The term microbiota refers to the entire population of microorganisms that colonizes a particular location and includes not just bacteria, but also other microbes such as fungi, archaea, viruses, and protozoans. Cows have bacteria inhabiting the uterus even before calving and establish a unique endometrial microbiome within 20 min of calving where the microbiome is similar between cows that develop metritis and cows without endometritis until at least the second day postpartum. The microbiome of cows that develop metritis has higher relative abundance of Bacteroidetes and Fusobacteria and lower relative abundance of Proteobacteria and Tenericutes.
- genital
- microbiome
- health
- cattle
- pathogens
1. The Genital Tract Contains a Dynamic Microbiota
2. Significance of the Genital Microbiome
3. Origin of the Genital Microbiome
4. Diversity of the Genital Microbiome
4.1. Vagina
4.2. Uterus
5. Factors Affecting Genital Microbiome Diversity
5.1. Intrinsic Factors
5.1.1. Species
5.1.2. Breed (Genetic Background)
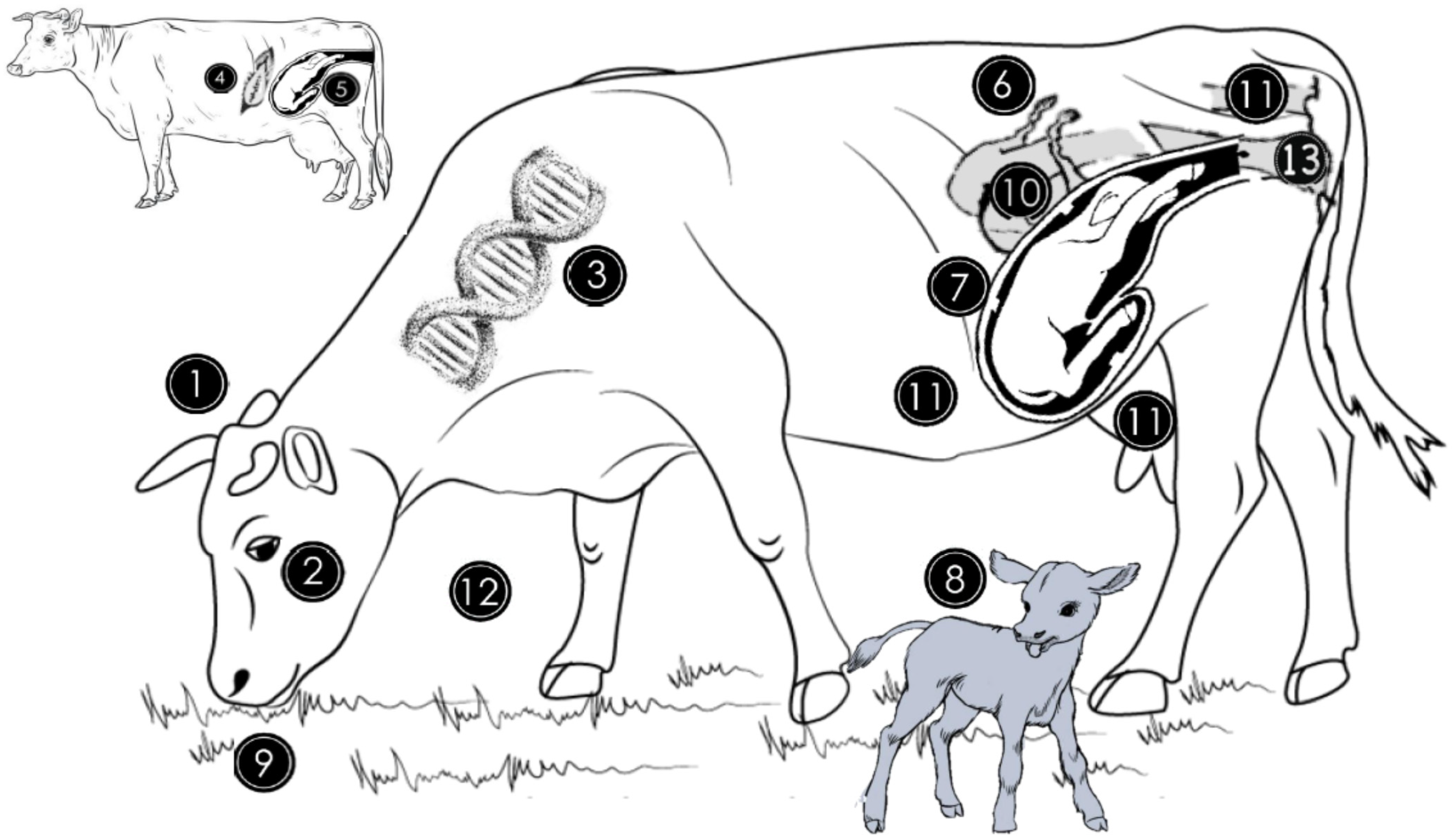
Figure 1. Origin of genital microbiome and factors that can affect the abundance and diversity of the microbial population. The genital microbiome is highly variable between species (1) and individuals of the same species (2). For instance, in cattle, genital microbiota is different between Gyr, Nellore, and Holstein breeds (3). The general microbiome of the newborn is similar to the mother’s skin microbiome if the delivery method was caesarean section (4) and is similar to the vaginal microbiome if it was natural delivery (5). Estrogen and progesterone hormone concentration variations during the estrous cycle influence bacterial growth in the genitalia by favoring some species at different times (6). During pregnancy, bacterial quantity and diversity decrease while archaeal abundance increases in vaginal milieu (7). Vaginal and uterine microbiomes of cows not diagnosed with metritis during the first month postpartum are similar in cows without uterine infections but differ from those with uterine infections (8). Dietary quality and quantity peripartum alter the endometrial microbiome through the provision of energy and protein nutrients (9). The uterine microbiome between 10 and 35 days postpartum is similar in cows not diagnosed with subclinical endometritis and those that will develop subclinical endometritis (10). Rumen, skin, rectum or feces (11) contribute to the establishment of the genital microbiome, while the environment (12) and intravaginal antibiotic therapy (13) can also alter the endometrial microbiota during a female’s lifetime.
