Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Lisa Mapelli and Version 3 by Amina Yu.
Autism spectrum disorders (ASD) are pervasive neurodevelopmental disorders that include a variety of forms and clinical phenotypes. This heterogeneity complicates the clinical and experimental approaches to ASD etiology and pathophysiology. To date, a unifying theory of these diseases is still missing. With the recent acknowledgement of the cerebellar contribution to cognitive functions and the social brain, its involvement in ASD has become unmistakable, though its extent is still to be elucidated.
- cerebellum
- autism spectrum disorders
- social brain
- excitatory/inhibitory balance
1. Neural Bases for Impaired Social Cognition in Autism Spectrum Disorders (ASD)
1. Neural Bases for Impaired Social Cognition in ASD
In humans, focal brain lesions and social task-based fMRI studies have largely contributed to identifying a network of brain regions (called “the social brain”) implicated in social cognition [1][2][3][4][5][33,34,35,36,37]. The primary regions of the social brain include the medial prefrontal cortex (mPFC) [6][7][38,39], the temporoparietal junction (TPJ), the posterior superior temporal sulcus (pSTS), the inferior frontal gyrus, the anterior cingulate cortex (ACC) [8][40], and the anterior insula (AI) [9][41] (Figure 1). Moreover, the hippocampal formation, the ventral tegmental area (VTA), the nucleus accumbens (NAcc), the amygdala, and the cerebellum are highly connected with the social network structure, acting as important functional hubs [10][11][12][13][14][15][16][42,43,44,45,46,47,48]. In ASD subjects, several studies identified a combination of atypical structural and functional features in these areas. Structurally, cortical and subcortical measurements in ASD postmortem brain tissue, primarily in frontal and temporal cortices and the amygdala, described an aberrant organization, such as small cell size and increased packing density [17][18][49,50], white matter volume increase [19][51], decreased cortical thickness [20][52], and more numerous and narrower minicolumns [21][53]. Functionally, a growing number of stusdies used fMRI to examine changes in intrinsic functional connectivity (FC) of specific brain regions and circuits [22][23][54,55] between individuals with ASD and normally developing controls. In most cases, FC analysis indicated that ASD subjects exhibit long-range under-connectivity and local over-connectivity [24][25][26][27][28][29][30][31][56,57,58,59,60,61,62,63]. Long-range under-connectivity between prefrontal cortex (PFC) and posterior brain regions were most often reported [24][25][26][32][33][56,57,58,64,65], but it was also described in other regions, as between the amygdala and temporal cortex [34][66], the supplementary motor areas and the thalamus [35][67], the PFC and premotor and somatosensory cortices [36][68], and among the PFC, amygdala, and hippocampus [37][69]. However, some studies reported increased FC among individuals with ASD [37][69]. Specifically, long-range over-connectivity was discovered within thalamocortical [38][70], striatocortical [39][71], and corticocortical circuits [40][72]. In contrast, local over-connectivity in ASD is less solidly determined. For instance, local over-connectivity was found in ASD in the extrastriate cortex, frontal and temporal regions, amygdala, and parahippocampal gyri [41][42][43][44][45][73,74,75,76,77]. Other studies, however, reported a reduction of local connectivity, principally in the fusiform face area and in the somatosensory cortex [46][47][78,79], or a combination of both patterns [48][80]. Several experimental evidence suggested a higher excitatory/inhibitory (E/I) ratio as a possible correlate for local over-connectivity [49][50][51][81,82,83], for example, through an increased glutamatergic or reduced GABAergic signaling [52][53][84,85]. Cortical GABAergic neurons are thought to control the functional integrity and segregation of minicolumns via lateral inhibition [54][86]. Casanova and colleagues [21][53] found significant differences between frontal and temporal cortices of ASD and typically developing individuals in the number of minicolumns, in the horizontal space between minicolumns, and their internal structure. Ultimately, minicolumns were more numerous, smaller, and less compact in their cellular configuration. Mechanisms underlying this deficit are still unknown. Moreover, GABAergic neurotransmission is involved in generating gamma-band oscillatory activity [55][87]. Gamma-band oscillations are involved in a wide range of cognitive processes