev. Immunol. 2001, 1, 177–186.
- Guinamard, R.; Okigaki, M.; Schlessinger, J.; Ravetch, J.V. Absence of marginal zone B cells in Pyk-2–deficient mice defines their role in the humoral response. Nat. Immunol. 2000, 1, 31–36.
- Martin, F.; Oliver, A.M.; Kearney, J.F. Marginal Zone and B1 B Cells Unite in the Early Response against T-Independent Blood-Borne Particulate Antigens. Immunity 2001, 14, 617–629.
- Song, H.; Cerny, J. Functional heterogeneity of marginal zone B cells revealed by their ability to generate both early anti-body-forming cells and germinal centers with hypermutation and memory in response to a T-dependent antigen. J. Exp. Med. 2003, 198, 1923–1935.
- Chappell, C.P.; Draves, K.E.; Giltiay, N.V.; Clark, E.A. Extrafollicular B cell activation by marginal zone dendritic cells drives T cell-dependent antibody re-sponses. J. Exp. Med. 2012, 209, 1825–1840.
- Cinamon, G.; Zachariah, M.A.; Lam, O.M.; Foss, F.W.; Cyster, J.G. Follicular shuttling of marginal zone B cells facilitates antigen transport. Nat. Immunol. 2008, 9, 54–62.
- Biram, A.; Davidzohn, N.; Shulman, Z. T cell interactions with B cells during germinal center formation, a three-step model. Immunol. Rev. 2019, 288, 37–48.
- He, B.; Qiao, X.; Klasse, P.J.; Chiu, A.; Chadburn, A.; Knowles, D.M.; Moore, J.P.; Cerutti, A. HIV-1 Envelope Triggers Polyclonal Ig Class Switch Recombination through a CD40-Independent Mechanism Involving BAFF and C-Type Lectin Receptors. J. Immunol. 2006, 176, 3931–3941.
- Roco, J.A.; Mesin, L.; Binder, S.C.; Nefzger, C.; Gonzalez-Figueroa, P.; Canete, P.F.; Ellyard, J.; Shen, Q.; Robert, P.A.; Cappello, J.; et al. Class-Switch Recombination Occurs Infrequently in Germinal Centers. Immunity 2019, 51, 337–350.e7.
- Gatto, D.; Brink, R. The germinal center reaction. J. Allergy Clin. Immunol. 2010, 126, 898–907.
- Weller, S.; Mamani-Matsuda, M.; Picard, C.; Cordier, C.; Lecoeuche, D.; Gauthier, F.; Weill, J.C.; Reynaud, C.A. Somatic diversification in the absence of antigen-driven responses is the hallmark of the IgM+ IgD+ CD27+ B cell repertoire in infants. J. Exp. Med. 2008, 205, 1331–1342.
- Scheeren, F.A. T cell-independent development and induction of somatic hypermutation in human IgM+ IgD+ CD27+ B cells. J. Exp. Med. 2008, 205, 2033–2042.
- Puga, I.; Cols, M.; Barra, C.M.; He, B.; Cassis, L.; Gentile, M.; Comerma, L.; Chorny, A.; Shan, M.; Xu, W.; et al. B cell–helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nat. Immunol. 2011, 13, 170–180.
- He, B.; Santamaria, R.; Xu, W.; Cols, M.; Chen, K.; Puga, I.; Shan, M.; Xiong, H.; Bussel, J.B.; Chiu, A.; et al. The transmembrane activator TACI triggers immunoglobulin class switching by activating B cells through the adaptor MyD88. Nat. Immunol. 2010, 11, 836–845.
- Treml, L.S.; Carlesso, G.; Hoek, K.L.; Stadanlick, J.E.; Kambayashi, T.; Bram, R.J.; Cancro, M.P.; Khan, W.N. TLR stimulation modifies BLyS receptor expression in follicular and marginal zone B cells. J. Immunol. 2007, 178, 7531–7539.
- Attanavanich, K.; Kearney, J.F. Marginal Zone, but Not Follicular B Cells, Are Potent Activators of Naive CD4 T Cells. J. Immunol. 2004, 172, 803–811.
- Marinkovic, D.; Marinkovic, T. Putative role of marginal zone B cells in pathophysiological processes. Scand. J. Immunol. 2020, 92, e12920.
- Bialecki, E.; Paget, C.; Fontaine, J.; Capron, M.; Trottein, F.; Faveeuw, C. Role of Marginal Zone B Lymphocytes in Invariant NKT Cell Activation. J. Immunol. 2009, 182, 6105–6113.