5.1.3. Delivery Mode
5.1.4. Estrous Cyclicity
5.1.5. Pregnancy
5.1.6. Postpartum
5.2. Extrinsic Factors
5.2.1. Nutrition
5.2.2. Genital Pathologies
References
- Jeffrey D. Swartz; Medora Lachman; Kelsey Westveer; Thomas O’Neill; Thomas Geary; Rodney W. Kott; James G. Berardinelli; Patrick G. Hatfield; Jennifer M. Thomson; Andrew Roberts; et al.Carl J. Yeoman Characterization of the Vaginal Microbiota of Ewes and Cows Reveals a Unique Microbiota with Low Levels of Lactobacilli and Near-Neutral pH. Frontiers in Veterinary Science 2014, 1, 19, 10.3389/fvets.2014.00019.The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth A. Miller; Deanna E. Beasley; Robert R. Dunn; Elizabeth A. Archie; Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique?. Jeffrey D. Swartz; Medora Lachman; Kelsey Westveer; Thomas O’Neill; Thomas Geary; Rodney W. Kott; James G. Berardinelli; Patrick G. Hatfield; Jennifer M. Thomson; Andrew Roberts; et al.Carl J. Yeoman Characterization of the Vaginal Microbiota of Ewes and Cows Reveals a Unique Microbiota with Low Levels of Lactobacilli and Near-Neutral pH. Frontiers in MVetericrobiology nary Science 2016, 7, 1936, 10.3389/fmicb.2016.01936.4, 1, 19, 10.3389/fvets.2014.00019.
- Wallace Jeng Yang Chee, Shu Yih Chew & Leslie Thian Lung Than; Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Elizabeth A. Miller; Deanna E. Beasley; Robert R. Dunn; Elizabeth A. Archie; Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique?. Frontiers in Microbiaol Cell Factories ogy 2020, 19, 203, https://doi.org/10.1186/s12934-020-01464-4.16, 7, 1936, 10.3389/fmicb.2016.01936.
- Deirdre E. O’Hanlon; Thomas R. Moench; Richard A. Cone; Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLWallace Jeng Yang Chee, Shu Yih Chew & Leslie Thian Lung Than; Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. MicroSbial Cell ONE Factories 202013, 8, e80074, 10.1371/journal.pone.0080074., 19, 203, https://doi.org/10.1186/s12934-020-01464-4.
