Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 3 by Beatrix Zheng and Version 2 by Beatrix Zheng.
Hepatocellular carcinoma (HCC) is the most common primary liver cancer with an increasing worldwide mortality rate. Cholangiocarcinoma (CCA) is the second most common primary liver cancer. In both types of cancers, early detection is very important. Biomarkers are a relevant part of diagnosis, enabling non-invasive detection and control of cancer recurrence, as well as in the application of screening tests in high-risk groups. Furthermore, some of these biomarkers are useful in controlling therapy and treatment selection. Detection of some markers presents higher sensitivity and specificity in combination with other markers when compared with a single detection. Some gene aberrations are also prognostic markers in the two types of cancers.
- biomarker
- genetic marker
- hepatocellular carcinoma
- cholangiocarcinoma
- screening
- diagnosis
- therapy
- treatment
1. Introduction
Hepatocellular carcinoma (HCC) is the most common primary liver cancer (representing 75–85% of cases) and the fourth most common cause of cancer-related deaths in the world [1]. The mortality rate has been increasing for the past several dozens of years worldwide [2]. Although morbidity rate trends have recently been dropping in traditionally high-risk regions such as parts of Asia, they have lately increased in others such as North America and Europe [3]. The risk factors also depend on the region, but overall, the primary risk factor is liver cirrhosis (regardless of its etiology). More precisely, over a half (54%) of cases of HCC are caused by viral-related liver cirrhosis (including hepatitis B virus (HBV) with 33%, and hepatitis C virus (HCV) with 21%) while the rest of the cases include mostly alcoholic liver cirrhosis and non-alcoholic liver cirrhosis, the latter a consequence of the progressive form of non-alcoholic steatohepatitis (NASH) [1]. Other less common causes of HCC include various carcinogens such as nitrites, hydrocarbons, aflatoxins, and organochlorine pesticides, and genetic disorders including hemochromatosis, Wilson’s disease, and alfa-1 antitrypsin deficiency [4] (Figure 1).
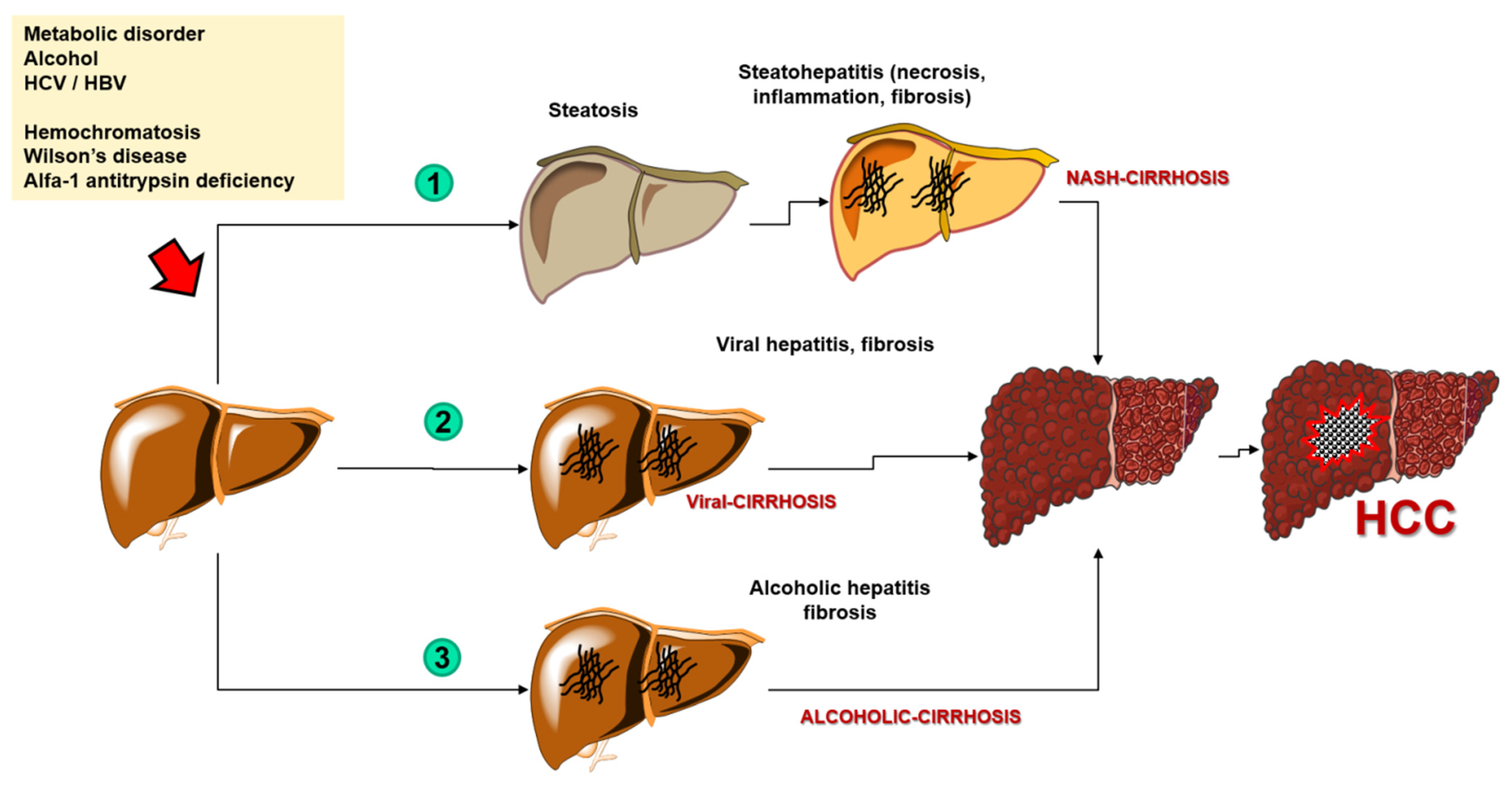
Figure 1. The process of carcinogenesis of the hepatocellular carcinoma. Abbreviations: HCC—hepatocellular carcinoma; NASH—nonalcoholic steatohepatitis; HCV—hepatitis C virus; HBV—hepatitis B virus.