- Mauri, C.; Menon, M. The expanding family of regulatory B cells. Int. Immunol. 2015, 27, 479–486.
- Mauri, C.; Bosma, A. Immune Regulatory Function of B Cells. Annu. Rev. Immunol. 2012, 30, 221–241.
- Iizuka-Koga, M.; Nakatsukasa, H.; Ito, M.; Akanuma, T.; Lu, Q.; Yoshimura, A. Induction and maintenance of regulatory T cells by transcription factors and epigenetic modifications. J. Autoimmun. 2017, 83, 113–121.
- Yanaba, K.; Bouaziz, J.-D.; Matsushita, T.; Tsubata, T.; Tedder, T.F. The Development and Function of Regulatory B Cells Expressing IL-10 (B10 Cells) Requires Antigen Receptor Diversity and TLR Signals. J. Immunol. 2009, 182, 7459–7472.
- Yanaba, K.; Bouaziz, J.-D.; Haas, K.M.; Poe, J.C.; Fujimoto, M.; Tedder, T.F. A Regulatory B Cell Subset with a Unique CD1dhiCD5+ Phenotype Controls T Cell-Dependent Inflammatory Responses. Immunity 2008, 28, 639–650.
- Huber, K.; Sármay, G.; Kövesdi, D. MZ B cells migrate in a T-bet dependent manner and might contribute to the remission of collagen-induced arthritis by the secretion of IL-10. Eur. J. Immunol. 2016, 46, 2239–2246.
- Evans, J.G.; Chavez-Rueda, K.A.; Eddaoudi, A.; Meyer-Bahlburg, A.; Rawlings, D.J.; Ehrenstein, M.; Mauri, C. Novel Suppressive Function of Transitional 2 B Cells in Experimental Arthritis. J. Immunol. 2007, 178, 7868–7878.
- Hsu, L.-H.; Li, K.-P.; Chu, K.-H.; Chiang, B.-L. A B-1a cell subset induces Foxp3− T cells with regulatory activity through an IL-10-independent pathway. Cell Mol. Immunol. 2015, 12, 354–365.
- Fillatreau, S. Natural regulatory plasma cells. Curr. Opin. Immunol. 2018, 55, 62–66.
- Matsumoto, M.; Baba, A.; Yokota, T.; Nishikawa, H.; Ohkawa, Y.; Kayama, H.; Kallies, A.; Nutt, S.L.; Sakaguchi, S.; Takeda, K.; et al. Interleukin-10-Producing Plasmablasts Exert Regulatory Function in Autoimmune Inflammation. Immunity 2014, 41, 1040–1051.
- Ding, Q.; Yeung, M.; Camirand, G.; Zeng, Q.; Akiba, H.; Yagita, H.; Chalasani, G.; Sayegh, M.H.; Najafian, N.; Rothstein, D.M. Regulatory B cells are identified by expression of TIM-1 and can be induced through TIM-1 ligation to promote tolerance in mice. J. Clin. Investig. 2011, 121, 3645–3656.
- Zhang, Y.; Li, J.; Zhou, N.; Zhang, Y.; Wu, M.; Xu, J.; Shen, C.; An, X.; Shen, G.; Yang, M.; et al. The Unknown Aspect of BAFF: Inducing IL-35 Production by a CD5(+)CD1d(hi)FcγRIIb(hi) Regulatory B-Cell Subset in Lupus. J. Invest. Derm. 2017, 137, 2532–2543.
- Ray, A.; Basu, S.; Williams, C.B.; Salzman, N.; Dittel, B.N. A Novel IL-10–Independent Regulatory Role for B Cells in Suppressing Autoimmunity by Maintenance of Regulatory T Cells via GITR Ligand. J. Immunol. 2012, 188, 3188–3198.
- Lundy, S.K. Killer B lymphocytes: The evidence and the potential. Agents Actions 2009, 58, 345–357.
- Tian, J.; Zekzer, D.; Hanssen, L.; Lu, Y.; Olcott, A.; Kaufman, D.L. Lipopolysaccharide-Activated B Cells Down-Regulate Th1 Immunity and Prevent Autoimmune Diabetes in Nonobese Diabetic Mice. J. Immunol. 2001, 167, 1081–1089.
- Khan, A.R.; Hams, E.; Floudas, A.; Sparwasser, T.; Weaver, C.T.; Fallon, P. PD-L1hi B cells are critical regulators of humoral immunity. Nat. Commun. 2015, 6, 5997.
- LeBleu, V.S.; Kalluri, R. Exosomes Exercise Inhibition of Anti-Tumor Immunity during Chemotherapy. Immunity 2019, 50, 547–549.