- Gilda Tachedjian; Muriel Aldunate; Catronia S. Bradshaw; Richard A. Cone; The role of lactic acid production by probiotic Lactobacillus species in vaginal health. ResearchDeirdre E. O’Hanlon; Thomas R. Moench; Richard A. Cone; Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLoS in Microbiology ONE 2017, 163, 8, 782-792, 10.1016/j.resmic.2017.04.001., e80074, 10.1371/journal.pone.0080074.
- Paola Nardini; Rogers Alberto Nahui Palomino; Carola Parolin; Luca Laghi; Claudio Foschi; Roberto Cevenini; Beatrice Vitali; Antonella Marangoni; Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. SGilda Tachedjian; Muriel Aldunate; Catronia S. Bradshaw; Richard A. Cone; The role of lactic acid production by probiotic Lactobacillus species in vaginal health. Research ientific Reports n Microbiology 2016, 7, 16, 29024, 10.1038/srep29024.8, 782-792, 10.1016/j.resmic.2017.04.001.
- Michelle A Graver; Jeremy J Wade; The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth. Annals of CliniPaola Nardini; Rogers Alberto Nahui Palomino; Carola Parolin; Luca Laghi; Claudio Foschi; Roberto Cevenini; Beatrice Vitali; Antonella Marangoni; Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. Scal Microbiology and Antimicrobialentific Reports 2011, 10, 8-8, 10.1186/1476-0711-10-8.6, 6, 29024, 10.1038/srep29024.
- Erika V. Valore; Christina H. Park; Sorina L. Igreti; Tomas Ganz; Antimicrobial components of vaginal fluid. Michelle A Graver; Jeremy J Wade; The role of acidification in the inhibition of Neisseria gonorrhoeae by vaginal lactobacilli during anaerobic growth. Americnnan Journal of Obstetrics and Gynecology ls of Clinical Microbiology and Antimicrobials 2002, 11, 187, 561-568, 10.1067/mob.2002.125280.0, 8-8, 10.1186/1476-0711-10-8.
- Taha E. Taha; Donald R. Hoover; Gina A. Dallabetta; Newton I. Kumwenda; Laban A. R. Mtimavalye; Li-Ping Yang; George N. Liomba; Robin L. Broadhead; John D. Chiphangwi; Paolo G. Miotti; et al. Bacterial vaginosis and disturbances of vaginal flora. Erika V. Valore; Christina H. Park; Sorina L. Igreti; Tomas Ganz; Antimicrobial components of vaginal fluid. AIDS 1998, merican Journal of Obstetrics and Gynecology 2002, 12, 1699-1706, 10.1097/00002030-199813000-00019.87, 561-568, 10.1067/mob.2002.125280.
- Thuy Hoang; Emily Toler; Kevin Delong; Nomfuneko A. Mafunda; Seth M. Bloom; Hannah C. Zierden; Thomas R. Moench; Jenell S. Coleman; Justin Hanes; Douglas S. Kwon; et al.Samuel K. LaiRichard A. ConeLaura M. Ensign The cervicovaginal mucus barrier to HIV-1 is diminished in bacterial vaginosis. PLoTaha E. Taha; Donald R. Hoover; Gina A. Dallabetta; Newton I. Kumwenda; Laban A. R. Mtimavalye; Li-Ping Yang; George N. Liomba; Robin L. Broadhead; John D. Chiphangwi; Paolo G. Miotti; et al. Bacterial vaginosis and disturbances of vaginal flora. AIDS Pathogens 2020, 1998, 16, e1008236, 10.1371/journal.ppat.1008236.2, 1699-1706, 10.1097/00002030-199813000-00019.
- Muriel Aldunate; David Tyssen; Adam Johnson; Tasnim Zakir; Secondo Sonza; Thomas Moench; Richard Cone; Gilda Tachedjian; Vaginal concentrations of lactic acid potently inactivate HIV. JThuy Hoang; Emily Toler; Kevin Delong; Nomfuneko A. Mafunda; Seth M. Bloom; Hannah C. Zierden; Thomas R. Moench; Jenell S. Coleman; Justin Hanes; Douglas S. Kwon; et al.Samuel K. LaiRichard A. ConeLaura M. Ensign The cervicovaginal mucus barrier to HIV-1 is diminished in bacterial vaginosis. PLournS Pal of Antimicrobial Chemotherapy thogens 202013, , 168, 2015-2025, 10.1093/jac/dkt156., e1008236, 10.1371/journal.ppat.1008236.