from the perception of gestalt [56][88] to selective attention [57][58][59][89,90,91] and working memory [60][61][92,93]. Magnetoencephalography (MEG) and electroencephalography (EEG) studies have reported correlations between gamma-band oscillatory activity and ASD severity as measured by the Social Responsiveness Scale [62][63][64][94,95,96]. It should also be considered that an unbalanced E/I ratio might be amplified by delayed brain development, resulting in retardation of synaptogenesis, pruning, and myelination [65][66][67][97,98,99]. Lastly, strong evidence is also reported for alterations in glutamatergic signaling pathways in ASD, involving metabotropic glutamate receptor 5 (mGluR5) upregulation and genetic aberrations associated with NMDA receptors [68][69][100,101]. However, the scenario is much more complicated, with both increases and decreases in glutamate-mediated signaling reported in association with the ASD phenotype [70][102]. Overall, the above-presented data lend support to models hypothesizing well-defined neural substrates of social cognition and propose specific neural bases that may govern social cognitive impairments in ASD. By contrast, further investigations are needed to better understand the complex interactions between social brain areas, connectivity, frequency bands, and physiological aspects (for i.example., roles of specific cell types, maturational processes, receptors) and how they relate to different cognitive processes.
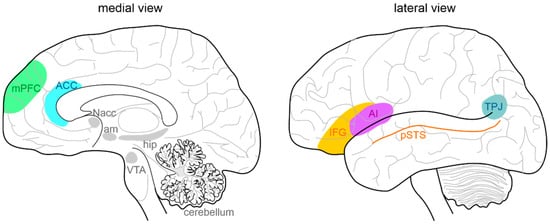
Figure 1. Anatomy of the social brain. The main brain areas involved in the “social brain” are reported in the medial (left) and lateral (right) schematic view of the human brain: medial prefrontal cortex (mPFC), anterior cingulate cortex (ACC), temporo-parietal junction (TPJ), posterior superior temporal sulcus (pSTS), inferior frontal gyrus (IFG), and anterior insula (AI). The main regions connected to the “social brain” are reported in grey: hippocampus (hip), amygdala (am), ventral tegmental area (VTA), nucleus accumbens (NAcc), and cerebellum.
2. Cerebellar Involvement in ASD
2.1. Cerebellar Circuit Microanatomy
The cerebellum, meaning “little brain” in Latin, has been historically considered a subcortical motor structure that controls the coordination of voluntary movements, balance, posture, and muscle tone. Furthermore, it contributes to different forms of motor learning. There is now robust evidence that the cerebellum may be related to a variety of cognitive and emotional functions such as language, attention, fear, and pleasure responses [71][72][73][74][30,32,103,104]. The cerebellum is composed of tightly folded layers of grey matter forming the cerebellar cortex, with the white matter underneath surrounding four deep cerebellar nuclei (DCN) [75][105]. The cerebellar cortex is organized into three layers. The outer molecular layer (ML) is composed of two types of inhibitory neurons: stellate (SCs) and basket cells (BCs). The Purkinje cell layer consists of a large pear-shaped Purkinje cell (PC) soma monolayer. The inner granular layer is composed of excitatory granule cells (GrCs) and inhibitory Golgi cells (GoCs). The primary input pathways entering the cerebellum are the mossy fibers (MFs) and climbing fibers (CFs). In the granular layer, MFs directly synapse on the dendrites of GrCs, whose axons ascend toward the ML, where they bifurcate to form T-shaped branches named parallel fibers (PFs) [76][77][78][106,107,108]. PCs receive excitatory input from PFs and CFs, which originate in the inferior olive (IO) [79][109] and project their axons to DCN neurons. DCN neurons provide the final output of the cerebellum by integrating inhibitory and excitatory inputs from PC axons, MF, and CF collaterals, respectively [80][81][110,111]. The activity of PCs is modulated by three types of inhibitory interneurons that are activated by PFs and classified into two main types: BCs and SCs, which are found in the ML, and GoCs, located in the granular layer. Specifically, BCs are found in the deep ML and their axons form pericellular nests in close proximity to PC soma as well as specialized terminals known as pinceaux surrounding the initial segment of PC axons. SCs are located in the upper ML and their axons terminate on the shafts of PC dendrites [82][83][112,113]. GoCs receive excitatory synaptic input from MFs on the basal dendrites and PFs on the apical dendrites [84][85][114,115], and their axons make inhibitory synapses with GrCs [86][87][116,117]. Thus, GoC activity indirectly affects PC output by modulating GrC discharge [82][88][112,118] (Figure 2).