Cholangiocarcinoma (CCA) is the second most common primary liver cancer. The anatomical classification contains intrahepatic (iCCA) and extrahepatic (perihilar–pCCA and distal–dCCA) CCAcholangiocarcinoma. CCA shares several similarities with HCC by virtue of its increased incidence rate and its higher prevalence in Asian countries [2][5]. Mortality rates vary depending on the type of CCA, whether it comprises an increased amount of iCCA and decreased amount of extrahepatic CCA [6]. Overall, the prognosis of CCA is poor—a 5-year survival rate equals only 10%. Most of the cases of CCA are sporadic, although currently determined main risk factors include ‘biliary duct disorders, parasitic infections, and toxins and viral hepatitis B and C’, so that, along with liver cirrhosis, there is another resemblance to HCC. The main diagnostic and staging tests for CCA consist of high-quality cross-sectional imaging (CT or MRI), although diagnostic methods may be different for each type of malignancy [7]. These other methods include imaging tests such as abdominal ultrasound, positron emission tomography (PET), percutaneous transhepatic cholangiography, and endoscopic retrograde cholangiopancreatography (ERCP) or endoscopic ultrasound; or non-radiological methods represented by histological analysis and non-specific biomarkers. However, difficult early-stage detection and hard to define risk factors currently result in a high mortality rate. The diagnosis of CCA is often made late, limiting the effectiveness of surgical treatment that is the only efficient method of therapy [8]. Therefore, the need for specific and non-invasive biomarkers which may facilitate an early diagnosis and possible curative treatment of CCA has led to promising studies discovering such markers in various types of tissue and bodily fluids [9]. Due to the anatomy of the biliary tract system, and to the typically small size of CCA lesions, a procedure of obtaining tissue samples using ERCP presents several limitations. ERCP with intraductal ultrasonography (IDUS)-guided needle aspiration is linked to a risk of biliary tract perforation, and potentially severe biliary hemorrhage. The use of circulating nucleic acids, available in most biofluids is reported as a promising solution, providing tumor genomic information in a less invasive way, using plasma or bile samples [10].
2. Genetic Markers for HCC
2.1. MicroRNAs
MicroRNAs is a type of non-coding RNA that presents an 18–25 nucleotide length. It has a major role in regulating posttranscriptional gene expression, and recently it has been shown that miRNAs can indicate diagnosis and prognosis of different cancer types [11][12]. While obtaining miRNAs from tissue samples is an invasive procedure, it has been shown that, similarly to previous nucleic biomarkers, miRNAs can be available from serum [13].
It is now believed that incorrect miRNA expression plays an important role in the carcinogenesis of HCC [14]. Multiple types of miRNAs are potentially useful in HCC diagnostics, e.g., miRNA-224, whose expression level is higher in HCC cells than in normal hepatocytes and which affects the cell proliferation rate and cancer invasiveness. Mir-224 concentration is higher in the serum of HCC patients and its levels correlate with survival time—the higher the concentration in the serum, the shorter survival time [15]. Overexpression of microRNA-766 promotes proliferation and metastases, and also cell migration. The elevated expression level of microRNA-766 is a negative predictor of HCC treatment, and miRNA-766 itself could be potentially useful in treatment and diagnostics [16]. MicroRNA-23 is an important oncogene for HCC progression and its high levels are detectable in patients’ serum so that it can be used as a marker of HCC. Produced in adipocytes, microRNA-23 promotes cancer cells’ proliferation, and inhibition of its uptake might be a target for inhibiting the progression of HCC [17]. MiR-494 promotes metastasizing and cancer invasiveness and it can also act as a biomarker for predicting the risk of reoccurrence in AFP-negative HCC patients. In this case, inhibition of expression also seems to be a target in the therapy of HCC [18]. In turn, measuring the levels of three miRNAs at once: miR-10b, miR-106b, and miR-181a is characterized by a high sensitivity and specificity, and this enables their usage for the HCC screening test [19].
2.2. Genetic Markers
Genomic analyses may enable better characteristics of a tumor, which will allow for treatment optimization of patients with HCC, and therefore research on HCC genetic markers is being conducted all over the world. The genes which are most subject to mutations in HCC are catenin beta-1 (CTNNB1) and tumor protein P53 (TP53) [20].
CTNNB1 gene encodes β-catenin which plays an important part in cell adhesion and communication. A correlation between CTNNB1 gene mutation and alcoholic HCC has been shown [20]. It has been stated that the inactivation of chromatin remodelers was dominant in tumors related to alcohol. An association of mutations in ribosomal protein S6 kinase A3-Axin 1 (RPS6KA3-AXIN1) and NFE2 like BZIP transcription factor 2-catenin beta-1 (NFE2L2-CTNNB1) genes suggest that Wnt/β-catenin signaling pathway could take part in hepatic cancerogenesis. It could affect oxidative stress metabolism and Ras/mitogen-activated protein kinase (MAPK) pathways [21]. TP53 is a tumor suppressor gene encoding a protein that takes part in many cellular processes, especially in the activation of DNA repair mechanisms or in the induction of apoptosis in response to DNA damage. Whereas in the case of TP53 mutation the cells with damaged DNA might avoid apoptosis and transform into cancer cells. A correlation has been shown between TP53 mutation and HBV-related HCC. Inactivation of p53, either through mutation or binding to other viral and cellular oncoproteins, is often observed in neoplasms. Such interaction has been described in the case of p53 and the hepatitis B virus X gene (HBx) since HBx can inactivate the apoptosis which is mediated by p53 [22]. In tumors related to hepatitis B virus (HBV), there have also been shown suppressor properties of interferon regulatory factor 2 (IRF2), inactivation of which has led to a disabled function of TP53 [21]. It has also been proven that TP53 mutation can cause an inhibition of immune response in HCC [23]. The next genes regarding the frequency of mutations in HCC are AXIN1 and AT-rich interaction domain 1A (ARID1A) [24].
