Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by John Cushman.
Camelina sativa (L.) Crantz., also known as false or wild flax, German sesame, gold-of-pleasure, or linseed dodder, is an allohexaploid (2n = 40) oilseed crop within the Brassicaceae.
- Camelina sativa
- semi-arid lands
- biofuel feedstock
- biodiesel
1. Introduction
Camelina sativa (L.) Crantz., also known as false or wild flax, German sesame, gold-of-pleasure, or linseed dodder, is an allohexaploid (2n = 40) oilseed crop within the Brassicaceae [1,2,3,4,5,6][1][2][3][4][5][6]. Interest in C. sativa increased in recent years due to its adaptability to diverse environmental conditions, low requirements for water and nutrients, relatively strong resistance to insect pests and microbial diseases, and unique oil composition and characteristics suitable for the production of food and fodder, biofuels, and bio-based products [1,2,7,8][1][2][7][8]. These positive agronomic traits and environmental attributes, along with the recent development of methods for transgenesis [9,10,11,12,13][9][10][11][12][13] and CRISPR/Cas genome editing [14[14][15][16],15,16], triggered great interest in C. sativa as an industrial oilseed crop. The ongoing interest in C. sativa is documented by the large number of peer-reviewed publications from various databases retrieved when “camelina” was used as a search term, for example, in a query of the ScienceDirect (2309 publications from 1997–2021), Web of Science (1525 publications from 2000–2021) and Agricola (677 publications from 2000–2021) databases (reported on December 30, 2021). The large number of publications and data reported on C. sativa highlight the immense potential of this crop and the interest in genetic improvement to allow it to gain more widespread acceptance and economic viability. Most publications focus on the uses of C. sativa oil and meal, the composition of oil and fatty acids, its genetics and breeding, its physiology, and its production and agronomic management. Agronomic management refers to the practice of minimizing input factors such as fertilizer, irrigation, tillage, herbicides, fungicides, and insecticides to maximize crop yield outputs such as seed yield, oil content, and biodiesel production. Many reviews report on various aspects of its general use as an oilseed crop [2,4[2][4][17][18][19][20][21][22][23][24],17,18,19,20,21,22,23,24], and as a platform for the production of biofuels [25,26,27,28][25][26][27][28] and industrial lipids [1,6,8][1][6][8]. After providing some essential background, wresearchers summarize the present status of C. sativa research and identify areas for its future improvement with a particular emphasis on enhancing the climate resilience of this highly versatile crop.
2. C. sativa Yield Potential under Hot and Dry Conditions
Global climate change is related to increasing surface aridification and the increasing duration and frequency of droughts in many regions of the world [65,66][29][30]. Interest in C. sativa as a low-input crop for use on marginal lands has grown in recent years [50,67,68,69,70,71][31][32][33][34][35][36]. Thus, recent studies investigated C. sativa grown under semi-arid conditions with limited water inputs, with the goal of assessing its performance under reduced water and fertilizer inputs [36,37,59,62,72][37][38][39][40][41].