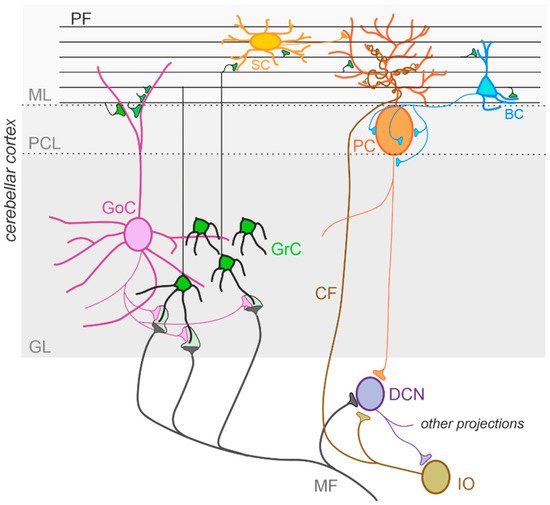
Figure 2. The cerebellar circuit. Schematic representation of the main components of the cerebellar circuit. The inputs are provided by mossy fibers (MF) and climbing fibers (CF), these latter originating in the inferior olive (IO). Both inputs send collaterals to the deep cerebellar nuclei (DCN) before entering the cerebellar cortex. Granule cells (GrC) and Golgi cells (GoC) are the main neuronal types present in the granular layer (GL) of the cerebellar cortex. GrC axons reach the molecular layer (ML) where they bifurcate originating the parallel fibers (PF). The inhibitory interneurons in the ML are stellate cells (SC) and basket cells (BC), which inhibit Purkinje cells (PC) in the Purkinje cell layer (PCL). The PC provides the output of the cerebellar cortex, inhibiting DCN neurons, which in turn provide the main output of the cerebellar circuit. Notice that DCN project to the IO, generating a loop mediated by the CF.
