Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Beatrix Zheng and Version 1 by Lilian M. Mennink.
Accumulating evidence suggests that inflammation plays a role in the pathogenesis of subjective tinnitus. Noise exposure and salicylate administration both lead to inflammation throughout the whole auditory pathway. In particular, TNF-α, IL-1β, glia and activated platelets are associated with acute tinnitus. TNF-α and IL-1β influence NMDA and GABA receptors, leading to an increased excitatory and decreased inhibitory neurotransmission. These changes can lead to neuroplasticity and thus chronic tinnitus. Whether inflammatory mediators still play a role in chronic tinnitus remains to be elucidated. Nevertheless, drugs targeting the involved inflammatory mediators could be a potential effective treatment for (acute) tinnitus.
- tinnitus
- inflammation
- microglia
- astrocytes
- cytokines
- platelets
1. Introduction
Subjective tinnitus is the perception of sound without the presence of an external source. Chronic tinnitus is prevalent among 5% to 42% of the general population, of which 3% to 30% report their tinnitus to be bothersome [1]. Quality of life is negatively affected by tinnitus in 1–4% of the general population, making tinnitus a substantial medical and socioeconomic problem [2]. Current treatments for tinnitus, such as hearing aids, cognitive behavioral therapy, and sound therapy aim to reduce tinnitus perception or to develop new coping strategies. However, there is no treatment that targets the pathophysiologic mechanism.
Knowledge of the pathophysiology of tinnitus is essential for the development of new treatment strategies. Previous research has established that tinnitus is associated with aberrant neuronal firing, regardless of the cause of tinnitus [3,4][3][4]. However, the processes that contribute to the emergence of aberrant neuronal firing in tinnitus are still a matter of debate.
2. Current Insights
Experimental studies found an increase of TNF-α in the cochlea, the CN, the IC, and the AC, as well as an increase of IL-1β in the cochlea, the IC, the MGB, and the AC of animals with tinnitus (Figure 31). In addition, microglial markers were increased in the MGB and AC, but decreased in the CN. Astrocytic markers were elevated in the CN and AC. Only human CBC measurements could be meta-analyzed. MPV and PDW in blood samples were increased only if outliers/influential studies were removed. Together, these results indicate that inflammation may play a role in tinnitus.
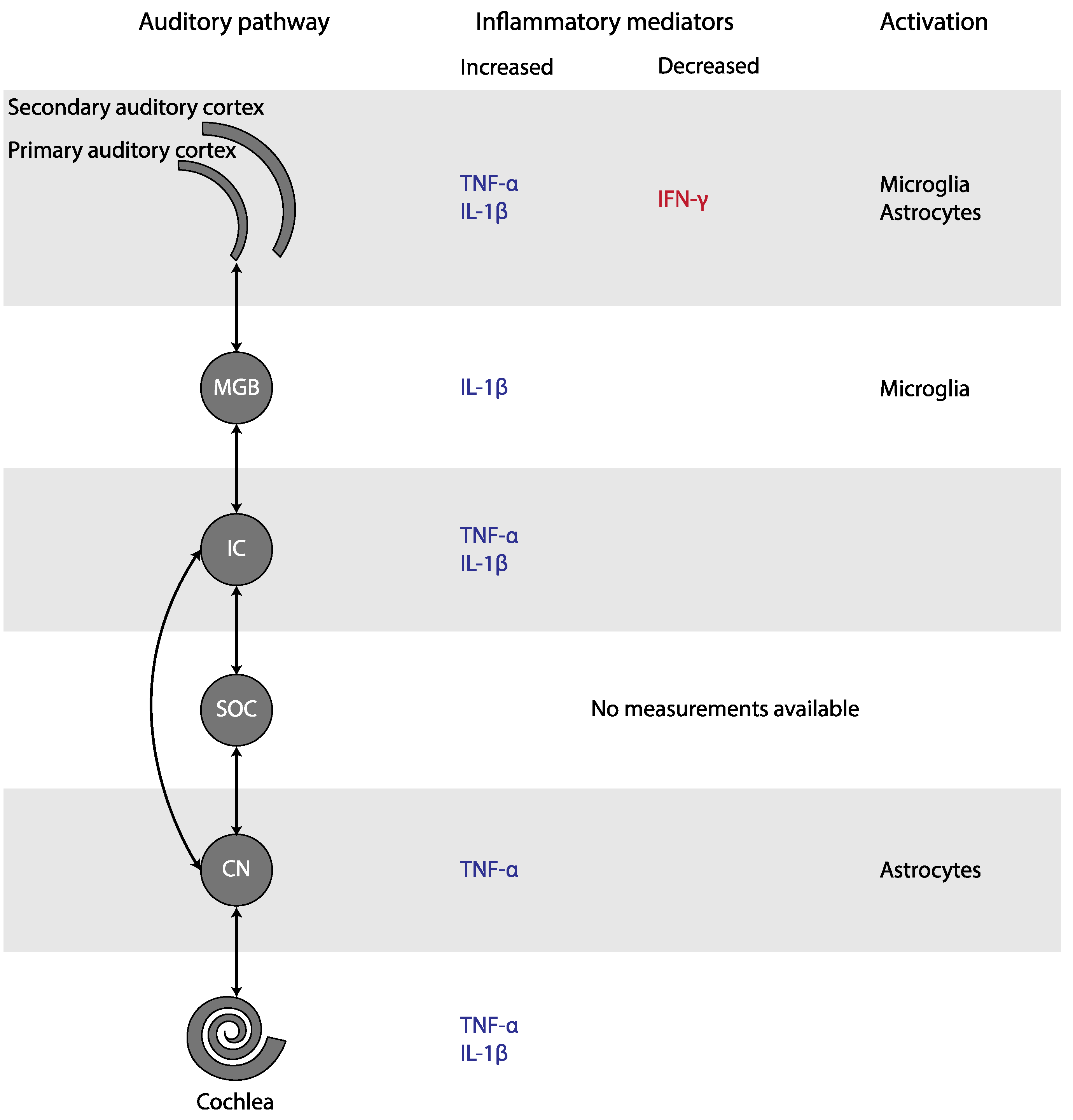
Figure 31. Changes in inflammatory mediators and glial activation in tinnitus, depicted in the simplified auditory pathway. Abbreviations: CN, cochlear nucleus; IC, inferior colliculus; IFN, interferon; IL, Interleukin; MGB, medial geniculate body; SOC, superior olivary complex; TNF, Tumor Necrosis Factor.