AXIN1 gene, being a negative regulator of the Wnt/β-catenin signal pathway, undergoes loss-of-function mutations. Research has shown that AXIN1 mutations occur independently of Wnt/β-catenin pathway and they correlate with Notch and YAP pathways which can represent interesting targets in HCC treatment [25].
It has been found that decreased ARID1A expression level is associated with tumor progression, metastasis, and reduced survival [26]. It has been proven that patients with mutations in the ARID1A gene have a poor prognosis [27]. It also turned out that ARID1A mutation, along with CTNNB1, telomerase reverse transcriptase (TERT), and SWI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily A, member 2 (SMARCA2) mutations occur in alcohol-related HCC [28].
Another gene undergoing a mutation in HCC is cyclin dependent kinase inhibitor 2A (CDKN2A). It encodes two proteins that work as cancer suppressors through cell-cycle regulation. The most common mechanism of inactivation of the CDKN2A gene in HCC is homozygous deletions [29]. Moreover, it has been stated that CDKN2A inactivation and fibroblast growth factor/cyclin D1 (FGF/CCND1) amplification correlated with poor prognosis [20] (Figure 2).
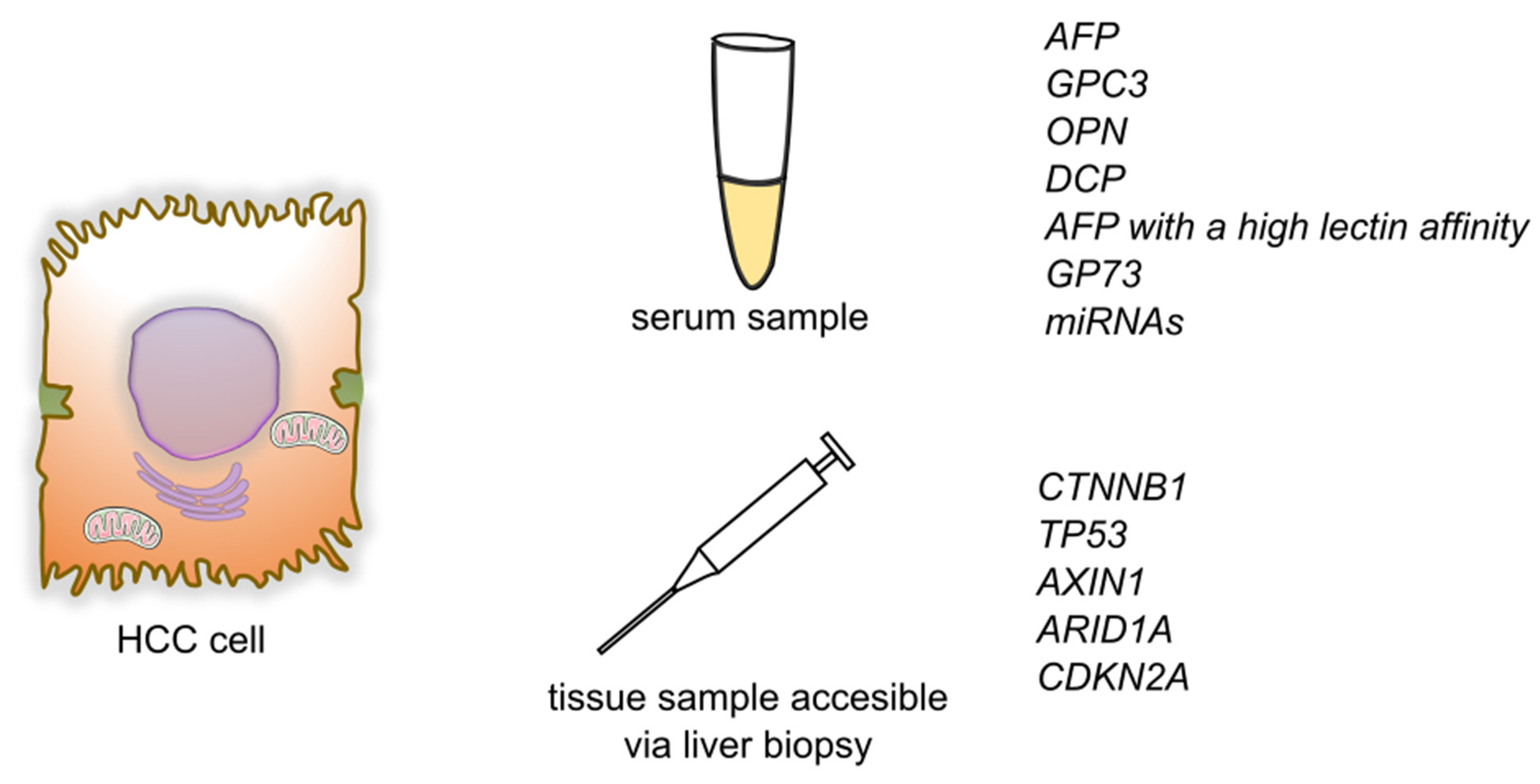
Figure 2. Summary of detection of biomarkers and genetic markers for hepatocellular carcinoma. Abbreviations: AFP—alphafetoprotein; GPC3—glypican-3; OPN—osteopontin; DCP—des-γ-carboxy prothrombin; GP73—Golgi protein-73; miRNAs—micro RNAs; CTNNB1—Catenin beta-1; TP53—Tumor protein P53; AXIN1—Axin 1; ARID1A—AT-rich interaction domain 1A; CDKN2A—cyclin dependent kinase inhibitor 2A.