2.2. Cerebellar Connectivity to Social Brain Areas
Experiments using task-based fMRI and positron emission tomography (PET) revealed that separate regions of the cerebellum are associated with distinct cerebral areas through polysynaptic circuits, forming a functional topography [89][90][91][92][119,120,121,122]. The sensorimotor cerebellum is represented in the anterior lobe (lobules I-V) and lobule VIII, while the cognitive cerebellum comprises the posterior lobe (lobules VI and VII), including hemispheric extensions (CrusI/CrusII) [92][122]. Finally, the posterior vermis and hemispheres represent the limbic cerebellum [92][93][94][122,123,124]. The DCN send direct projections to the ventrolateral (VL) and the intralaminar thalamic nuclei, particularly the dorsomedial (MD) nucleus [95][125]. The VL nucleus, classically known as an integrative center for sensorimotor transformations, targets the primary motor cortex (M1) [10][96][97][42,126,127], whereas the MD nucleus, like other intralaminar nuclei, has widespread cortical projections including the medial prefrontal cortex (mPFC) (Figure 3A) and the superior temporal sulcus [98][99][100][101][102][128,129,130,131,132]. Past anatomical studies demonstrated that the cerebellum is interconnected with parts of the limbic system, including the hippocampus, amygdala, and cingulate cortex (Figure 3B) [103][104][133,134]. Recently, Bohne and colleagues [105][135] reported an elegant tracing onestudy identifying a new cerebellar-hippocampal connection via the VL thalamic nucleus in mice. In support of this finding, unilateral removal of the cerebellar hemispheres [106][136] or PC signaling deficits [107][137] determined an impairment in hippocampal-based behavioral tasks as goal-directed navigation tests. Furthermore, monosynaptic projections originate in the hippocampus to primarily target the PFC in rodents and primates [108][109][138,139]. Several shreds of evidence obtained using different methodologies show that the cerebellum and amygdala are connected [110][140]. For example, Sang and colleagues found functional connectivity between cerebellar lobules I-V and the amygdala, analyzing resting-state fMRI in healthy young adults [111][141]. Heath and Harper, recording evoked potentials or using histological tract-tracing, showed connections between DCN and amygdala in cats and monkeys [112][142]. Finally, Morris and colleagues found amygdala and cerebellum coactivation during the presentation of facial expressions in human subjects [113][143]. The cerebellum is also connected with the cingulate cortex indicating its involvement in motivational and emotional processing [114][144]. Early animal onstudies showed electrophysiological responses in the ACC following electrical stimulation of the vermis area [112][115][142,145]. These results were confirmed almost forty years later by Krienen and Buckner using resting-state fMRI in young adults, showing that CrusI and anterior cingulate cortex were functionally connected [116][146]. Lastly, electrical stimulation of DCN was reported to evoke dopamine release in the mPFC in rodents [117][118][119][147,148,149]. Cerebellar modulation of dopamine release onto the mPFC could be mediated by two separate neuronal pathways originating from the DCN. The first one activates the mesocortical dopaminergic pathway via reticulo-tegmental nuclei (RTN), which, in turn, project to pedunculopontine nuclei (PPT) and then directly stimulate VTA dopaminergic cells that send their axons to the mPFC [103][120][121][122][123][133,150,151,152,153]. The second is by modulation of mesocortical dopaminergic release via glutamatergic afferents originating in the thalamic nuclei (VL and MD) [11][124][125][43,154,155] (Figure 3B). More intriguingly, Ra recentl study has shown, using optogenetic manipulation, the existence of a direct cerebellum-VTA pathway suggesting a prominent role of the cerebellum in modulating social behavior [126][156]. The primary cerebellar connections reported above are summarized in Figure 3. Altogether, these findings propose that dysfunctions described within the cerebral cortical network, usually associated with social features of ASD, could be at least partly related to an impaired connectivity between the cerebellum and key social brain areas.
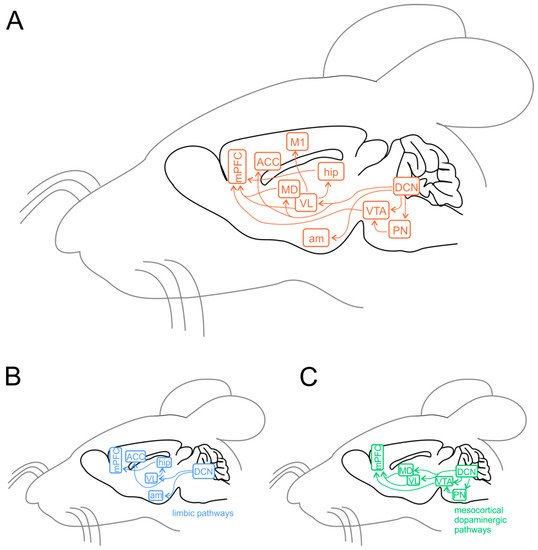
Figure 3. Cerebellar connectivity to other brain areas. The cerebellum is one of the most interconnected structures in the brain. (A) Schematic representation of the mouse brain and the main cerebellar connections thought to be relevant for its role in ASD. DCN, deep cerebellar nuclei; PN, pontine nuclei (including reticulo-tegmental nuclei and pedunculopontine nuclei); VTA, ventral tegmental area; am, amygdala; hip, hippocampus; VL, ventrolateral thalamic nucleus; MD, mediodorsal thalamic nucleus; M1, primary motor cortex; ACC, anterior cingulate cortex; mPFC, medial prefrontal cortex. (B) Same representation as in (A), showing the pathways involving the limbic system. (C) Same representation as in (A), showing the connections involved in the mesocortical dopaminergic pathways, regulating mPFC activity modulation.