Many studies were conducted across semi-arid or arid regions of the globe with distinct mean annual precipitation levels to compare seed yield (kg ha−1), oil content (%), protein content (%), and biodiesel production (L ha−1) (Figure 1, Table 1). Differences in cultivars (genotypes), water availability, environment, physical and chemical characteristics of soils, and management practices such as irrigation, nitrogen application, sowing date, seeding methods and rates, clearly impact overall C. sativa productivity. Among these studies, the overall mean seed yield of C. sativa was 1410 kg ha−1 (Figure 1, Table 1). The highest reported seed yield ranges were attained in Austria (2419–3625 kg ha−1) [73][42] and in southern Ethiopia (2795–3200 kg ha−1) [74][43]. The lowest C. sativa seed yield ranges were reported by studies performed in Kansas [75][44] and Nevada, USA [36,37,59,62][37][38][39][40]. Low seed yield of C. sativa may occur due to drought and high temperatures during the flowering and pod filling stage [61[45][46],76], as well as poor soil quality [77][47]. Consistent with these findings, environmental factors (e.g., temperature and rainfall) were responsible for approximately 73% of the variation in seed yield in north-eastern Poland [72][41]. In contrast, only ~6% of the variation in seed yield was due to genetic factors [72][41]. Mild weather conditions, along with moderate in-season precipitation, favor higher seed yields [35,39][48][49]. Additionally, high C. sativa seed yields were associated with adequate moisture and mild temperatures during seed filling, which is critical for the production and transport of sugars from source to sink tissues [3,77,78][3][47][50]. In contrast, low seed yields were observed under conditions of hot temperature and low precipitation [79,80][51][52]. Studies showed an increase in seed yields ranging from 425 to 2867 kg ha−1 as a result of increasing amounts of applied irrigation ranging from 187 to 536 mm [36,69,70,81][37][34][35][53]. Seasonal water requirements range from 332–490 mm for C. sativa based upon cumulative evapotranspiration (ET) estimates [82,83,84][54][55][56]. Notably, C. sativa might serve as a reliable alternative to Canola in locations where seasonal water supply is less than 250 mm [85][57]. In addition to increased water inputs, increasing nitrogen fertilizer application can increase seed yield. Seed yields of 1800 kg ha−1 can be attained with 150 kg N ha−1 and water inputs in the range of 320–376 mm [86][58]. Similar estimates for optimal N input were reported for rainfed field settings [87,88][59][60]. The use of organic fertilizers during the production of C. sativa for animal feed reduces its environmental impact [89][61].
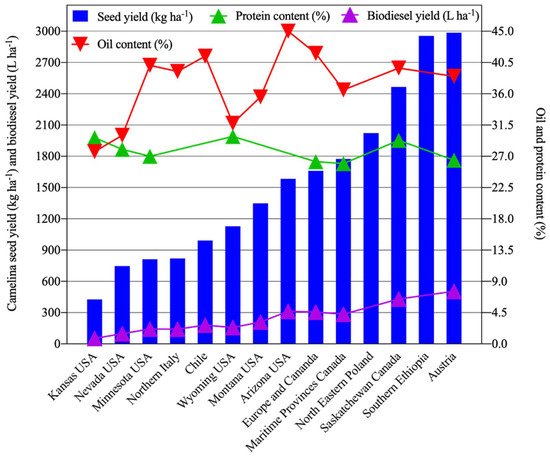

Figure 1.
Average seed yield (kg ha
−1
), oil (%), and protein content (%) of
C. sativa grown at various locations across the world.
grown at various locations across the world.
Another important measure of productivity for any oilseed crop is oil and protein production of the seed meal. For C. sativa, oil and protein content varied widely across various parts of the world, with overall mean seed oil and protein contents of ~36% and ~28%, respectively (Figure 1, Table 1). Highest seed oil contents were reported in the range of 39.8–45.7% in Chile [61][45], followed by 45% in Arizona, USA [84][56], and 38–43% in western Canada [39][49]. The lowest seed oil content, ranging from 25.7 to 31.8%, was observed in Nevada, USA [37][38]. Similarly to seed yield, oil yield is strongly influenced by environmental factors [64,90][62][63]. For example, oil content variation was chiefly associated with mean air temperature and water availability during seed development [61][45].
Table 1.
C. sativa
seed yield, oil, and protein content reported across various regions of the world.