- Kaku, H. A novel mechanism of B cell-mediated immune suppression through CD73 expression and adenosine pro-duction. J. Immunol. 2014, 193, 5904–5913.
- Rafei, M.; Hsieh, J.; Zehntner, S.; Li, M.; Forner, K.; Birman, E.; Boivin, M.-N.; Young, Y.K.; Perreault, C.; Galipeau, J. A granulocyte-macrophage colony–stimulating factor and interleukin-15 fusokine induces a regulatory B cell population with immune suppressive properties. Nat. Med. 2009, 15, 1038–1045.
- Doyon-Laliberté, K.; Chagnon-Choquet, J.; Byrns, M.; Aranguren, M.; Memmi, M.; Chrobak, P.; Stagg, J.; Poudrier, J.; Roger, M. NR4A Expression by Human Marginal Zone B-Cells. Antibodies 2019, 8, 50.
- Blair, P.A.; Yassin-Noreña, L.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.; Ehrenstein, M.; Mauri, C. CD19+CD24hiCD38hi B Cells Exhibit Regulatory Capacity in Healthy Individuals but Are Functionally Impaired in Systemic Lupus Erythematosus Patients. Immunity 2010, 32, 129–140.
- Jin, L.; Weiqian, C.; Lihuan, Y. Peripheral CD24 hi CD27 + CD19 + B cells subset as a potential biomarker in naïve systemic lupus erythematosus. Int. J. Rheum. Dis. 2013, 16, 698–708.
- van de Veen, W.; Stanic, B.; Yaman, G.; Wawrzyniak, M.; Söllner, S.; Akdis, D.G. IgG4 production is confined to human IL-10-producing regulatory B cells that suppress antigen-specific immune responses. J. Allergy Clin. Immunol. 2013, 131, 1204–1212.
- Gu, X.L.; He, H.; Lin, L.; Luo, G.X.; Wen, Y.F.; Xiang, D.C.; Qiu, J. Tim-1(+) B cells suppress T cell interferon-gamma production and promote Foxp3 expression, but have impaired regulatory function in coronary artery disease. Apmis 2017, 125, 872–879.
- Mao, H.; Pan, F.; Wu, Z.; Wang, Z.; Zhou, Y.; Zhang, P.; Gou, M.; Dai, G. CD19loCD27hi Plasmablasts Suppress Harmful Th17 Inflammation Through Interleukin 10 Pathway in Colorectal Cancer. DNA Cell Biol. 2017, 36, 870–877.
- Rybojad, M.; Hau, E.; Monfort, B.J.; Branchtein, M.; Michonneau, D.; Dessirier, V.; Socié, G. CD24(hi)CD27+ and plasmablast-like regulatory B cells in human chronic graft-versus-host disease. Blood 2015, 125, 1830–1839.
- Fehres, C.M.; Van Uden, N.O.; Yeremenko, N.G.; Fernandez, L.; Salinas, G.F.; Van Duivenvoorde, L.M.; Huard, B.; Morel, J.; Spits, H.; Hahne, M.; et al. APRIL Induces a Novel Subset of IgA+ Regulatory B Cells That Suppress Inflammation via Expression of IL-10 and PD-L1. Front. Immunol. 2019, 10.
- Wang, X.; Zhu, Y.; Zhang, M.; Wang, H.; Jiang, Y.; Gao, P. Ulcerative Colitis Is Characterized by a Decrease in Regulatory B Cells. J. Crohn's Colitis 2016, 10, 1212–1223.
- van Rensburg, I.C.; Loxton, A.G. B-cells with a FasL expressing regulatory phenotype are induced following successful anti-tuberculosis treatment. Tuberculosis 2018, 108, 114–117.
- Figueiro, F.; Muller, L.; Funk, S.; Jackson, E.K.; Battastini, A.M.O.; Whiteside, T.L. Phenotypic and functional characteristics of CD39high human regulatory B cells (Breg). OncoImmunology 2016, 5, e1082703.
- Nouël, A.; Pochard, P.; Simon, Q.; Ségalen, I.; Le Meur, Y.; Pers, J.; Hillion, S. B-Cells induce regulatory T cells through TGF-β/IDO production in A CTLA-4 dependent manner. J. Autoimmun. 2015, 59, 53–60.
- Sabourin-Poirier, C.; Fourcade, L.; Chagnon-Choquet, J.; Labbé, A.C.; Alary, M.; Guédou, F.; Roger, M. Blood B Lymphocyte Stimulator (BLyS)/BAFF levels may reflect natural immunity to HIV in highly exposed uninfected Beninese Commercial Sex Workers. Sci. Rep. 2016, 6, 32318.