3. Genetic Markers of CCA
3.1. Circulating Nucleic Acids
Cell-free DNA (cfDNA) and Cell-free RNA (cfRNA) are potential diagnostic and prognostic markers that can be found in most biofluids. They appear in the circulating system in two ways—they can be actively exported or originate from dying cells [9]. cfDNA could potentially be a less invasive way to provide genetic information, compared to tissue sampling.
3.1.1. Cell-Free DNA
cfDNA was first identified in blood samples in 1948 by Mandel and Métais [30]. It was used as a potential biomarker of neoplasm in the 1970s showing a link with neoplasm aggressiveness and size [9].
Symptoms of CCA and other biliary diseases are similar. Misdiagnosis of the mentioned diseases could be prevented by the usage of specific biomarkers. Wasenang et al. [31] investigated the potential serum of cfDNA biomarkers that could be used to differentiate CCA and other biliary conditions. In their study, they found that methylation levels of opioid binding protein/cell adhesion molecule like (OPCML) and homeobox D9 (HOXD9) differed significantly between CCA and other biliary diseases. Assessment of the biomarkers showed that sensitivity and specificity of OPCML were 80.0% and 90.0%, respectively, while in the case of HOXD9 it was 67.5% and 90%, respectively. A combined marker between OPCML and HOXD9 showed sensitivity and specificity of 62.50% and 100%, respectively (AUC: 0.812). The authors suggest that OPCML and HOXD9 methylation measurement could be a beneficial method for minimally invasive, early differentiation of CCA and other biliary diseases.
In another study, a comparison was made between bile cfDNA and tissue sampling in detecting bile tract tumors mutations. Each of 10 patients (4 with gallbladder carcinoma (GBC) and 6 with CCA) provided a tumor tissue sample and paired bile sample. A panel of 150 tumor-related genes was used to analyze mutations individually in tissue and bile samples and then compared using targeted deep gene sequencing. In the aspect of single nucleotide variation (SVC) or insertion and deletion (Indel), the bile cfDNA showed high sensitivity and specificity (94.7%, and 99.9%, respectively) compared with the tissue sampling results. In the aspect of copy number variation (CNV) the cfDNA targeted deep sequencing had a sensitivity and specificity of 75.0%, and 98.9%, respectively. The authors suggest that targeted deep sequencing of bile cfDNA can be an accurate and sensitive method in detecting mutational variations of biliary tract carcinomas [10]. In a similar study, Driescher et al. [32] investigated the use of bile and serum cfDNA as a biomarker of extrahepatic CCA and pancreatic ductal adenocarcinoma (PDAC). Similarly, the results were compared to the results from matched tumor samples. In their study, sequencing of bile cfDNA showed a 96.2% concordance with tumor tissue samples with sensitivity and specificity of 100% and 100%, respectively. On the other hand, sequencing of plasma cfDNA revealed a concordance of 31.6%, compared to tumor tissue samples, and a concordance of 48%, compared to sequencing bile cfDNA. The authors suggest that sequencing of bile cfDNA might be a beneficial, ancillary diagnostic method in the diagnosis of pancreatobiliary cancers, while plasma cfDNA performs less reliably.
Wang et al. [33] evaluated the value of plasma CNV assays in the diagnosis of biliary tract carcinomas. Their prospective study was held among 47 patients with suspicious biliary lesions, of which 21 were diagnosed with CCA, and another 8 were diagnosed with GBC. In this research, CNV assays had a sensitivity and specificity of 89.7%, and 88.9%, respectively, while CA 19-9 had a sensitivity and specificity of 58.6%, and 72.2%, respectively. AUC of CNV assays was 0.91, which significantly outperformed CA 19-9, whose AUC was 0.62. Additionally, the authors showed that a higher CNV ratio was associated with decreased survival.
3.1.2. Cell-Free RNA
Exosomes are a type of extracellular vesicles with a diameter of 30 to 150 nm containing molecules including DNA, mRNA, and different types of non-coding RNA, including piwi-interacting RNAs (piRNAs) [34]. Exosomes are excreted into the circulating system and thus are easily accessible during routine blood drawing [34].
piRNAs are a type of small, non-coding RNA with an ability to bind to PIWI family protein specifically. In their study, Gu et al. [34] investigated the potential role of plasma piRNAs in a diagnosis of the CCA and (GBC) in the group of 5 CCA and 4 GBC patients. It was shown that 694 and 323 piRNAs were upregulated in CCA and GBC, respectively, among which 163 molecules were upregulated concordantly. Thirty-six and 191 piRNAs were downregulated in CCA and GBC patients, respectively, among which 25 were downregulated both in CCA and GBC. On the other hand, piR-4333713 was significantly downregulated for GBC and significantly upregulated in the CCA, suggesting differences between CCA and GBC in the exosomal signature. Additionally, the authors found that plasma levels of piR-10506469 and piR-20548188 decreased significantly after surgery for both CCA and GBC, and plasma levels of piR-20548188 were correlated with CCA malignancy grade. Concluding, the authors suggest that exosomal piRNAs may have promising diagnostic values in the diagnosis of CCA and GBC.
3.1.3. Cell-Free Long Non-Coding RNA
Recent studies have provided evidence that long non-coding RNAs (lncRNAs) play a role in carcinogenesis [35]. Bai et al. [35] in their study investigated the role of colon cancer-associated transcript 2 (CCAT2) in CCA. In their study, they found that CCAT2 was overexpressed in 70.8% of CCA tissue samples, compared to adjected non-malignant tissue. The AUC for overall survival and progression-free survival was 0.702, and 0.715, respectively, suggesting that CCAT2 may be useful as a prognostic biomarker. High expression of CAAT2 was also linked to clinical features including microvascular invasion, differentiation grade, CCA progression, and metastasis. The authors suggest that CCAT2 may be a promising prognostic factor and therapeutic target in the CCA.