| Locations | Seed Yield (kg ha−1) |
Seed Oil Content (%) 1 |
Seed Meal Protein Content (%) 2 |
Biodiesel Yield (L ha−1) 3 |
Major Sources of Variation | Reference | |
|---|---|---|---|---|---|---|---|
| Mean | Range | Range | Range | Mean | |||
| Austria | 2986 | 2419–3625 | 37.0–40.0 | 25.0–27.9 | 505 | Nitrogen and sulfur rates | [73][42] |
| Southern Ethiopia | 2956 | 2795–3200 | - | - | - | Seeding rates and nitrogen fertilizer | [74][43] |
| Saskatchewan, Canada | 2466 | 2184–2747 | 38 to 43 (39.7) |
27 to 32 (29.3) |
430 | Genotypes and environment | [39][49] |
| North–Eastern Poland | 2023 | 1700–2210 | - | - | - | Genotypes | [72][41] |
| Maritime Provinces of Eastern Canada | 1775 | 1638–1911 | 33.8–39.0 (36.6) |
25.0–26.8 (26.0) |
285 | Breeding lines, nitrogen and sulfur rates | [87][59] |
| Europe and Canada | 1660 | 1100–2700 | (41.8) | (26.2) | 305 | Genotypes and environment | [77][47] |
| Arizona, USA | 1583 | 1527–1638 | (45) | - | 313 | Nitrogen rates, water use and irrigation scheduling | [84][56] |
| Montana USA | 1349 | 546–2942 | 33.5–37.6 (36.0) |
- | 211 | Cultivars and locations | [78][50] |
| Wyoming, USA | 1129 | 832–1643 | 31.1–32.4 | 29.3–30.4 | 157 | Nitrogen and sulfur rates | [91][64] |
| Chile | 991 | 387–2314 | 39.8–45.7 (41.41) |
- | 180 | Cultivars, planting dates, and locations | [61][45] |
| Northern Italy | 820 | 600–940 | (39.2) | 141 | Low input and growing seasons | [71][36] | |
| Nevada, USA | 899 | 770–1013 | 31.8–33.3 (32.4) |
27.1–28.1 (27.5) |
128 | Cultivars and irrigation rates | [36][37] |
| 784 | 534–1010 | 31.8–32.6 (32.2) |
26.2–30.4 (28.4) |
111 | Cultivars, nitrogen sources and rates | [59][39] | |
| 735 | 34–1921 | 26.6–30.8 | - | 69.7 | Cultivars and year | [37][38] | |
| 570 | 130–921 | 28.4–29.5 | - | 72 | cultivars, sowing date, and methods | [62][40] | |
| Minnesota, USA | 812 | 650–944 | 39.4–40.7 (40.1) |
26.2–27.9 (27.0) |
143 | Genotypes and seeding rate | [3] |
| Kansas, USA | 427 | 317–503 | 27–29 (27.7) |
29–30 (29.7) |
52 | Cultivar and planting date | [75][44] |
| Overall mean | 1410 | 36.0 | 27.8 | 208.4 | |||
1 Oil and 2 protein content inside brackets indicate the average oil and protein content (%). 3 Biodiesel yield was estimated by multiplying seed yield (kg ha−1) and oil content (%) using the volumetric conversion factor of 1 kg ha−1 to 0.439 L ha−1 [92][65].
Compared with seed oil values, protein content generally varied less widely relative to the location of production across the globe, with the lowest and highest ranges of 25.0–26.8% and 27–32%, respectively, reported in Canada, with an overall mean of 27.8% (Figure 1, Table 1). However, more wide-ranging protein contents (23–47%) of C. sativa seeds were reported depending upon local field conditions [60,77][66][47]. High air temperatures during flowering and seed filling are generally associated with low oil contents but high protein contents within C. sativa seed [62][40]. Furthermore, increased N application rates can increase protein contents, with an associated decrease in oil contents [73,87,93,94][42][59][67][68]. Such increased nitrogen applications are known to increase protein at the expense of fatty acid synthesis due to the competition for carbon skeleton during carbohydrate metabolism [95][69].
A major use of C. sativa seed oil is biodiesel production. Biodiesel production, which is closely linked with overall oil production, varied widely with a mean of 208 L ha−1 across the globe (Figure 1, Table 1). The highest biodiesel yield (505 L ha−1) was reported in Austria [73][42], followed by western Canada (430 L ha−1) [39][49]. A multi-location study performed in Europe and Canada reported a mean biodiesel yield of 305 L ha−1 [77][47]. The lowest biodiesel yield (52 L ha−1) was reported in Kansas, USA [75][44]. The wide variation in biodiesel production is likely accounted for by differences in temperature and water availability, including ambient precipitation, length of growing season, and managerial factors such as fertilizer application rates and irrigation practices.