- Safe, S.; Jin, U.-H.; Morpurgo, B.; Abudayyeh, A.; Singh, M.; Tjalkens, R.B. Nuclear receptor 4A (NR4A) family—orphans no more. J. Steroid Biochem. Mol. Biol. 2015, 157, 48–60.
- Sekiya, T.; Hibino, S.; Saeki, K.; Kanamori, M.; Takaki, S.; Yoshimura, A. Nr4a Receptors Regulate Development and Death of Labile Treg Precursors to Prevent Generation of Path-ogenic Self-Reactive Cells. Cell Rep. 2018, 24, 1627–1638.e6.
- Bandukwala, H.S.; Rao, A. Nurrishing Treg cells: Nr4a transcription factors control Foxp3 expression. Nat. Immunol. 2013, 14, 201–203.
- Ashouri, J.; Weiss, A. Endogenous Nur77 Is a Specific Indicator of Antigen Receptor Signaling in Human T and B Cells. J. Immunol. 2016, 198, 657–668.
- Crean, D.; Cummins, E.P.; Bahar, B.; Mohan, H.; McMorrow, J.P.; Murphy, E.P. Adenosine Modulates NR4A Orphan Nuclear Receptors To Attenuate Hyperinflammatory Responses in Monocytic Cells. J. Immunol. 2015, 195, 1436–1448.
- Wan, P.K.-T.; Siu, M.K.-Y.; Leung, T.H.-Y.; Mo, X.-T.; Chan, K.K.-L.; Ngan, H.Y.-S. Role of Nurr1 in Carcinogenesis and Tumor Immunology: A State of the Art Review. Cancers 2020, 12, 3044.
- Hibino, S.; Chikuma, S.; Kondo, T.; Ito, M.; Nakatsukasa, H.; Omata-Mise, S.; Yoshimura, A. Inhibition of Nr4a Receptors Enhances Antitumor Immunity by Breaking Treg-Mediated Immune Tolerance. Cancer Res. 2018, 78, 3027–3040.
- Boulet, S.; Daudelin, J.-F.; Odagiu, L.; Pelletier, A.-N.; Yun, T.J.; Lesage, S.; Cheong, C.; Labrecque, N. The orphan nuclear receptor NR4A3 controls the differentiation of monocyte-derived dendritic cells following microbial stimulation. Proc. Natl. Acad. Sci. USA 2019, 116, 15150–15159.
- Mildner, A.; Schönheit, J.; Giladi, A.; David, E.; Lara-Astiaso, D.; Vivas, E.L.; Paul, F.; Chappell-Maor, L.; Priller, J.; Leutz, A.; et al. Genomic Characterization of Murine Monocytes Reveals C/EBPβ Transcription Factor Dependence of Ly6C—Cells. Immunity 2017, 46, 849–862.e7.
- Hanna, R.N.; Carlin, L.M.; Hubbeling, H.G.; Nackiewicz, D.; Green, A.M.; Punt, J.A.; Geissmann, F.; Hedrick, C.C. The transcription factor NR4A1 (Nur77) controls bone marrow differentiation and the survival of Ly6C− monocytes. Nat. Immunol. 2011, 12, 778–785.
- Tel-Karthaus, N.; Kers-Rebel, E.D.; Looman, M.W.; Ichinose, H.; De Vries, C.J.; Ansems, M. Nuclear Receptor Nur77 Deficiency Alters Dendritic Cell Function. Front. Immunol. 2018, 9.
- Karki, K.; Wright, G.A.; Mohankumar, K.; Jin, U.-H.; Zhang, X.-H.; Safe, S. A Bis-Indole–Derived NR4A1 Antagonist Induces PD-L1 Degradation and Enhances Antitumor Immunity. Cancer Res. 2020, 80, 1011–1023.
- Bridi, M.S.; Hawk, J.D.; Chatterjee, S.; Safe, S.; Abel, T. Pharmacological Activators of the NR4A Nuclear Receptors Enhance LTP in a CREB/CBP-Dependent Manner. Neuropsychopharmacology 2016, 42, 1243–1253.
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The Role of the Transcription Factor CREB in Immune Function. J. Immunol. 2010, 185, 6413–6419.
- Duren, R.P.; Boudreaux, S.P.; Conneely, O.M. Genome Wide Mapping of NR4A Binding Reveals Cooperativity with ETS Factors to Promote Epigenetic Activation of Distal Enhancers in Acute Myeloid Leukemia Cells. PLoS ONE 2016, 11, e0150450.