In another study concerning lncRNAs as prognostic factors, Angenard et al. [36] showed that 9 of the concerned lncRNAs were significantly correlated with overall survival and disease-free survival. Four of them: (cyclin-dependent kinase 9 (lnc-CDK9-1), XLOC l2 009441, CDKN2B antisense RNA 1 (CDKN2B-AS1), HOXC13 antisense RNA (HOXC13-AS) were highly expressed in case of poor-prognosis iCCA, and the remaining 5 (long non-coding Coiled-Coil Alpha-Helical Rod Protein 1 (lnc-CCHCR1-1), lnc-AF131215.3.1, long non-coding Cbl Proto-Oncogene B (lnc-CBLB-5), COL18A1 antisense RNA 2 (COL18A1-AS2), long non-coding RELT like 2 (lnc-RELL2-1)) were upregulated in the case of better prognosis. The authors showed that CDKN2B-AS1 was related to poor prognosis also in the other types of tumors, including HCC and kidney renal clear cell carcinoma.
3.1.4. Micro RNA
Micro RNA has been studied as a marker of HCC, however, it has been also suggested as marker of CCA. miR-21 is one of the best evaluated miRNAs in the CCA [9]. It has been shown that in the serum of CCA patients, the expression of miR-21 was significantly upregulated and related to clinical features, including clinical stage, lymph vessel infiltration, and metastasis status. Compared with the low miR-21 expression group, in the high miR-21 expression group, values of overall survival and progression-free survival were significantly lower. Additionally, it has been found that miR-21 levels decreased significantly after tumor operation. On the other hand, the receiver operating characteristic curve (ROC) analysis suggested that serum miR-21 is defective in distinguishing CCA patients from healthy controls with an area under curve (AUC) value of 0.871. Its diagnostic results, including the sensitivity of 66.7% and specificity of 99.33%, were worse than CA 19-9 results (91.7%, and 99.33%, respectively). What is more, no significant difference was shown between CCA TNM Classification of Malignant Tumors (TNM) stage I patients and healthy volunteers or hepatolithiasis patients. The authors suggest that serum miR-21 is an effective prognostic, but not a diagnostic marker of CCA [11].
MiR-885-5p is another miRNA molecule that is decreased as part of many malignancies, including pancreatic cancer and HCC [12]. It has a role in inhibiting tumor progression by reducing the expression of several genes including hexokinase 2, astrocyte elevated gene-1 (AEG1), and CTNNB1 [12]. Lixin et al. [12] investigated the role of miR-885-5p in the tissue samples of 33 iCCA patients. Comparing with tissue samples of healthy individuals, miR-885-5p expression was aberrantly decreased in the CCA samples. Downregulation of miR-885-5p was also associated with clinical features including vascular invasion and lymph node metastasis and with shorter disease-free and overall survival. In a nude mice model with injected iCCA cells, overexpression of miR-885-5p reduced the incidence of lung and liver metastasis, while miR-885-5p inhibition acted inversely. The authors suggest that miR-885-5p could be a beneficial prognostic marker and therapy target in iCCA management.
In another study, Yao et al. [37] showed that, based on sequencing results, 83 of the miRs were upregulated and 45 of the miRs were downregulated in the samples of 36 CCA patients compared with 9 control samples. In particular, miR-3913-5p was highly expressed in the malignant tissues and high miR-3913-5p expression was identified as an independent prognostic factor of lower overall survival. It has also been shown that ring finger protein 24 (RNF24) and sialic acid binding Ig like lectin (SIGLEC), presumably involved in promoting proliferation and metastasis might be the target genes of miR-3913-5p.
Another miRNA molecule, a miR-130a-3p was studied by Asukai et al. [38]. Results of the research suggest that miR-130a-3p may be associated with gemcitabine resistance among patients with CCA. The authors also showed that peroxisome proliferator activated receptor gamma (PPARG) gene is a target of miR-130a-3p and pioglitazone, which is a PPARγ activator, alleviated gemcitabine resistance and had a synergic effect with gemcitabine.
3.2. Genetic Markers
The advent of new genome analysis technologies played a major role in the process of understanding tumor pathogenesis and heterogeneity. In a large study conducted among 489 CCA patients from 10 countries, Jusakul et al. [39] analyzed genetic features of CCA, including the whole genome sequencing (WGS) (n = 71), and DNA methylation (n = 138) assessment. In the WGS analysis, a total of 1,309,932 mutations were detected across 71 tumor samples, including 4541 nonsilient single nucleotide variations (SNVs) and 1251 nonsilient indels. On average, each tumor had 82 nonsilient mutations, including 64 SNV and 18 indels. Fluke-positive CCAs were significantly more plentiful in somatic mutations comparing with Fluke-negative tumors (median of 4700 vs. 3143 per tumor, respectively). Fluke infection was also correlated with poorer survival. Based on the results of their analysis, the authors suggest dividing CAA cases into 4 molecular clusters:
Cluster 1 was characterized by mostly Fluke-positive CCAs with hypermethylation of promoter CpG islands, enrichment of TP53, ARID1A, and BRCA1/2 DNA repair associated (BCRA1/2) mutations with reduced expression of Tet methylcytosine dioxygenase 1 (TET1), and enhanced expression of enhancer of zeste 2 polycomb repressive complex 2 subunit (EZH2) and Erb-B2 receptor tyrosine kinase 2 (ERBB2) amplification.
Cluster 2 was also enriched in TP53 mutations and ERBB2 amplification, CTNNB1, Wnt family member 5B (WNT5B), and AKT serine/threonine kinase 1 (AKT1). It consisted of both Fluke-positive and Fluke-negative CCAs and represented a low level of methylation.
Cluster 3 showed specific upregulation of immune system genes, including immune checkpoint genes (programmed death receptor 1 (PD-1), programmed cell death 1 ligand 2 (PD-L2), and B- and T-lymphocyte-associated protein (BTLA)) and pathways related to the costimulation of T lymphocytes. Similar to Cluster 2, it was also characterized by a low methylation level.