2.3. Cerebellar Structural Abnormalities in ASD
The cerebellum is the brain structure most constantly found abnormal in ASD, and an increased risk for ASD is dependently associated with cerebellar damage [127][128][129][130][131][157,158,159,160,161]. Early anatomical onstudies examining postmortem ASD brain tissue reported a significant reduction in the number of PCs in the lateral hemisphere compared with the medial vermis [132][133][162,163]. In subsequent years, the reduction in PC density has been widely documented (about 75% of ASD wcaseres reported in [134][135][136][137]the literature [164,165,166,167]). Fatemi and colleagues [138][168] found a reduction in PC size in about 25% of ASD cases. Additionally, a variable decrease in GrCs numerosity was reported [139][169], while the molecular interneurons were preserved [140][170]. Animal models of spontaneous cerebellar mutations are frequently characterized by PC loss, often showing a failure in the regression of multiple innervations of PCs by CFs, with each PC receiving up to four CF inputs [141][142][143][171,172,173] instead of a single one [144][174]. A similar PC hyper-innervation due to decreased PC number has not yet been described in human ASD brains, but it could provide support to IO neurons, which are unaffected in number [145][146][147][175,176,177]. Concerning the DCN, the neurons presented an enlarged size at a younger age, while older ASD cerebella showed abnormally smaller neurons, also reduced in number in fastigial and interposed nuclei [139][146][169,176]. MRI has emerged as a powerful tool for visualization and diagnostics of neuroanatomical abnormalities in ASD since its inception in the 1980s. Nevertheless, many results are contradictory due to the heterogeneity of underlying causes and the complexity of this disorder. Structural MRI studies in ASD patients described a reduction in the size of several regions of the cerebellum including the vermis, particularly the lobules VI and VII [146][148][149][150][151][152][176,178,179,180,181,182]. In contrast, Piven and colleagues [153][183] did not confirm these findings but revealed an enlarged cerebellar volume in ASD patients. Interestingly, the cerebellar volume was proportionally scaled to the total brain volume [154][184]. Further findings included an enlargement in cerebellar white matter volume and a reduction in the grey/white matter ratio [155][156][185,186]. In conclusion, the abnormalities of cerebellar structural integrity may be considered as significant predictive factors of ASD and cause differences in functional connectivity reported in ASD young adults (see paragraph below).