- Ge, W.; Arp, J.; Lian, D.; Liu, W.; Baroja, M.L.; Jiang, J.; Ramcharran, S.; Eldeen, F.Z.; Zinser, E.; Steinkasserer, A.; et al. Immunosuppression Involving Soluble CD83 Induces Tolerogenic Dendritic Cells That Prevent Cardiac Allograft Rejection. Transplantation 2010, 90, 1145–1156.
- Lan, Z.; Ge, W.; Arp, J.; Jiang, J.; Liu, W.; Gordon, D.; Healey, D.; Debenedette, M.; Nicolette, C.; Garcia, B.; et al. Induction of Kidney Allograft Tolerance by Soluble CD83 Associated With Prevalence of Tolerogenic Dendritic Cells and Indoleamine 2,3-Dioxygenase. Transplantation 2010, 90, 1286–1293.
- Lin, H.; Liang, S.; Zhong, Z.; Wen, J.; Li, W.; Wang, L.; Xu, J.; Zhong, F.; Li, X. Soluble CD83 inhibits human monocyte differentiation into dendritic cells in vitro. Cell. Immunol. 2014, 292, 25–31.
- Bates, J.M.; Flanagan, K.; Mo, L.; Ota, N.; Ding, J.; Ho, S.; Liu, S.; Roose-Girma, M.; Warming, S.; Diehl, L. Dendritic cell CD83 homotypic interactions regulate inflammation and promote mucosal homeostasis. Mucosal Immunol. 2014, 8, 414–428.
- Li, Z.; Ju, X.; Silveira, P.A.; Abadir, E.; Hsu, W.-H.; Hart, D.N.J.; Clark, G. CD83: Activation Marker for Antigen Presenting Cells and Its Therapeutic Potential. Front. Immunol. 2019, 10, 1312.
- Wild, A.B.; Krzyzak, L.; Peckert, K.; Stich, L.; Kuhnt, C.; Butterhof, A.; Seitz, C.; Mattner, J.; Grüner, N.; Gänsbauer, M.; et al. CD83 orchestrates immunity toward self and non-self in dendritic cells. JCI Insight 2019, 4, e126246.
- Sénéchal, B.; Boruchov, A.M.; Reagan, J.L.; Hart, D.N.J.; Young, J.W. Infection of mature monocyte-derived dendritic cells with human cytomegalovirus inhibits stimulation of T-cell proliferation via the release of soluble CD83. Blood 2004, 103, 4207–4215.
- Bock, F.; Rössner, S.; Onderka, J.; Lechmann, M.; Pallotta, M.T.; Fallarino, F.; Boon, L.; Nicolette, C.; Debenedette, M.A.; Tcherepanova, I.Y.; et al. Topical Application of Soluble CD83 Induces IDO-Mediated Immune Modulation, Increases Foxp3+ T Cells, and Prolongs Allogeneic Corneal Graft Survival. J. Immunol. 2013, 191, 1965–1975.
- Doebbeler, M.; Koenig, C.; Krzyzak, L.; Seitz, C.; Wild, A.; Ulas, T.; Baßler, K.; Kopelyanskiy, D.; Butterhof, A.; Kuhnt, C.; et al. CD83 expression is essential for Treg cell differentiation and stability. JCI Insight 2018, 3, e99712.
- Saze, Z.; Schuler, P.J.; Hong, C.-S.; Cheng, D.; Jackson, E.K.; Whiteside, T.L. Adenosine production by human B cells and B cell–mediated suppression of activated T cells. Blood 2013, 122, 9–18.
- Antonioli, L.; Pacher, P.; Vizi, E.S.; Haskó, G. CD39 and CD73 in immunity and inflammation. Trends Mol. Med. 2013, 19, 355–367.
- Minguet, S.; Huber, M.; Rosenkranz, L.; Schamel, W.W.; Reth, M.; Brummer, T. Adenosine and cAMP are potent inhibitors of the NF-kappa B pathway downstream of immunoreceptors. Eur. J. Immunol. 2005, 35, 31–41.
- Dwyer, K.M.; Deaglio, S.; Gao, W.; Friedman, D.; Strom, T.B.; Robson, S.C. CD39 and control of cellular immune responses. Purinergic Signal. 2007, 3, 171–180.
- Allard, B.; Allard, D.; Buisseret, L.; Stagg, J. The adenosine pathway in immuno-oncology. Nat. Rev. Clin. Oncol. 2020, 17, 611–629.