2.1. Cytokine Involvement in Tinnitus
2.1.1. TNF-α in Tinnitus
TNF-α is a potent pro-inflammatory molecule produced by brain-resident astrocytes, microglia, and neurons [45][7]. It appears to play a critical role in the pathogenesis of experimental tinnitus. TNF-α was consistently increased after tinnitus induction and infusion of TNF-α in the AC of healthy mice resulted in tinnitus [16,17,18,19,46][8][9][10][11][12]. Moreover, genetic deletion and pharmacological blockade of TNF-α prevented the occurrence of tinnitus and noise-induced microglial activation. After infusion of TNF-α, these animals developed tinnitus as well. Additionally, pharmacological depletion of microglia before noise exposure prevented a TNF-α increase and noise-induced tinnitus [16][8]. These results suggest that microglia are (for a large part) responsible for increased TNF-α expression, and that microglial activation and TNF-α depend on each other in tinnitus induction [16][8]. Once TNF-α is present, it is able to activate parenchymal microglia, forming a self-stimulating loop [47][13].
In contrast with the experimental studies, there was no increase in TNF-α present in humans with tinnitus. This discrepancy may be caused by the timing of measurements relative to the start of tinnitus. In animals, the increases in TNF-α were detected almost directly after tinnitus induction in most cases, while in humans, cytokine levels were measured in subjects with chronic tinnitus, i.e., the measurements took place much longer after the start of tinnitus. TNF-α levels may have been elevated initially and reduced to normal levels in the chronic state. Additionally, in animal models, cytokine levels were measured in brain tissue, whereas human cytokine levels were measured in blood. Last, damage because of the tinnitus induction methods in animals itself could be responsible for an increase in TNF-α, independent of the occurrence of tinnitus. However, animals that were treated the same but did not develop tinnitus also did not show an increase in TNF-α. Taken altogether, it is probable that noise exposure and salicylate administration cause an increase in TNF-α, which leads to acute tinnitus. In contrast, in chronic tinnitus TNF-α may not play a role anymore.
2.1.2. IL-1β in Tinnitus
IL-1β (gene) expression was consistently increased in experimental tinnitus. Similar to TNF-α, IL-1β was not increased in humans, although it was only evaluated in one study [31][14]. IL-1β is a potent pro-inflammatory cytokine that stimulates microglia and astrocytes, and stimulates the expression of inflammatory mediators, including itself [48][15]. It can be produced by microglia, astrocytes, endothelial cells, infiltrating leukocytes, neurons, and oligodendrocytes [49][16]. Since IL-1β is hardly detectable in healthy brains [48,50][15][17], the elevation in its (gene) expression levels in rats with tinnitus suggests that IL-1β is present in tinnitus. The positive association between tinnitus scores and IL-1β levels in mice seems to confirm this [20][18]. However, it is not clear whether the increased (gene) expression of IL-1β (partly) caused tinnitus or vice versa. For the difference between IL-1β expression between human and animal studies, the same reasons as mentioned for TNF-α could apply.
2.2. Neuroglial Involvement in Tinnitus
Microglial activation is characterized by the increased expression of specific markers, such as Iba-1, and morphological changes from the ramified shape to a non-ramified shape. The number of Iba-1 positive microglia and their activation state were increased in the MGB and in the primary AC, but decreased in the CN. After activation, microglia produce proinflammatory cytokines such as TNF-α. These cytokines further activate the microglia, forming a self-stimulating loop [16][8]. The central role of microglia is further supported by the observation that microglial depletion prevented an increase in TNF-α expression and noise-induced tinnitus [16][8]. Similar to microglial activation, astrocytic activation is characterized by increased expression of specific markers, such as GFAP, and deramification of their processes [51][19]. The number of GFAP-positive astrocytes and their activation state were increased in the ventral CN and primary AC, but not in the MGB. Astrocytes are important in regulating levels of neurotransmitter and ion concentrations, controlling synapse formation and function, and repairing the nervous system [52,53][20][21]. In response to neuronal degeneration, they promote synaptic regrowth and axonal sprouting [22,54][22][23]. This may lead to enhanced synaptic activity. Moreover, astrocytes can directly excite neighboring neurons through a calcium-dependent glutamate release and promote neural synchrony mediated by extra-synaptic excitatory receptors [55,56][24][25]. Furthermore, they can produce the proinflammatory cytokines implicated in tinnitus [57][26]. Thus, both microglia and astrocytes may play an important role in tinnitus.
2.3. Platelet Involvement in Tinnitus
In humans with tinnitus, MPV was increased in six out of ten studies, but this was only significant when outliers or influential cases were removed. PDW was increased in three out of four studies that examined this marker, but like MPV this was only significant when an influential study was removed. MPV indicates the size of platelets, whereas PDW reflects variation of platelet size distribution. Since platelets change their shape when activated, increased MPV and PDW can both be used as a sign of activated platelets. In line with these results, Chrbolka et al. (2020) showed that platelet activity was increased in patients with tinnitus [36][27]. Platelet count and NLR seem to remain unchanged.