2.4. Cerebellar Functional Abnormalities in ASD
CStudies in children and young adults with ASD, using resting-state FC (rsFC) techniques, documented a general cerebro-cerebellar over-connectivity [157][187]. However, both under- and over- connectivity have been observed depending on the brain regions investigated. For example, rsFC were increased between non-motor areas of the cerebellum (lobules VI and CrusI) and sensorimotor cerebral cortical regions, such as premotor and primary motor cortices, somatosensory temporal cortex, and occipital lobe; and decreased in cerebro-cerebellar circuits involving language and social interaction, particularly between CrusI/II and PFC, posterior parietal cortex, and the inferior/middle temporal gyrus [127][157][158][157,187,188]. It should be noted that no specific correlation between FC and behavioral profiles in individuals with ASD has been established [42][159][160][161][74,189,190,191], although novel findings reported abnormalities in FC related to ASD symptom severity [162][163][192,193]. To date, very few studies addressed the FC between the cerebellum and cortical regions, focusing on motor task performance in ASD. For example, during self-paced sequential finger tapping, fMRI in children with ASD did not display the activation in the lobules IV/V and in the anterior cerebellum present in typically developing groups [35][67]. Furthermore, Jack and Morris [164][194] investigated coordinated activity between the neocerebellum (particularly CrusI) and pSTS during a task that requires perception and use of information about others’, and remarkably found stronger CrusI–pSTS connectivity positively associated with mentalizing ability, in young adults with ASD. Therefore, together with the structural data described above, these findings are consistent with the idea that ASD is a disorder characterized by abnormalities in cerebellum-cerebral functional connectivity, which could be related to symptom severity.
2.5. Cerebellar Neurochemical Abnormalities in ASD
Neurochemical research has progressed in the last 20 years and has produced promising results. For example, reelin expression was reduced in the cerebellum of ASD individuals [165][195]. This glycoprotein regulates proper cortex lamination and neuronal migration during development and adult life, sustaining cell signaling and synaptic function. Furthermore, serotonin concentration is also altered in the ASD cerebellum. Specifically, Chugani and colleagues [166][167][196,197], using PET scanning with a tracer for serotonin synthesis in ASD young adults, reported reduced serotonin levels in the thalamus and the frontal cortex associated with increased serotonin concentration in DCN. Serotonin is well known for its role in neurodevelopment, regulating cell migration and proliferation [168][198], neurite outgrowth, and neuronal survival [169][199] as well as synaptogenesis [170][200]. Therefore, aberrant serotoninergic neuromodulation of dentatothalamocortical pathway connecting the cerebellum with social structures could compromise cognitive and behavioral maturation in ASD. Additionally, a reduced expression in PCs of glutamic acid decarboxylase 67 (GAD67) mRNA, an essential enzyme for converting glutamate to GABA, is a consistent finding in the postmortem cerebellum of ASD patients [171][172][201,202]. Conversely, a higher expression of GAD67 mRNA in cerebellar molecular layer interneurons was observed [173][203], suggesting the existence of an upregulation mechanism to counterbalance the altered inhibition of DCN by PCs. Interestingly, the larger-sized subpopulation of GABAergic neurons in the DCN, which project specifically to the IO [96][174][126,204], were reported to exhibit a reduction in GAD65 mRNA expression [175][205]. Thus, GABAergic neurotransmission alteration in DCN could profoundly affect olivary oscillations and subsequently affect the timing of PC activity (Figure 4). To date, accumulating evidence hints for the hypothesis that core features of ASD emerge from disturbances in the E/I balance within neural circuits [49][53][145][176][81,85,175,206]. In conclusion, the above findings highlight the role of reelin, serotonin concentration, GABAergic neurotransmission and GAD enzymes in ASD. However, more investigations are needed to better evaluate the mechanisms underlying E/I balance.