- M. Srinivasan; M. Adnane; G. Archunan; Significance of cervico-vaginal microbes in bovine reproduction and pheromone production – A hypothetical review. ReseMuriel Aldunate; David Tyssen; Adam Johnson; Tasnim Zakir; Secondo Sonza; Thomas Moench; Richard Cone; Gilda Tachedjian; Vaginal concentrations of lactic acid potently inactivate HIV. Journal of Antimicrch in Veterinarobial Chemotherapy Science 2021, 135, 66-71, 10.1016/j.rvsc.2021.01.003.3, 68, 2015-2025, 10.1093/jac/dkt156.
- Archunan, G.; Rajanarayanan, S.; Karthikeyan, K. Neurobiology of Chemical Communication; Carla Mucignat-Caretta, Eds.; CRC Press/Taylor & Francis LLC: Boca Raton, FL, USA,, 2014; pp. 461-479.M. Srinivasan; M. Adnane; G. Archunan; Significance of cervico-vaginal microbes in bovine reproduction and pheromone production – A hypothetical review. Research in Veterinary Science 2021, 135, 66-71, 10.1016/j.rvsc.2021.01.003.
- G Archunan; Reproductive enhancement in buffalo: looking at urinary pheromones and hormones.. Iranian journal of veterinary research 2020, 21, 163-171.Archunan, G.; Rajanarayanan, S.; Karthikeyan, K. Neurobiology of Chemical Communication; Carla Mucignat-Caretta, Eds.; CRC Press/Taylor & Francis LLC: Boca Raton, FL, USA,, 2014; pp. 461-479.
- S. LeClaire; J. F. Nielsen; C. M. Drea; Bacterial communities in meerkat anal scent secretions vary with host sex, age, and group membership. BehG Archunan; Reproductive enhancement in buffalo: looking at urinary pheromones and hormones.. Iravnioral Ecologan journal of veterinary research 202014, , 25, 996-1004, 10.1093/beheco/aru074.1, 163-171.
- R Sankar; G Archunan; Flehmen response in bull: role of vaginal mucus and other body fluids of bovine with special reference to estrus. S. LeClaire; J. F. Nielsen; C. M. Drea; Bacterial communities in meerkat anal scent secretions vary with host sex, age, and group membership. Behavioural Processes Ecology 20014, 67, 81-86, 10.1016/j.beproc.2004.02.007., 25, 996-1004, 10.1093/beheco/aru074.
- Sarah LeClaire; Staffan Jacob; Lydia K. Greene; George R. Dubay; Christine M. Drea; Social odours covary with bacterial community in the anal secretions of wild meerkats. SciR Sankar; G Archunan; Flehmen response in bull: role of vaginal mucus and other body fluids of bovine with special reference to estrus. Benthavific Reportoural Processes 20017, 4, 67, 1-13, 10.1038/s41598-017-03356-x., 81-86, 10.1016/j.beproc.2004.02.007.
- Daniel Lazzari Quadros; Ricardo Zanella; Carlos Bondan; Giovana Ciacci Zanella; Fernanda Luiza Facioli; Arthur Nery da Silva; Eraldo Lourenso Zanella; Study of vaginal microbiota of Holstein cows submitted to an estrus synchronization protocol with the use of intravaginal progesterone device. ResearSarah LeClaire; Staffan Jacob; Lydia K. Greene; George R. Dubay; Christine M. Drea; Social odours covary with bacterial community in the anal secretions of wild meerkats. Sch in Veterinary Sciencentific Re ports 2020, 131, 1-6, 10.1016/j.rvsc.2020.03.027.17, 7, 1-13, 10.1038/s41598-017-03356-x.
- Maria G. Dominguez-Bello; Elizabeth K. Costello; Monica Contreras; Magda Magris; Glida Hidalgo; Noah Fierer; Rob Knight; Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. PDaniel Lazzari Quadros; Ricardo Zanella; Carlos Bondan; Giovana Ciacci Zanella; Fernanda Luiza Facioli; Arthur Nery da Silva; Eraldo Lourenso Zanella; Study of vaginal microbiota of Holstein cows submitted to an estrus synchronization protocol with the use of intravaginal progesterone device. Researoceedings of the National Academy of h in Veterinary Sciences 20120, , 13107, 11971-11975, 10.1073/pnas.1002601107., 1-6, 10.1016/j.rvsc.2020.03.027.