Platelets may be involved in the development of tinnitus in several ways. Platelets release granule-stored cytokines such as IL-6, IL-8 and TNF-α within seconds to minutes after activation. In addition, they produce various chemokines and cytokines, such as IL-1β, within hours after activation [58,59][28][29]. These cytokines, and thus platelets, are potent inducers of the acute phase response [60][30]. Therefore, platelets are an important source of cytokines in the inflammatory response. On the other hand, increased platelet volumes may be an indicator of a prothrombic condition or even cause thrombosis, e.g., in the internal auditory artery leading to hypoperfusion of the cochlea, impairing its function and contributing to the development of tinnitus [28,38,42][31][32][33]. Finally, platelets are involved in glutamate uptake. Reduced systemic glutamate uptake by platelets, as well as glutamate excitotoxicity, has been demonstrated in various neurodegenerative disorders, such as Alzheimer’s disease, Parkinson’s disease, and amyotrophic lateral sclerosis [61][34]. Although speculative, reduced systemic glutamate uptake by platelets could play a role in tinnitus as well, given the elevated glutamate concentration in tinnitus [62][35].
2.4. Inflammation in the Pathophysiology of Tinnitus
The question remains how inflammation would be involved in the pathophysiology of tinnitus. Tinnitus has been associated with an increase in excitatory and a decrease in inhibitory neurotransmission [62][35]. Inflammation, on the other hand, may lead to alterations in synaptic transmission and synaptic organization.
Salicylate-induced tinnitus is associated with the increased activity of the NMDA (N-methyl-D-aspartate) receptor, one of the excitatory glutamate receptors [63][36] (Figure 42). Some animal studies included here also studied the (gene) expression of NR2A and NR2B, two NMDA receptor subunits. Both NR2A and NR2B (gene) expression were consistently elevated in the IC, the AC, and the cochlea in animals with tinnitus after salicylate injection. Moreover, a positive association between NR2B and tinnitus scores was present [17,18,19,20,23,46][9][10][11][18][37][12]. When tinnitus was diminished, (gene) expression of both NR2A and NR2B returned to normal as well. Thus, in animals with salicylate-induced tinnitus, NMDA (gene) expression is increased. Interestingly, this increase was consistently coexistent with increases in TNF-α and IL-1β in animals with tinnitus throughout the entire auditory pathway [17,18,19,46][9][10][11][12]. The specific effect of TNF-α on the NMDA receptor is still a matter of debate as both an increase and decrease of NMDA receptor currents have been demonstrated [64,65,66,67][38][39][40][41]. However, most studies show an increase. Additionally, TNF-α enhances surface localization of NMDA receptor subunits [67][41]. Last, Wang et al. (2019) showed that tinnitus was associated with TNF-α-dependent increased excitatory and decreased inhibitory synaptic currents [16][8]. Besides TNF-α, IL-1β has also been shown to enhance NMDA-induced currents [66,68][40][42]. In sum, the increased expression of TNF-α, and possibly also IL-1β, leads to increased NMDA receptor-dependent calcium influx and enhanced post-synaptic currents. This leads to increased neural activity.
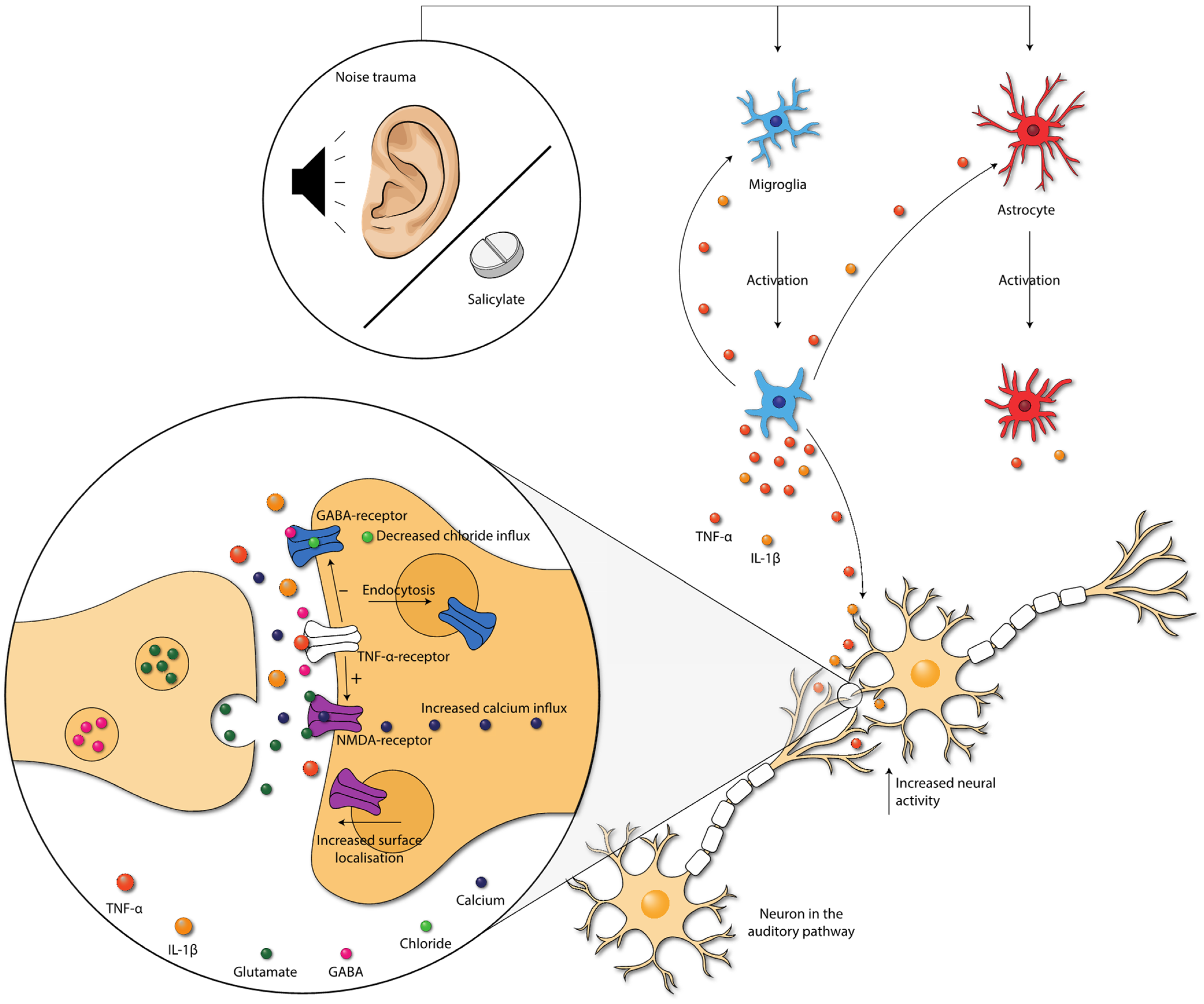
Figure 42. Damage caused by noise exposure and salicylate administration leads to the activation of neuroglia. Neuroglia produce cytokines that further activate the neuroglia. Moreover, cytokines influence neurotransmission within the auditory pathway. TNF-α increases the NMDA-receptor and decreases the GABAA-receptor activity, via the TNF-α-receptor. Moreover, NMDA receptor surface localization increases, and GABA-receptor surface localization decreases. These changes lead to increased neural activity in the auditory pathway. Abbreviations: GABA, gamma-aminobutyric acid; IL, interleukin; NMDA, N-methyl-D-aspartate; TNF, tumor necrosis factor.