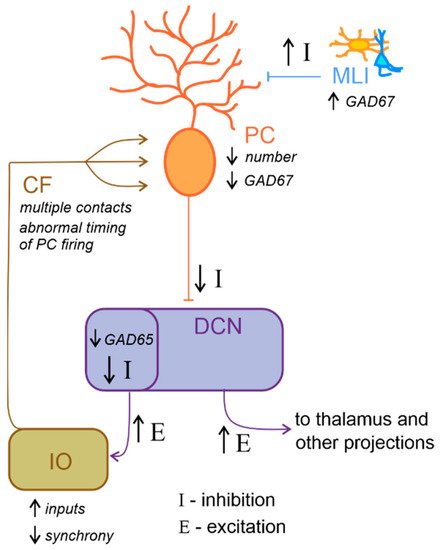
Figure 4. Altered excitatory/inhibitory balance in the cerebellum-inferior olive loop in ASD. Schematic representation of the main alterations described in the cerebellum-inferior olive circuit in ASD, as described in the main text. Briefly, Purkinje cells (PC) are reduced in number and show a decreased level of GAD67 mRNA expression, while molecular layer interneurons (MLI) show an increased inhibition over PC. These would likely determine a decrease in the inhibition (I) over deep cerebellar nuclei (DCN) neurons. DCN disinhibition would increase the excitatory (E) level increasing the output towards the thalamus and other brain regions. Concerning the loop with the inferior olive (IO), DCN neurons which project to this area show decreased GAD65 mRNA expression levels, thus resulting in a decreased inhibition over IO neurons, likely increasing the excitatory inputs and decreasing synchronicity. In ASD, multiple climbing fibers (CF) impinging on the same PC were described. Together with the alterations in IO activity, this anatomical abnormality likely contributes to impair the timing of PC spiking activity.
2.6. Cerebellar Inflammation in ASD
Cells of the immune system and their products are able to directly regulate neuronal function, cell migration, proliferation, adhesion, differentiation [177][207], and synapse formation and plasticity [178][208]. Thus, they play crucial roles in modulating neuronal circuits that constitute the basis for human social cognition and behavior [130][160]. Immune profile alterations have been described in ASD patients [179][180][181][182][183][9,10,11,12,13] and may contribute to the disorders. Postmortem brain tissue from ASD individuals often shows persistent neuroinflammation [184][14]. Specifically, in cerebellar tissue samples, aberrant microglia and astrocyte activation was detected in combination with a massive monocyte and macrophage accumulation, particularly in the granular layer and white matter [184][14]. These findings correlated with marked histological abnormalities including decreased numbers of PCs and GrCs together with reduced numbers of axons within the PC layer [180][184][10,14]. Moreover, in ASD patients, increased levels of many cytokines and chemokines were described in the brain and cerebrospinal fluid, precisely, interleukin (IL)-6, tumor necrosis factor alpha (TNF-α), transforming growth factor beta 1 (TGFβ1), and C-C motif ligand 2 and 17 (CCL2 and CCL17) in the cerebellum [184][185][186][187][14,209,210,211]. Furthermore, antibodies against cerebellar proteins have also been characterized in ASD individuals and are strongly associated with impairments in behaviors, in particular deficits in social interactions and communication [188][189][190][191][212,213,214,215]. The antigenic target of these antibodies has not yet been precisely identified but robust specific reactivity was shown against cerebellar GABAergic interneurons, including Golgi cells [189][192][213,216]. Whether these antibodies alter activity of its target neurons or mark them for destruction by phagocytes requires further investigation. Remarkably, Black and Tan BRachyury (BTBR) inbred mice were identified only fifteen years ago as showing strong and consistent autism-relevant behaviors, including reduced social interactions, impaired play, low exploratory activity, unusual vocalizations and high anxiety [193][194][217,218]. These mice show a number of immunological abnormalities, many of which were described in postmortem brains of ASD subjects [195][196][197][198][219,220,221,222]. They are characterized by elevated cytokine levels in the brain, and an increased proportion of microglial cells. In particular, among the brain regions that Heo and colleagues [195][219] examined, the cerebellum exhibited significantly higher expression of IL-33, IL-18 and IL-6 in BTBR mice than in control, suggesting that it could be a crucial area for neuroinflammation in humans with ASD. Finally, a recent onestudy revealed an abnormal cerebellar development (enhanced foliation and PC hypotrophy with altered dendritic spine formation) concomitant with the progression of motor impairments in BTBR mice [199][223]. In summary, although there is a growing body of evidence supporting the relationship between cytokine alterations and ASD, systematic and large scale investigations are needed to better clarify the role of cerebellar inflammation on the emergence of ASD and the contribution to its etiological heterogeneity.