- Sarkis K. Mazmanian; Cui Hua Liu; Arthur O. Tzianabos; Dennis L. Kasper; An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. CMaria G. Dominguez-Bello; Elizabeth K. Costello; Monica Contreras; Magda Magris; Glida Hidalgo; Noah Fierer; Rob Knight; Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proceedings of the Nationall Academy of Sciences 20105, , 122, 107-118, 10.1016/j.cell.2005.05.007.07, 11971-11975, 10.1073/pnas.1002601107.
- Hannah M. Wexler; Shyam Daya; Kenneth I. Berns; Bacteroides : the Good, the Bad, and the Nitty-Gritty. Sarkis K. Mazmanian; Cui Hua Liu; Arthur O. Tzianabos; Dennis L. Kasper; An Immunomodulatory Molecule of Symbiotic Bacteria Directs Maturation of the Host Immune System. Celinical Microbiology Reviews 2007, 5, 1220, 593-621, 10.1128/cmr.00008-07., 107-118, 10.1016/j.cell.2005.05.007.
- Nicole R. Kraipowich; David L. Morris; Gayle L. Thompson; Gary L. Mason; Bovine abortions associated with Bacteroides fragilis fetal infection.. JourHannah M. Wexler; Shyam Daya; Kenneth I. Berns; Bacteroides : the Good, the Bad, and the Nitty-Gritty. Clinical of Veterinary Diagnostic InveMicrobiology Reviewstigation 2000, 17, 2, 369-371, 10.1177/104063870001200413.0, 593-621, 10.1128/cmr.00008-07.
- Mateus Laguardia-Nascimento; Kelly Moreira Grillo Ribeiro Branco; Marcela Ribeiro Gasparini; Silvia Giannattasio-Ferraz; Laura Rabelo Leite; Flávio Marcos Gomes Araújo; Anna Christina De Matos Salim; Jacques Robert Nicoli; Guilherme Correa De Oliveira; Edel Barbosa-Stancioli; et al. Vaginal Microbiome Characterization of Nellore Cattle Using Metagenomic Analysis. PLNicole R. Kraipowich; David L. Morris; Gayle L. Thompson; Gary L. Mason; Bovine abortions associated with Bacteroides fragilis fetal infection.. Journal of Veterinary DiagnoSstic ONE Investigation 20015, 0, 10, e0143294, 10.1371/journal.pone.0143294.2, 369-371, 10.1177/104063870001200413.
- Feilong Deng; Maryanna McClure; Rick Rorie; Xiaofan Wang; Jianmin Chai; Xiaoyuan Wei; Songjia Lai; Jiangchao Zhao; The vaginal and fecal microbiomes are related to pregnancy status in beef heifers. Journal Mateus Laguardia-Nascimento; Kelly Moreira Grillo Ribeiro Branco; Marcela Ribeiro Gasparini; Silvia Giannattasio-Ferraz; Laura Rabelo Leite; Flávio Marcos Gomes Araújo; Anna Christina De Matos Salim; Jacques Robert Nicoli; Guilherme Correa De Oliveira; Edel Barbosa-Stancioli; et al. Vaginal Microbiome Characterization of Nellore Cattle Using Metagenomic Analysis. PLof Animal Science and Biotechnology ONE 2019, 5, 10, 1-13, 10.1186/s40104-019-0401-2., e0143294, 10.1371/journal.pone.0143294.
- Klibs N. Galvão; Rodrigo C. Bicalho; Soo Jin Jeon; Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. Feilong Deng; Maryanna McClure; Rick Rorie; Xiaofan Wang; Jianmin Chai; Xiaoyuan Wei; Songjia Lai; Jiangchao Zhao; The vaginal and fecal microbiomes are related to pregnancy status in beef heifers. Journal of DAnimairyl Science and Biote chnology 2019, 102, 11786-11797, 10.3168/jds.2019-17106., 1-13, 10.1186/s40104-019-0401-2.