3. Biotic Production Constraints of C. sativa
Weeds are a major constraint to C. sativa production (Berti et al., 2016). C. sativa is considered to be very competitive against weeds with good stand establishment [22,96][22][70], due to its ability to produce and release secondary metabolites that prevent the growth of seedlings of neighboring plants [97,98,99][71][72][73]. However, the competitiveness of C. sativa for sunlight, nutrients, and water can be challenged by some weed species such as cheatgrass (Bromus tectorum), green foxtail (Setaria viridis L.), and Russian thistle (Salsola kali L.), among others [2,5,100,101][2][5][74][75]. Similarly, perennial broadleaf weeds such as field bindweed (Convolvulus arvensis L.), Canada thistle (Cirsium arvense L. Scop.), and skeleton weed (Chondrilla juncea L.) can also present challenges for C. sativa. Notably, C. sativa can exhibit susceptibility to dodder (Cuscuta spp.), a parasitic weed of many crops (D. Neupane, personal observation). C. sativa does not hybridize efficiently with cultivated Brassica species or wild Brassica species within the tribe Camelineae [102][76]. However, C. sativa does exhibit the ability to outcross with common Brassica weed species, such as North American weeds C. alyssum and C. microcarpa [103][77]. Fortunately, C. sativa is less competitive and shows lower invasive potential than Canola [104][78].
3.1. Weed Control
Both chemical and non-chemical approaches for weed control are available for C. sativa production systems. For chemical control, Sethoxydim (Poast®) (2-[(E)-N-ethoxy-C-propylcarbonimidoyl]-5-(2-ethylsulfanylpropyl)-3-hydroxycyclohex-2-en-1-one), a post-emergent and selective grass herbicide, which belongs to the cyclohexanone group, is currently the only registered herbicide for C. sativa in the USA [59,62,105,106][39][40][79][80]. Tepraloxydim (2-[(E)-N-[(E)-3-chloroprop-2-enoxy]-C-ethylcarbonimidoyl]-3-hydroxy-5-(oxan-4-yl)cyclohex-2-en-1-one) was also used to control post-emergent grass weeds [61][45]. Similarly, Clethodim (2-[(E)-N-[(E)-3-chloroprop-2-enoxy]-C-ethylcarbonimidoyl]-5-(2-ethylsulfanylpropyl)-3-hydroxycyclohex-2-en-1-one), which is a related post-emergent cyclohexanone herbicide, was successfully used for grassy weed control for C. sativa [36][37]. Quizalofop (2-[4-(6-chloroquinoxalin-2-yl)oxyphenoxy]propanoic acid), which is a selective, postemergence phenoxy herbicide registered in Canada and other parts of the world, was also used to control annual and perennial grass weeds of C. sativa [2,5,107][2][5][81].
In contrast to selective grass herbicides, C. sativa withstands very few broadleaf herbicides, and few have been approved for use on C. sativa in the USA [2,5,22,107,108][2][5][22][81][82]. Dinitroaniline pre-emergent herbicides, such as Trifluralin (2,6-dinitro-N,N-dipropyl-4-(trifluoromethyl)aniline), are widely used in the USA for early season weed control. Pre-emergent herbicides, including dimethenamid-P (2-chloro-N-(2,4-dimethylthiophen-3-yl)-N-[(2S)-1-methoxypropan-2-yl]acetamide), pendimethalin (3,4-dimethyl-2,6-dinitro-N-pentan-3-ylaniline), pyroxasulfone (3-[[5-(difluoromethoxy)-1-methyl-3-(trifluoromethyl)pyrazol-4-yl]methylsulfonyl]-5,5-dimethyl-4H-1,2-oxazole), quinclorac (3,7-dichloroquinoline-8-carboxylic acid), and S-metolachlor (2-chloro-N-(2-ethyl-6-methylphenyl)-N-[(2S)-1-methoxypropan-2-yl]acetamide), were tested at three different concentrations with C. sativa to examine weed control and injury to the crop [109][83]. Pendimethalin and S-metolachlor caused some injury, but did not lower C. sativa seed yields. Dimethenamid-P, used at a low rate, did not affect plant populations and seed yield; however, when applied at a high rate, it caused 60% injury with a 31% reduction in seed yield [109,110][83][84]. Quinclorac did not significantly injure the crop and was considered the safest of all the herbicides tested [109][83]. C. sativa mutants with resistance to acetolactate synthase (ALS) inhibitors were developed, displaying increased resistance to imazethapyr (5-ethyl-2-(4-methyl-5-oxo-4-propan-2-yl-1H-imidazol-2-yl)pyridine-3-carboxylic acid), sulfosulfuron (1-(4,6-dimethoxypyrimidin-2-yl)-3-(2-ethylsulfonylimidazo[1,2-a]pyridin-3-yl)sulfonylurea), and flucarbazone (3-methoxy-4-methyl-5-oxo-N-[2-(trifluoromethoxy)phenyl]sulfonyl-1,2,4-triazole-1-carboxamide) [110][84].