The gamma-aminobutyric acid (GABA)-receptor is one of the major inhibitory neurotransmitter receptors. A potent tinnitus inducer, sodium salicylate, has been shown to suppress GABA(A)-induced currents, and tinnitus was completely eliminated after oral administration of the GABA agonist Vigabatrin, which suggests the involvement of decreased inhibitory neurotransmission in tinnitus [69,70,71][43][44][45]. Interestingly, inhibitory neurotransmission is also impacted by cytokines. TNF-α expression influences inhibitory GABAA (gamma-aminobutyric acid) receptors by causing an endocytosis of GABAA receptors, resulting in fewer surface GABAA receptors and a decrease in inhibitory synaptic strength [72][46]. Additionally, IL-1β has been shown to suppress GABA-induced currents [66][40]. So, besides increasing excitatory neurotransmission, TNF-α and IL-1β presumably also suppress inhibitory neurotransmission in tinnitus.
Vice versa, the abnormal release and/or uptake of neurotransmitters can also result in inflammation [57][26]. Intraperitoneal injections of memantine, a non-competitive NMDA receptor antagonist, for seven consecutive days was successful in decreasing NR2B expression and attenuating tinnitus. It also decreased expression of TNF-α [46][12]. Therefore, it seems probable that salicylate and noise exposure cause both an imbalance in neurotransmission and neuroinflammation, leading to tinnitus. This could lead to a vicious circle in which inflammation and neurotransmitter imbalance reinforce each other, exacerbating tinnitus.
Changes in the activity of receptors can lead to synaptic plasticity, which is the ability of synapses to strengthen or weaken over time [73][47]. Therefore, the cytokine-induced changes in excitatory and inhibitory neurotransmission could influence neural plasticity associated with tinnitus [74][48]. Moreover, it has been stated that the proinflammatory cytokines as TNF-α and IL-1β itself may also directly alter long-term synaptic plasticity [66,75,76,77][40][49][50][51]. Synaptic plasticity may occur throughout the whole auditory pathway, both ascending and descending. The detailed, specific mechanisms and types of neuroplasticity in tinnitus are beyond the scope of this entreviewy. Therefore, wethe researchers refer to the reviews of Roberts (2018) and Wu et al. (2016) on this topic [78,79][52][53]. Concluding, synaptic plasticity caused by inflammation could explain the long-term effects after the induction of tinnitus.
Various studies have reported that psychological stress increases the bothersomeness and loudness of tinnitus [80][54]. Baigi et al. (2011) even suggested that stress is a key factor in modulating the severity of tinnitus [81][55]. Acute stress stimulates the inflammatory response by increasing the amount of circulating IL-6, IL-1β, TNF-α and IL-10 [82][56]. On the other hand, chronic stress is also associated with increases in the amount of circulating cytokines, such as IL-6 and TNF-α [82][56]. Therefore, it is not implausible that the effect of stress on the severity of tinnitus is effectuated via inflammatory pathways.
In conclusion, damage caused by noise exposure and salicylate administration leads to inflammation. In turn, proinflammatory cytokines seem to influence synaptic transmission by either increasing excitatory synaptic transmission and/or decreasing inhibitory synaptic transmission throughout the whole auditory tract. Neuroplasticity occurs because of the altered activity of these receptors. This could explain the long-term effects of tinnitus.
2.5. Challenges in the Interpretation of Results on Inflammation in Tinnitus
2.5.1. Translation from Animal to Human
When both animal studies and human studies are compared, translatability may become an issue. Whereas the included animal studies studied the effects of tinnitus in the acute phase, the human studies investigated effects of the chronic phase. Moreover, in most animal studies, tinnitus was induced by salicylate. However, in humans, most cases of chronic tinnitus are caused by noise exposure or are related to age-related hearing loss [2]. For salicylate to cause tinnitus in humans, high doses are required [83][57]. Even though these are valid concerns, it does not mean that animal studies are not of value for unraveling the pathophysiology of tinnitus in humans. A recent study showed that, although different methods are employed to study the activity and connectivity of brain areas, there is consistency within the results between these approaches in human and animal studies [84][58]. Therefore, the inflammatory effects shown in animal models for tinnitus may very well play a role in humans as well.