- N.F. Rodrigues; J. Kästle; Tarcisio José Domingos Coutinho; Aline Teixeira Amorim; G.B. Campos; V.M. Santos; L.M. Marques; J. Timenetsky; S.T. De Farias; Qualitative analysis of the vaginal microbiota of healthy cattle and cattle with genital-tract disease. GeKlibs N. Galvão; Rodrigo C. Bicalho; Soo Jin Jeon; Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. Journetics and Molecular Resl of Dairy Sciencearch 2015, 9, 14, 6518-6528, 10.4238/2015.june.12.4.02, 11786-11797, 10.3168/jds.2019-17106.
- Riley Messman; Zully E Contreras-Correa; Henry A Paz; George Perry; Caleb O Lemley; Vaginal bacterial community composition and concentrations of estradiol at the time of artificial insemination in Brangus heifers. JourN.F. Rodrigues; J. Kästle; Tarcisio José Domingos Coutinho; Aline Teixeira Amorim; G.B. Campos; V.M. Santos; L.M. Marques; J. Timenetsky; S.T. De Farias; Qualitative analysis of the vaginal microbiota of healthy cattle and cattle with genital-tract disease. Genetics al of Animal Science nd Molecular Research 2020, 98, 98, 10.1093/jas/skaa178.15, 14, 6518-6528, 10.4238/2015.june.12.4.
- Cecilia Christensen Karstrup; Kirstine Klitgaard; Tim Kåre Jensen; Jørgen Steen Agerholm; Hanne Gervi Pedersen; Presence of bacteria in the endometrium and placentomes of pregnant cows. TheRiley Messman; Zully E Contreras-Correa; Henry A Paz; George Perry; Caleb O Lemley; Vaginal bacterial community composition and concentrations of estradiol at the time of artificial insemination in Brangus heifers. Journal of Aniogenology mal Science 202017, , 99, 41-47, 10.1016/j.theriogenology.2017.05.013.8, 98, 10.1093/jas/skaa178.
- Stephen Moore; A. C. Ericsson; S. E. Poock; P. Melendez; M. C. Lucy; Hot topic: 16S rRNA gene sequencing reveals the microbiome of the virgin and pregnant bovine uterus. JCecilia Christensen Karstrup; Kirstine Klitgaard; Tim Kåre Jensen; Jørgen Steen Agerholm; Hanne Gervi Pedersen; Presence of bacteria in the endometrium and placentomes of pregnant cows. Theriourgenal of Dairology Science 2017, 100, 4953-4960, 10.3168/jds.2017-12592., 99, 41-47, 10.1016/j.theriogenology.2017.05.013.
- I. Martin Sheldon; James G. Cronin; John J. Bromfield; Tolerance and Innate Immunity Shape the Development of Postpartum Uterine Disease and the Impact of Endometritis in Dairy Cattle. AStephen Moore; A. C. Ericsson; S. E. Poock; P. Melendez; M. C. Lucy; Hot topic: 16S rRNA gene sequencing reveals the microbiome of the virgin and pregnant bovine uterus. Journnual Review of Animal Biosof Dairy Sciences 2019, 7, 361-384, 10.1146/annurev-animal-020518-115227.7, 100, 4953-4960, 10.3168/jds.2017-12592.
- Guda van der Burgt; Wendy Clark; Richard Knight; Cattle fertility problems and Histophilus somni. VI. Martin Sheldon; James G. Cronin; John J. Bromfield; Tolerance and Innate Immunity Shape the Development of Postpartum Uterine Disease and the Impact of Endometritis in Dairy Cattle. Annual Retvierinary Record w of Animal Biosciences 2007, 160, 600-600, 10.1136/vr.160.17.600.19, 7, 361-384, 10.1146/annurev-animal-020518-115227.
- Nicholas, R.; Ayling, R.; McAuliffe. Mycoplasma Diseases of Ruminants; CAB International: Oxfordshire, UK, 2008; pp. 208-215.Guda van der Burgt; Wendy Clark; Richard Knight; Cattle fertility problems and Histophilus somni. Veterinary Record 2007, 160, 600-600, 10.1136/vr.160.17.600.
- S. Kovachev; Defence factors of vaginal lactobacilli. Critical Reviews in Microbiology 2017, 44, 31-39, 10.1080/1040841x.2017.1306688.Nicholas, R.; Ayling, R.; McAuliffe. Mycoplasma Diseases of Ruminants; CAB International: Oxfordshire, UK, 2008; pp. 208-215.