Several non-chemical strategies for weed control were shown to be effective for C. sativa [96][70]. Mechanical weeding is a well-established and useful approach for small scale production systems (e.g., research plots). For large-scale production, early season planting that promotes stand establishment is one of the most effective strategies for overcoming weed competition in C. sativa fields [111][85]. Furthermore, increasing sowing rates can effectively suppress weeds in C. sativa plots, likely through increased utilization of resources by the crop at the expense of weeds [96][70]. Mixed cropping of C. sativa grain with peas (Pisum sativum) (or barley (Hordeum vulgare)) afforded significant weed suppression and acted as a smother crop and weed antagonist, leading to enhanced C. sativa seed yields [96,112][70][86].
Use of C. sativa as a winter cover crop suppressed weed abundance while increasing cash crop yields [113][87]. Including C. sativa in crop rotations for winter cereals can suppress winter weed populations and increase grain yields in semi-arid Mediterranean climates [114][88]. C. sativa was also used in innovative double- and relay-cropping systems with forage sorghum (Sorghum bicolor), maize (Zea mays), and soybean (Glycine max) with an increased potential for biofuel and energy feedstock production [115][89]. When C. sativa was intersown as a cover crop with maize or soybean, establishment and winter survival rates increased, with the greatest success being achieved with soybean [116][90]. C. sativa was also used as an intercrop with Jatropha integerrima in semi-arid regions of India with seed production improvements observed when grown alternatively with leguminous fodder crops [117][91]. Further exploration of the potential allelopathic effects of C. sativa in intercropping systems for weed suppression is needed [99][73].
3.2. Insect Pests
Insects can limit C. sativa production, but these generally present lower production barriers than weeds. C. sativa is known to be relatively resistant to insect predation and infestation. C. sativa shows resistance to several common insect pests of Canola [118,119][92][93]. For example, C. sativa shows high resistance to flea beetles (Phyllotreta spp.), a common insect pest of B. rapa, B. napus, and B. juncea [39,49,120,121][49][94][95][96]. This resistance is thought to be due to the presence of repellents or the absence of stimulatory volatile phytochemicals [122][97]. More recent studies confirmed that the presence of quercetin glycosides in C. sativa leaves is likely responsible for flea beetle resistance [123][98]. In addition to flea beetles, C. sativa was shown to be a poor host for the diamondback moth (Plutella xylostella L.) and root maggots (Delia spp.) [124][99]. Feeding damage by the Bertha armyworm (Mamestra configurata Walker) on C. sativa was consistent with Canola, but larvae and pupae weighed less when reared on C. sativa leaves [124][99]. Leafhopper (Macrosteles quadrilineatus Forbes) did not cause damage to C. sativa; however, this insect can transmit aster yellows phytoplasma, which can result in chronic, systemic disease resulting in chlorosis, phyllody, and virescence, malformed seeds, and reduced seed set [124][99]. C. sativa also appeared to show resistance against the cabbage seedpod weevil (CeutorhyncusCeutorhyncus obstrictus obstrictus (Marsh.) [125][100]. However, stem feeding below the soil by several weevil species (e.g., Ceutorhynchus cyanipennis and C. americanus) was associated with stand failures for fall-planted C. sativa [126][101]. Although aphids can colonize C. sativa under field conditions, they are not known to cause significant economic damage; however, such colonization can create a reservoir for different aphid species that can negatively impact other crops in the rotation [127][102].