2.5.2. Contribution of Hearing Loss
Another difficulty in the interpretation of results on inflammation in tinnitus is the potential contribution of hearing loss. As stated before, the pathophysiology of tinnitus and that of acquired hearing loss are closely related to such an extent that they often co-exist. Moreover, inflammation has been implicated in the pathophysiology of hearing loss as well [5,6][5][6]. Therefore, it is a challenge to disentangle both pathophysiological mechanisms.
Animal studies included here showed that both salicylate administration and noise trauma not only induce tinnitus, but also affect hearing thresholds [16,22,24,46,85][8][22][59][12][60]. In the study of Wang et al. (2019), TNF-α knockout mice had a similar auditory brainstem response as wild type mice after noise exposure, but the TNF-α knockout mice did not develop tinnitus, whereas wild type mice did [16][8]. This proves that TNF-α is essential for the development of tinnitus, but not for hearing loss. Therefore, elevated levels of TNF-α can (also) be attributed to the presence of tinnitus in animals.
In many human studies, hearing thresholds were not reported, and patients with hearing loss were not excluded. Only Kemal et al. (2016), Yildiz et al. (2020), Bayraktar and Taşolar (2017) and Chrbolka et al. (2020) excluded patients with a pure tone average over 20dB, 25dB and 40dB, respectively [25,28,36,43][61][31][27][62]. Moreover, Ceçen et al. (2021), Ozbay et al. (2015) and Savastano et al. (2006) excluded patients with (moderate or severe) hearing loss, but did not provide cut-off values [30,35,41][63][64][65]. Studies comparing human cytokine levels between patients with and without tinnitus did also not account for hearing loss in their analysis [31,32][14][66]. Blood cytokine levels in humans with tinnitus may not be altered (except for IL-6 in one study), but a potential effect of tinnitus on the presence of cytokines may also be masked by the simultaneous presence of hearing loss. To conclude, patients included in the human studies are very heterogenic, and it cannot be ruled out that the presence of hearing loss affects the results.
2.6. Potential Treatment Options
Given the role of TNF-α, IL-1β and microglia in the tinnitus pathophysiology, these mediators could potentially be a target for treatment. Whereas inflammation seems to play a role in the acute phase of tinnitus, it is less clear in the chronic phase. Therefore, inflammatory targets for treatment may be mainly beneficial in acute tinnitus.
2.6.1. Treatments Targeting Cytokines
Commonly used TNF-α inhibitors include adalimumab, certolizumab pegol, etanercept, golimumab, and infliximab. With regard to tinnitus, only etanercept has been studied. Hwang et al. (2017) showed that intraperitonially injected Etanercept significantly reduced behavioral evidence of tinnitus in mice [24][59]. Alongside gene expression of TNFR1, TNFR2 and NR2B was reduced. Etanercept has been shown to be effective in patients with Alzheimer’s disease, in which it was suggested that it reversed neuronal excitability associated with TNF-α exposure by acting as a decoy receptor [86][67]. Currently, the first clinical trial is planned to study the effect of etanercept in humans with blast-induced tinnitus (ClinicalTrials.gov, identifier: NCT04066348).
Another drug targeting TNF-α is dithiothalidomide (dTT). DTT is an immunosuppressive used to treat multiple myeloma and erythema nodosum leprosum, among other afflictions. In mice, dTT injections for five days within an hour after noise exposure prevented the increases in TNF-α, IL-1β, IL-18, NLRP3 and microglial morphological changes in the primary AC. Moreover, the mice showed no behavioral evidence of tinnitus [16][8]. In rats, dTT did not prevent the onset of tinnitus when administered directly after noise exposure, but it did alleviate subsequent tinnitus [16][8]. These studies indicate that TNF-α-inhibitors may be helpful in treating tinnitus or preventing its onset.
Anakinra is a competitive IL-1-type-1-receptor antagonist which inhibits the binding of IL-1α and IL-1β. It is mainly used to treat rheumatoid arthritis. Canakinumab is a human monoclonal antibody targeting IL-1β which is used in the treatment of systemic juvenile idiopathic arthritis and active Still’s disease. However, no study on the effect of IL-1β antagonists on tinnitus has been performed yet, either in animals or in humans.
2.6.2. Treatments Targeting Microglia
Prevention of microglial activation may be a potential treatment strategy. Intrathecal administration of Minocycline, a broad-spectrum tetracycline antibiotic, inhibits the expression of pro-inflammatory cytokines by microglia, leading to reduced attenuated mRNA expression and decreased cerebrospinal fluid levels of IL-1β and TNF-α [87][68]. However, it has not been studied with regard to tinnitus.
References
- McCormack, A.; Edmondson-Jones, M.; Somerset, S.; Hall, D. A Systematic Review of the Reporting of Tinnitus Prevalence and Severity. Hear. Res. 2016, 337, 70–79.
- Eggermont, J.J.; Roberts, L.E. The Neuroscience of Tinnitus. Trends Neurosci. 2004, 27, 676–682.
- Baguley, D.; McFerran, D.; Hall, D. Tinnitus. Lancet 2013, 382, 1600–1607.
- Knipper, M.; Van Dijk, P.; Nunes, I.; Rüttiger, L.; Zimmermann, U. Advances in the Neurobiology of Hearing Disorders: Recent Developments Regarding the Basis of Tinnitus and Hyperacusis. Prog. Neurobiol. 2013, 111, 17–33.