Cluster 4 was characterized by enrichment of BRCA1 associated protein 1 (BAP1) and isocitrate dehydrogenase (NADP(+)) 1/isocitrate dehydrogenase (NADP(+)) 2 (IDH1/2) mutations and fibroblast growth factor receptor (FGFR) aberrations with upregulated expression of FGFR1, FGFR2, FGFR3 and FGFR4. Similar to Cluster 1 its level of methylation was high, but, inversely, the methylation phenotype included CpG shore hypermethylation instead of CpG island hypermethylation, suggesting distinct mutational pathways.
Additionally, Clusters 1 and 2 were mostly represented by extrahepatic tumors, while Clusters 3 and 4 were characterized by intrahepatic malignancies almost entirely. Clusters 3 and 4 were associated with significantly better overall survival.
In another study, Lowery et al. [40] investigated molecular profiling of intrahepatic and extrahepatic CCA. An analysis carried out among CCA samples of 195 patients showed that in intrahepatic CCA the most commonly seen aberrations were : IDH1 (30%), ARID1A (23%), BAP1 (20%), TP53 (20%), and FGFR2 gene fusions (14%). In the case of extrahepatic CCA, the most commonly found aberrations were: KRAS proto-oncogene, GTPase (KRAS), SMAD family member 4 (SMAD4), and serine/threonine kinase 11 (STK11) alterations. In addition, CDKN2A/B and ERBB2 gene alterations were correlated with reduced overall survival and time to progression on first-line chemotherapy. Forty-seven percent of the patients showed somatic alterations with potential therapeutic value and thus 16% of the patients were enrolled in clinical trials molecular therapies.
Li et al. [41] carried out a study aimed at exploring the biological functions and prognostic biomarkers involved in CCA through transcriptional analysis. Thirty-three samples were obtained from CCA patients and 8 normal tissue samples. They discovered a total of 1463 differentially expressed genes, of which 267 were significantly upregulated and the remaining 1196 were significantly downregulated. According to Gene Ontology (GO) analysis, upregulated genes were enriched in ‘cadherin binding in cell-cell adhesion’, ‘extracellular matrix organization’ and ‘cell-cell adherens junctions’, while the downregulated ones were enriched in ‘oxidation-reduction process’, ‘extracellular exosomes’ and ‘blood microparticles’. Twenty-one of the genes were defied as hub genes including 8 upregulated genes and 13 downregulated genes. For all of the 21 hub genes, AUC was >0.900. Among upregulated hub genes, the expression level of CDK1, marker of proliferation Ki-67 (MKI67), DNA topoisomerase II alpha (TOP2A), and protein regulator of cytokinesis 1 (PRC1) were significantly negatively correlated to overall survival, while no similar correlation was found in remaining hub genes. Additionally, among downregulated hub genes, the expression of acyl-CoA oxidase 1 (ACOX1), apolipoprotein A2 (APOA2), apolipoprotein B (APOB), fibrinogen alpha chain (FGA), and fibrinogen gamma chain (FGG) were negatively correlated with the tumor stage of CCA patients. The authors suggest that CDK1, MKI67, TOP2A, and PRC1 could be used as prognostic biomarkers of CCA. The summary of detection of the biomarkers is showed in Figure 3.
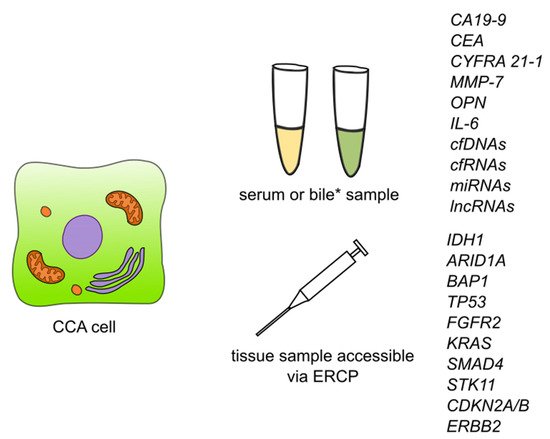
Figure 3. Summary of detection of biomarkers and some of the genetic markers for cholangiocarcinoma. Abbreviations: CA19-9—carbohydrate antigen 19-9/cancer antigen 19-9; CEA—carcinoembryonic antigen; CYFRA 21-1—cytokeratin fragment antigen 21-1; MMP-7—metalloproteinase 7; IL-6—interleukin 6; cfDNAs—circulating free DNAs; cfRNAs—circulating free RNAs; lncRNAs—long non-coding RNAs; ERCP—endoscopic retrograde cholangiopancreatography; IDH1—isocitrate dehydrogenase (NADP(+)) 1; ARID1A—AT-rich interaction domain 1A; BAP1—BRCA1 associated protein 1; TP53—tumor protein P53; FGFR2—fibroblast growth factor receptor 2; KRAS—KRAS proto-oncogene, GTPase; SMAD4—SMAD family member 4; STK11—serine/threonine kinase 11; CDKN2A/B—cyclin dependent kinase inhibitor 2A/cyclin dependent kinase inhibitor 2B; ERBB2—Erb-B2 receptor tyrosine kinase 2. * Detection of CCA biomarkers in bile has so far been described for CEA and cfDNAs.
References
- Singal, A.G.; Lampertico, P.; Nahon, P. Epidemiology and surveillance for hepatocellular carcinoma: New trends. J. Hepatol. 2020, 72, 250–261.
- Bertuccio, P.; Turati, F.; Carioli, G.; Rodriguez, T.; La Vecchia, C.; Malvezzi, M.; Negri, E. Global trends and predictions in hepatocellular carcinoma mortality. J. Hepatol. 2017, 67, 302–309.
- Kulik, L.; El-Serag, H.B. Epidemiology and Management of Hepatocellular Carcinoma. Gastroenterology 2019, 156, 477–491.