3.3. Fungal, Bacterial, and Viral Pathogens
Similar to its apparent resistance to many insect pests, C. sativa shows good resistance to a variety of microbial pathogens that typically target crops within the Brassicaceae [18,128][18][103]. For example, C. sativa showed strong resistance to blackspot fungal disease caused by Alternaria brassicae (Berk.) Sacc. [129,130,131][104][105][106]. Such resistance is associated with the relative expression of defense-related genes [132][107]. Resistance to blackspot disease caused by Alternaria brassicicola [(Schw.) Wiltsh] varies among C. sativa genebank accessions [133][108]. However, such resistance can be transferred to hybrids of C. sativa and rapid-cycling Brassica oleracea in an effort to confer resistance of this disease to Brassica vegetable crops [134,135][109][110]. Resistance to the blackspot pathogen was associated with the production of phytoalexins, specifically to camalexin, an antimicrobial phytoalexin [129,131,134,136][104][106][109][111]. Resistance was also correlated with the higher expression of chitinase gene family members in C. sativa relative to B. juncea [137][112]. C. sativa showed strong resistance to blackleg fungal disease [39][49] assessed using 80 different isolates of Leptosphaeria maculans (Desmaz.) [138][113]. However, for those C. sativa genotypes that are susceptible to blackleg disease, the fungicides metoconazole and a mixture of tebuconazole and triadimenol provided very effective control against the fungus [61][45].
C. sativa genotypes show various, but low, degrees of susceptibility to common Brassicaceae diseases, such as stem rot (Sclerotinia sclerotiorum (Lib.) de Bary), brown girdling root rot (Fusarium spp. and R. solani), downy mildew (Peronospora sparsa, P. parasitica), powdery mildew (Erysiphe sp.), grey mold (Botrytis cinerea), sore shin and damping-off disease (Rhizoctonia solani), Verticillium wilt (Verticillium dahliae), and white rust (Albugo candida) [18,128,139,140][18][103][114][115]. Some C. sativa genotypes are susceptible to downy mildew (Hyaloperonospora camelinae) [141][116]. Thirty diverse C. sativa genotypes showed varying degrees of resistance to S. sclerotiorum [142][117]. Resistance to S. sclerotiorum was associated with cell wall strengthening due to monolignol biosynthesis in C. sativa [143][118]. However, while camalexin production was induced by S. sclerotiorum inoculation, the relative degree of disease resistance was not correlated with levels of camalexin production, suggesting that other antimicrobial activities might be responsible for the observed disease resistance [142][117]. C. sativa also shows resistance to the soil-born fungus Fusarium virguliforme [144][119]. However, C. sativa is susceptible to other fungal diseases common to the Brassicaceae, such as damping-off (caused by Rhizoctonia solani), clubroot (Plasmodiophora brassicae Woronin.), and white rust (Albugo candida Pers. Kuntze) [128][103]. While susceptible, C. sativa showed greater resistance to Rhizoctonia solani than B. napus did, due to the presence of the phytoalexins camalexin and methoxycamelexin and two additional antimicrobial compounds in its roots [145][120].
C. sativa is susceptible to some bacterial diseases, such as bacterial blight (Pseudomonas syringae pv. spec.), phytoplasma disease, which was reported for C. sativa grown in Germany and Canada [139[114][115][121],140,146], and aster yellows diseases [124][99]. C. sativa also shows susceptibility to the bacterial pathogen Xanthomonas campestris Dowson pv. campestris, which causes black rot of Brassica crops globally [133][108].
Lastly, although relatively less well studied for viral diseases, C. sativa is susceptible to the Turnip crinkle virus and Turnip rosette virus [147][122], which are viral diseases transmitted by flea beetles via infested seed [2,148][2][123]. The beet western yellows mosaic virus reportedly caused up to 34% yield reductions in C. sativa [149][124]. Aphid (Myzus persicae) vector behavior and host palatability are key factors in the transmission of the Turnip yellows virus in C. sativa [150][125].