- Fuentes-Santamaría, V.; Alvarado, J.C.; Melgar-Rojas, P.; Gabaldón-Ull, M.C.; Miller, J.M.; Juiz, J.M. The Role of Glia in the Peripheral and Central Auditory System Following Noise Overexposure: Contribution of TNF-α and IL-1β to the Pathogenesis of Hearing Loss. Front. Neuroanat. 2017, 11, 9.
- Frye, M.D.; Ryan, A.F.; Kurabi, A. Inflammation Associated with Noise-Induced Hearing Loss. J. Acoust. Soc. Am. 2019, 146, 4020–4032.
- Pan, W.; Zadina, J.E.; Harlan, R.E.; Weber, J.T.; Banks, W.A.; Kastin, A.J. Tumor Necrosis Factor-α: A Neuromodulator in the CNS. Neurosci. Biobehav. Rev. 1997, 21, 603–613.
- Wang, W.; Zhang, L.S.; Zinsmaier, A.K.; Patterson, G.; Leptich, E.J.; Shoemaker, S.L.; Yatskievych, T.A.; Gibboni, R.; Pace, E.; Luo, H.; et al. Neuroinflammation Mediates Noise-Induced Synaptic Imbalance and Tinnitus in Rodent Models. PLoS Biol. 2019, 17, e3000307.
- Hu, S.S.; Mei, L.; Chen, J.Y.; Huang, Z.W.; Wu, H. Effects of Salicylate on the Inflammatory Genes Expression and Synaptic Ultrastructure in the Cochlear Nucleus of Rats. Inflammation 2014, 37, 365–373.
- Chen, X.H.; Zheng, L.L. Expression of Pro-Inflammatory Cytokines in the Auditory Cortex of Rats with Salicylate-Induced Tinnitus. Mol. Med. Rep. 2017, 16, 5643–5648.
- Hwang, J.-H.; Chen, J.-C.; Chan, Y.-C. Effects of C-Phycocyanin and Spirulina on Salicylate-Induced Tinnitus, Expression of NMDA Receptor and Inflammatory Genes. PLoS ONE 2013, 8, e58215.
- Jang, C.H.; Lee, S.; Park, I.Y.; Song, A.; Moon, C.; Cho, G.-W. Memantine Attenuates Salicylate-Induced Tinnitus Possibly by Reducing NR2B Expression in Auditory Cortex of Rat. Exp. Neurobiol. 2019, 28, 495–503.
- Nguyen, M.D.; Julien, J.P.; Rivest, S. Innate Immunity: The Missing Link in Neuroprotection and Neurodegeneration? Nat. Rev. Neurosci. 2002, 3, 216–227.
- Haider, H.F.; Ribeiro, S.F.; Martins, C.; Ribeiro, D.; Trigueiros, N.; Szczepek, A.J.; Caria, H.; Hoare, D.J.; Paço, J.; Borrego, L.M. Tinnitus, Hearing Loss and Inflammatory Processes in an Older Portuguese Population. Int. J. Audiol. 2020, 59, 323–332.
- Shaftel, S.S.; Griffin, W.S.T.; Kerry, K.M. The Role of Interleukin-1 in Neuroinflammation and Alzheimer Disease: An Evolving Perspective. J. Neuroinflamm. 2008, 5, 7.
- Eriksson, C.; Van Dam, A.M.; Lucassen, P.J.; Bol, J.G.J.M.; Winblad, B.; Schultzberg, M. Immunohistochemical Localization of Interleukin-1β, Interleukin-1 Receptor Antagonist and Interleukin-1β Converting Enzyme/Caspase-1 in the Rat Brain after Peripheral Administration of Kainic Acid. Neuroscience 1999, 93, 915–930.
- Dinarello, C.A.; Thompson, R.C. Blocking IL-1: Interleukin 1 Receptor Antagonist in Vivo and in Vitro. Immunol. Today 1991, 12, 404–410.
- Hwang, J.-H.; Chen, J.-C.; Yang, S.-Y.; Wang, M.-F.; Chan, Y.-C. Expression of Tumor Necrosis Factor-(Alpha) and Interleukin-1(Beta) Genes in the Cochlea and Inferior Colliculus in Salicylate-Induced Tinnitus. J. Neuroinflamm. 2011, 8, 2–7.
- Pekny, M.; Wilhelmsson, U.; Tatlisumak, T.; Pekna, M. Astrocyte Activation and Reactive Gliosis—A New Target in Stroke? Neurosci. Lett. 2019, 689, 45–55.
- Zeng, Z.; Roussakis, A.A.; Lao-Kaim, N.P.; Piccini, P. Astrocytes in Parkinson’s Disease: From Preclinical Assays to in Vivo Imaging and Therapeutic Probes. Neurobiol. Aging 2020, 95, 264–270.
- Kane, C.J.M.; Drew, P.D. Neuroinflammatory Contribution of Microglia and Astrocytes in Fetal Alcohol Spectrum Disorders. J. Neurosci. Res. 2020, 99, 1973–1985.
- Fang, L.; Fu, Y.; Zhang, T.-Y. Salicylate-Induced Hearing Loss Trigger Structural Synaptic Modifications in the Ventral Cochlear Nucleus of Rats via Medial Olivocochlear (MOC) Feedback Circuit. Neurochem. Res. 2016, 41, 1343–1353.
- Smith, L.; Gross, J.; Morest, D.K. Fibroblast Growth Factors (FGFs) in the Cochlear Nucleus of the Adult Mouse Following Acoustic Overstimulation. Hear. Res. 2002, 169, 1–12.
- Parpura, V.; Basarsky, T.A.; Liu, F.; Jeftinija, K.; Jeftinija, S.; Haydon, P.G. Glutamate-Mediated Astrocyte-Neuron Signalling. Nature 1994, 369, 744–747.