- De Stefano, F.; Chacon, E.; Turcios, L.; Marti, F.; Gedaly, R. Novel biomarkers in hepatocellular carcinoma. Dig. Liver Dis. 2018, 50, 1115–1123.
- Khan, A.S.; Dageforde, L.A. Cholangiocarcinoma. Surg. Clin. N. Am. 2019, 99, 315–335.
- Bertuccio, P.; Malvezzi, M.; Carioli, G.; Hashim, D.; Boffetta, P.; El-Serag, H.B.; La Vecchia, C.; Negri, E. Global trends in mortality from intrahepatic and extrahepatic cholangiocarcinoma. J. Hepatol. 2019, 71, 104–114.
- Bergquist, A.; Von Seth, E. Epidemiology of cholangiocarcinoma. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 221–232.
- Intuyod, K.; Armartmuntree, N.; Jusakul, A.; Sakonsinsiri, C.; Thanan, R.; Pinlaor, S. Current omics-based biomarkers for cholangiocarcinoma. Expert Rev. Mol. Diagn. 2019, 19, 997–1005.
- Macias, R.I.R.; Kornek, M.; Rodrigues, P.M.; Paiva, N.A.; Castro, R.E.; Urban, S.; Pereira, S.P.; Cadamuro, M.; Rupp, C.; Loosen, S.H.; et al. Diagnostic and prognostic biomarkers in cholangiocarcinoma. Liver Int. 2019, 39, 108–122.
- Shen, N.; Zhang, D.; Yin, L.; Qiu, Y.; Liu, J.; Yu, W.; Fu, X.; Zhu, B.; Xu, X.; Duan, A.; et al. Bile cell-free DNA as a novel and powerful liquid biopsy for detecting somatic variants in biliary tract cancer. Oncol. Rep. 2019, 42, 549–560.
- Liu, C.H.; Huang, Q.; Jin, Z.Y.; Xie, F.; Zhu, C.L.; Liu, Z.; Wang, C. Circulating microRNA-21 as a prognostic, biological marker in cholangiocarcinoma. J. Cancer Res. Ther. 2018, 14, 220–225.
- Lixin, S.; Wei, S.; Haibin, S.; Qingfu, L.; Tiemin, P. miR-885-5p inhibits proliferation and metastasis by targeting IGF2BP1 and GALNT3 in human intrahepatic cholangiocarcinoma. Mol. Carcinog. 2020, 59, 1371–1381.
- Hann, H.W.; Wang, M.; Hafner, J.; Long, R.E.; Kim, S.H.; Ahn, M.; Park, S.; Comunale, M.A.; Block, T.M.; Mehta, A. Analysis of GP73 in patients with HCC as a function of anti-cancer treatment. Cancer Biomarkers 2010, 7, 269–273.
- Shen, S.; Lin, Y.; Yuan, X.; Shen, L.; Chen, J.; Chen, L.; Qin, L.; Shen, B. Biomarker MicroRNAs for Diagnosis, Prognosis and Treatment of Hepatocellular Carcinoma: A Functional Survey and Comparison. Sci. Rep. 2016, 6, 1–21.
- Cui, Y.; Xu, H.F.; Liu, M.Y.; Xu, Y.J.; He, J.C.; Zhou, Y.; Cang, S.D. Mechanism of exosomal microRNA-224 in development of hepatocellular carcinoma and its diagnostic and prognostic value. World J. Gastroenterol. 2019, 25, 1890–1898.
- Yang, C.; Ma, X.; Guan, G.; Liu, H.; Yang, Y.; Niu, Q.; Wu, Z.; Jiang, Y.; Bian, C.; Zang, Y.; et al. MicroRNA-766 promotes cancer progression by targeting NR3C2 in hepatocellular carcinoma. FASEB J. 2019, 33, 1456–1467.
- Liu, Y.; Tan, J.; Ou, S.; Chen, J.; Chen, L. Adipose-derived exosomes deliver miR-23a/b to regulate tumor growth in hepatocellular cancer by targeting the VHL/HIF axis. J. Physiol. Biochem. 2019, 75, 391–401.
- Lin, H.; Huang, Z.P.; Liu, J.; Qiu, Y.; Tao, Y.P.; Wang, M.C.; Yao, H.; Hou, K.Z.; Gu, F.M.; Xu, X.F. MiR-494-3p promotes PI3K/AKT pathway hyperactivation and human hepatocellular carcinoma progression by targeting PTEN. Sci. Rep. 2018, 8, 1–9.
- Jiang, L.; Cheng, Q.; Zhang, B.H.; Zhang, M.Z. Circulating micrornas as biomarkers in hepatocellular carcinoma screening a validation set from China. J. Med. 2015, 94, 1–10.
- Schulze, K.; Imbeaud, S.; Letouzé, E.; Alexandrov, L.B.; Calderaro, J.; Rebouissou, S.; Couchy, G.; Meiller, C.; Shinde, J.; Soysouvanh, F.; et al. Exome sequencing of hepatocellular carcinomas identifies new mutational signatures and potential therapeutic targets. Nat. Genet. 2015, 47, 505–511.
- Guichard, C.; Amaddeo, G.; Imbeaud, S.; Ladeiro, Y.; Pelletier, L.; Maad, I.B.; Calderaro, J.; Bioulac-Sage, P.; Letexier, M.; Degos, F.; et al. Integrated analysis of somatic mutations and focal copy-number changes identifies key genes and pathways in hepatocellular carcinoma. Nat. Genet. 2012, 44, 694–698.
- Wang, X.W.; Hussain, S.P.; Huo, T.I.; Wu, C.G.; Forgues, M.; Hofseth, L.J.; Brechot, C.; Harris, C.C. Molecular pathogenesis of human hepatocellular carcinoma. Toxicology 2002, 181–182, 43–47.