- Fellin, T.; Pascual, O.; Gobbo, S.; Pozzan, T.; Haydon, P.G.; Carmignoto, G. Neuronal Synchrony Mediated by Astrocytic Glutamate through Activation of Extrasynaptic NMDA Receptors. Neuron 2004, 43, 729–743.
- Park, K.M.; Bowers, W.J. Tumor Necrosis Factor-Alpha Mediated Signaling in Neuronal Homeostasis and Dysfunction. Cell Signal. 2010, 22, 977–983.
- Chrbolka, P.; Alušík, Š.; Kalátová, D.; Paluch, Z. Increased Platelet Activity in Tinnitus Patients. Neuroendocrinol. Lett. 2020, 41, 102–106.
- Chen, Y.; Zhong, H.; Zhao, Y.; Luo, X.; Gao, W. Role of Platelet Biomarkers in Inflammatory Response. Biomark. Res. 2020, 8, 2–8.
- Morrell, C.N.; Aggrey, A.A.; Chapman, L.M.; Modjeski, K.L. Emerging Roles for Platelets as Immune and Inflammatory Cells. Blood 2014, 123, 2759–2767.
- Aggrey, A.A.; Srivastava, K.; Ture, S.; Field, D.J.; Morrell, C.N. Platelet Induction of the Acute-Phase Response Is Protective in Murine Experimental Cerebral Malaria. J. Immunol. 2013, 190, 4685–4691.
- Kemal, O.; Müderris, T.; Başar, F.; Kutlar, G.; Gül, F. Prognostic Value of Mean Platelet Volume on Tinnitus. J. Laryngol. Otol. 2016, 130, 162–165.
- Düzenli, U.; Bozan, N.; Aslan, M.; Özkan, H.; Turan, M.; Kıroğlu, A.F. A Retrospective Analysis of Haemotologic Parameters in Patients with Bilateral Tinnitus. East. J. Med. 2018, 23, 264–268.
- Ulusoy, B.; Bozdemir, K.; Akyol, M.; Mişe, H.I.; Kutluhan, A.; Korkmaz, M.H. Investigation of Neutrophil-to-Lymphocyte Ratio, Platelet-to-Lymphocyte Ratio and Mean Platelet Volume in Patients with Tinnitus. J. Laryngol. Otol. 2018, 132, 129–132.
- Behari, M.; Shrivastava, M. Role of Platelets in Neurodegenerative Diseases: A Universal Pathophysiology. Int. J. Neurosci. 2013, 123, 287–299.
- Eggermont, J.J. Tinnitus: Neurobiological Substrates. Drug Discov. Today 2005, 10, 1283–1290.
- Guitton, M.J.; Caston, J.; Ruel, J.; Johnson, R.M.; Pujol, R.; Puel, J.L. Salicylate Induces Tinnitus through Activation of Cochlear NMDA Receptors. J. Neurosci. 2003, 23, 3944–3952.
- Hwang, J.-H.; Chen, J.-C.; Yang, S.-Y.; Wang, M.-F.; Liu, T.-C.; Chan, Y.-C. Expression of COX-2 and NMDA Receptor Genes at the Cochlea and Midbrain in Salicylate-Induced Tinnitus. Laryngoscope 2011, 121, 361–364.
- Furukawa, K.; Mattson, M.P. The Transcription Factor NF-ΚB Mediates Increases in Calcium Currents and Decreases in NMDA- and AMPA/Kainate-Induced Currents Induced by Tumor Necrosis Factor-α in Hippocampal Neurons. J. Neurochem. 1998, 70, 1876–1886.
- Jara, J.H.; Singh, B.B.; Floden, A.M.; Combs, C.K. Tumor Necrosis Factor Alpha Stimulates NMDA Receptor Activity in Mouse Cortical Neurons Resulting in ERK-Dependent Death. J. Neurochem. 2007, 100, 1407–1420.
- Kawasaki, Y.; Zhang, L.; Cheng, J.K.; Ji, R.R. Cytokine Mechanisms of Central Sensitization: Distinct and Overlapping Role of Interleukin-1β, Interleukin-6, and Tumor Necrosis Factor-α in Regulating Synaptic and Neuronal Activity in the Superficial Spinal Cord. J. Neurosci. 2008, 28, 5189–5194.
- Wheeler, D.; Knapp, E.; Bandaru, V.V.R.; Wang, Y.; Knorr, D.; Poirier, C.; Mattson, M.P.; Geiger, J.D.; Haughey, N.J. Tumor Necrosis Factor-α-Induced Neutral Sphingomyelinase-2 Modulates Synaptic Plasticity by Controlling the Membrane Insertion of NMDA Receptors. J. Neurochem. 2009, 109, 1237–1249.
- Liu, T.; Jiang, C.Y.; Fujita, T.; Luo, S.W.; Kumamoto, E. Enhancement by Interleukin-1β of AMPA and NMDA Receptor-Mediated Currents in Adult Rat Spinal Superficial Dorsal Horn Neurons. Mol. Pain 2013, 9, 1.
- Wang, H.T.; Luo, B.; Zhou, K.Q.; Xu, T.L.; Chen, L. Sodium Salicylate Reduces Inhibitory Postsynaptic Currents in Neurons of Rat Auditory Cortex. Hear. Res. 2006, 215, 77–83.
- Jin, Y.; Luo, B.; Su, Y.Y.; Wang, X.X.; Chen, L.; Wang, M.; Wang, W.W.; Chen, L. Sodium Salicylate Suppresses GABAergic Inhibitory Activity in Neurons of Rodent Dorsal Raphe Nucleus. PLoS ONE 2015, 10, e0126956.
- Brozoski, T.J.; Spires, T.J.D.; Bauer, C.A. Vigabatrin, a GABA Transaminase Inhibitor, Reversibly Eliminates Tinnitus in an Animal Model. JARO J. Assoc. Res. Otolaryngol. 2007, 8, 105–118.