- Long, J.; Wang, A.; Bai, Y.; Lin, J.; Yang, X.; Wang, D.; Yang, X.; Jiang, Y.; Zhao, H. Development and validation of a TP53-associated immune prognostic model for hepatocellular carcinoma. EBioMedicine 2019, 42, 363–374.
- Ozen, C.; Yildiz, G.; Dagcan, A.T.; Cevik, D.; Ors, A.; Keles, U.; Topel, H.; Ozturk, M. Genetics and epigenetics of liver cancer. N. Biotechnol. 2013, 30, 381–384.
- Abitbol, S.; Dahmani, R.; Coulouarn, C.; Ragazzon, B.; Mlecnik, B.; Senni, N.; Savall, M.; Bossard, P.; Sohier, P.; Drouet, V.; et al. AXIN deficiency in human and mouse hepatocytes induces hepatocellular carcinoma in the absence of β-catenin activation. J. Hepatol. 2018, 68, 1203–1213.
- He, F.; Li, J.; Xu, J.; Zhang, S.; Xu, Y.; Zhao, W.; Yin, Z.; Wang, X. Decreased expression of ARID1A associates with poor prognosis and promotes metastases of hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2015, 34.
- Yang, H.; Huo, J.; Li, X. Identification and validation of a five-gene prognostic signature for hepatocellular carcinoma. World J. Surg. Oncol. 2021, 19, 1–13.
- Nahon, P.; Nault, J.C. Constitutional and functional genetics of human alcohol-related hepatocellular carcinoma. Liver Int. 2017, 37, 1591–1601.
- Piao, Z.; Park, C.; Lee, J.S.; Yang, C.H.; Choi, K.Y.; Kim, H. Homozygous deletions of the CDKN2 gene and loss of heterozygosity of 9p in primary hepatocellular carcinoma. Cancer Lett. 1998, 122, 201–207.
- Rizzo, A.; Ricci, A.D.; Tavolari, S.; Brandi, G. Circulating tumor DNA in biliary tract cancer: Current evidence and future perspectives. Cancer Genom. Proteom. 2020, 17, 441–452.
- Wasenang, W.; Chaiyarit, P.; Proungvitaya, S.; Limpaiboon, T. Serum cell-free DNA methylation of OPCML and HOXD9 as a biomarker that may aid in differential diagnosis between cholangiocarcinoma and other biliary diseases. Clin. Epigenet. 2019, 11, 39.
- Driescher, C.; Fuchs, K.; Haeberle, L.; Goering, W.; Frohn, L.; Opitz, F.V.; Haeussinger, D.; Knoefel, W.T.; Keitel, V.; Esposito, I. Bile-based cell-free DNA analysis is a reliable diagnostic tool in pancreatobiliary cancer. Cancers 2021, 13, 39.
- Wang, X.; Fu, X.H.; Qian, Z.L.; Zhao, T.; Duan, A.Q.; Ruan, X.; Zhu, B.; Yin, L.; Zhang, Y.J.; Yu, W.L. Non-invasive detection of biliary tract cancer by low-coverage whole genome sequencing from plasma cell-free DNA: A prospective cohort study. Transl. Oncol. 2021, 14, 100908.
- Gu, X.; Wang, C.; Deng, H.; Qing, C.; Liu, R.; Liu, S.; Xue, X. Exosomal piRNA profiling revealed unique circulating piRNA signatures of cholangiocarcinoma and gallbladder carcinoma. Acta Biochim. Biophys. Sin. 2021, 52, 475–484.
- Bai, J.G.; Tang, R.F.; Shang, J.F.; Qi, S.; Yu, G.D.; Sun, C. Upregulation of long non-coding RNA CCAT2 indicates a poor prognosis and promotes proliferation and metastasis in intrahepatic cholangiocarcinoma. Mol. Med. Rep. 2018, 17, 5328–5335.
- Angenard, G.; Merdrignac, A.; Louis, C.; Edeline, J.; Coulouarn, C. Expression of long non-coding RNA ANRIL predicts a poor prognosis in intrahepatic cholangiocarcinoma. Dig. Liver Dis. 2019, 51, 1337–1343.
- Yao, Y.; Jiao, D.; Liu, Z.; Chen, J.; Zhou, X.; Li, Z.; Li, J.; Han, X. Novel miRNA Predicts Survival and Prognosis of Cholangiocarcinoma Based on RNA-seq Data and in Vitro Experiments. Biomed Res. Int. 2020, 2020, 1–14.
- Asukai, K.; Kawamoto, K.; Eguchi, H.; Konno, M.; Asai, A.; Iwagami, Y.; Yamada, D.; Asaoka, T.; Noda, T.; Wada, H.; et al. Micro-RNA-130a-3p Regulates Gemcitabine Resistance via PPARG in Cholangiocarcinoma. Ann. Surg. Oncol. 2017, 24, 2344–2352.
- Jusakul, A.; Cutcutache, I.; Yong, C.H.; Lim, J.Q.; Huang, M.N.; Padmanabhan, N.; Nellore, V.; Kongpetch, S.; Ng, A.W.T.; Ng, L.M.; et al. Whole-genome and epigenomic landscapes of etiologically distinct subtypes of cholangiocarcinoma. Cancer Discov. 2017, 7, 1116–1135.
- Lowery, M.A.; Ptashkin, R.; Jordan, E.; Berger, M.F.; Zehir, A.; Capanu, M.; Kemeny, N.E.; O’Reilly, E.M.; El-Dika, I.; Jarnagin, W.R.; et al. Comprehensive molecular profiling of intrahepatic and extrahepatic cholangiocarcinomas: Potential targets for intervention. Clin. Cancer Res. 2018, 24, 4154–4161.
- Li, H.; Long, J.; Xie, F.; Kang, K.; Shi, Y.; Xu, W.; Wu, X.; Lin, J.; Xu, H.; Du, S.; et al. Transcriptomic analysis and identification of prognostic biomarkers in cholangiocarcinoma. Oncol. Rep. 2019, 42, 1833–1842.
More