- Stellwagen, D.; Beattie, E.C.; Seo, J.Y.; Malenka, R.C. Differential Regulation of AMPA Receptor and GABA Receptor Trafficking by Tumor Necrosis Factor-α. J. Neurosci. 2005, 25, 3219–3228.
- Gerrow, K.; Triller, A. Synaptic Stability and Plasticity in a Floating World. Curr. Opin. Neurobiol. 2010, 20, 631–639.
- Shore, S.E.; Roberts, L.E.; Langguth, B. Maladaptive Plasticity in Tinnitus-Triggers, Mechanisms and Treatment. Nat. Rev. Neurol. 2016, 12, 150–160.
- Rizzo, F.R.; Musella, A.; De Vito, F.; Fresegna, D.; Bullitta, S.; Vanni, V.; Guadalupi, L.; Stampanoni Bassi, M.; Buttari, F.; Mandolesi, G.; et al. Tumor Necrosis Factor and Interleukin-1β Modulate Synaptic Plasticity during Neuroinflammation. Neural Plast. 2018, 2018, 8430123.
- Sakamoto, K.; Karelina, K.; Obrietan, K. CREB: A Multifaceted Regulator of Neuronal Plasticity and Protection. J. Neurochem. 2011, 116, 1–9.
- Vezzani, A.; Viviani, B. Neuromodulatory Properties of Inflammatory Cytokines and Their Impact on Neuronal Excitability. Neuropharmacology 2015, 96, 70–82.
- Roberts, L.E. Neural Plasticity and Its Initiating Conditions in Tinnitus. HNO 2018, 66, 172–178.
- Wu, C.; Stefanescu, R.A.; Martel, D.T.; Shore, S.E. Tinnitus: Maladaptive Auditory-Somatosensory Plasticity. Hear. Res. 2016, 334, 20–29.
- Elarbed, A.; Fackrell, K.; Baguley, D.M.; Hoare, D.J. Tinnitus and Stress in Adults: A Scoping Review. Int. J. Audiol. 2021, 60, 171–182.
- Baigi, A.; Oden, A.; Almlid-Larsen, V.; Barrenäs, M.L.; Holgers, K.M. Tinnitus in the General Population with a Focus on Noise and Stress: A Public Health Study. Ear Hear. 2011, 32, 787–789.
- Rohleder, N. Stress and Inflammation—The Need to Address the Gap in the Transition between Acute and Chronic Stress Effects. Psychoneuroendocrinology 2019, 105, 164–171.
- Stolzberg, D.; Salvi, R.J.; Allman, B.L. Salicylate Toxicity Model of Tinnitus. Front. Syst. Neurosci. 2012, 6, 28.
- Koops, E.A.; Eggermont, J.J. The Thalamus and Tinnitus: Bridging the Gap between Animal Data and Findings in Humans. Hear. Res. 2021, 407, 108280.
- Hwang, J.-H.; Huang, D.C.-W.; Lu, Y.-C.; Yang, W.-S.; Liu, T.-C. Effects of Tumor Necrosis Factor Blocker on Salicylate-Induced Tinnitus in Mice. Int. Tinnitus J. 2017, 21, 24–29.
- Manohar, S.; Dahar, K.; Adler, H.J.; Dalian, D.; Salvi, R. Noise-Induced Hearing Loss: Neuropathic Pain via Ntrk1 Signaling. Mol. Cell. Neurosci. 2016, 75, 101–112.
- Bayraktar, C.; Taşolar, S. Relationship between Increased Carotid Artery Stiffness and Idiopathic Subjective Tinnitus. Eur. Arch. Oto-Rhino-Laryngol. 2017, 274, 2125–2130.
- Yildiz, S.; Karaca, H.; Toros, S.Z. Mean Platelet Volume and Neutrophil to Lymphocyte Ratio in Patients with Tinnitus: A Case-Control Study. Braz. J. Otorhinolaryngol. 2020.
- Savastano, M.; Celadin, M.; Pittoni, M.; Plebani, M.; Marioni, G. Western Blot Immunoassay for HSP-70 Antibodies in Idiopathic Tinnitus: A Preliminary Report. Ann. Otol. Rhinol. Laryngol. 2006, 115, 243–246.
- Çeçen, A.; Kemal, Ö.; Yildirim, U.; Kavaz, E.; Terzi, Ö. The Clinical and Prognostic Value of the Neutrophil Lymphocyte Ratio, the Platelet Lymphocyte Ratio and Mean Platelet Volume in Tinnitus Patients. J. Exp. Clin. Med. 2021, 38, 251–254.
- Ozbay, I.; Kahraman, C.; Balikci, H.H.; Kucur, C.; Kahraman, N.K.; Ozkaya, D.P.; Oghan, F. Neutrophil-to-Lymphocyte Ratio in Patients with Severe Tinnitus: Prospective, Controlled Clinical Study. J. Laryngol. Otol. 2015, 129, 544–547.
- Weber, C.; Arck, P.; Mazurek, B.; Klapp, B.F. Impact of a Relaxation Training on Psychometric and Immunologic Parameters in Tinnitus Sufferers. J. Psychosom. Res. 2002, 52, 29–33.
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of Pro-Inflammatory Cytokines Released from Microglia in Neurodegenerative Diseases. Brain Res. Bull. 2012, 87, 10–20.
- Ledeboer, A.; Sloane, E.M.; Milligan, E.D.; Frank, M.G.; Mahony, J.H.; Maier, S.F.; Watkins, L.R. Minocycline Attenuates Mechanical Allodynia and Proinflammatory Cytokine Expression in Rat Models of Pain Facilitation. Pain 2005, 115, 71–83.
More