- Gaoyan Tang; Todd Kitten; Cindy L. Munro; George C. Wellman; Keith P. Mintz; EmaA, a Potential Virulence Determinant of Aggregatibacter actinomycetemcomitans in Infective Endocarditis. InfS. Kovachev; Defence factors of vaginal lactobacilli. Critical Rectvion and Immunitews in Microbiology 2008, 76, 2316-2324, 10.1128/iai.00021-08.17, 44, 31-39, 10.1080/1040841x.2017.1306688.
- Daniela Ulrich; Sharon L. Edwards; Vincent Letouzey; Kai Su; Jacinta F. White; Anna Rosamilia; Caroline E. Gargett; Jerome A. Werkmeister; Regional Variation in Tissue Composition and Biomechanical Properties of Postmenopausal Ovine and Human Vagina. PLGaoyan Tang; Todd Kitten; Cindy L. Munro; George C. Wellman; Keith P. Mintz; EmaA, a Potential Virulence Determinant of Aggregatibacter actinomycetemcomitans in Infective Endocarditis. InfectioSn and ONE Immunity 2014, 9, e104972, 10.1371/journal.pone.0104972.08, 76, 2316-2324, 10.1128/iai.00021-08.
- The Human Microbiome Project Consortium; Structure, function and diversity of the healthy human microbiome. Daniela Ulrich; Sharon L. Edwards; Vincent Letouzey; Kai Su; Jacinta F. White; Anna Rosamilia; Caroline E. Gargett; Jerome A. Werkmeister; Regional Variation in Tissue Composition and Biomechanical Properties of Postmenopausal Ovine and Human Vagina. PLoS ONature E 2012, 486, 207-214, 10.1038/nature11234.4, 9, e104972, 10.1371/journal.pone.0104972.
- I. Sheldon; D. Noakes; A. Rycroft; Dirk Pfeiffer; H Dobson; Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. REPRODUCTIOThe Human Microbiome Project Consortium; Structure, function and diversity of the healthy human microbiome. N ature 20012, 123, 837-845, 10.1530/reprod/123.6.837., 486, 207-214, 10.1038/nature11234.
- Erin Williams; D.P. Fischer; D.E. Noakes; Gary England; A. Rycroft; H. Dobson; I.M. Sheldon; The relationship between uterine pathogen growth density and ovarian function in the postpartum dairy cow. I. Sheldon; D. Noakes; A. Rycroft; Dirk Pfeiffer; H Dobson; Influence of uterine bacterial contamination after parturition on ovarian dominant follicle selection and follicle growth and function in cattle. REPRODUCTheriogenology ION 2007, 68, 549-559, 10.1016/j.theriogenology.2007.04.056.2, 123, 837-845, 10.1530/reprod/123.6.837.
- C. Otero; C. Silva de Ruiz; R. Ibañez; O.R. Wilde; A.A.P. De Ruiz Holgado; M.E. Nader-Macias; Lactobacilli and Enterococci Isolated from the Bovine Vagina During the Estrous cycle. AnaErin Williams; D.P. Fischer; D.E. Noakes; Gary England; A. Rycroft; H. Dobson; I.M. Sheldon; The relationship between uterine pathogen growth density and ovarian function in the postpartum dairy cow. Theriobge 1999, 5, 305-307, 10.1006/anae.1999.0245.nology 2007, 68, 549-559, 10.1016/j.theriogenology.2007.04.056.
- C. Otero; L. Saavedra; C. Silva De Ruiz; O. Wilde; A.R. Holgado; M.E. Nader-Macias; Vaginal bacterial microflora modifications during the growth of healthy cows. Letters iC. Otero; C. Silva de Ruiz; R. Ibañez; O.R. Wilde; A.A.P. De Ruiz Holgado; M.E. Nader-Macias; Lactobacilli and Enterococci Isolated from the Bovine Vagina During the Estrous cycle. An Appliaed Microbiology 2000, 31, 251-254, 10.1046/j.1365-2672.2000.00809.x.robe 1999, 5, 305-307, 10.1006/anae.1999.0245.
- G. J. Sawyer; OBSERVATIONS ON THE BACTERIAL POPULATION OF THE OS CERVIX OF THE EWE BEFORE AND AFTER EMBRYO DEATH. AusC. Otero; L. Saavedra; C. Silva De Ruiz; O. Wilde; A.R. Holgado; M.E. Nader-Macias; Vaginal bacterial microflora modifications during the growth of healthy cows. Letteralian Veterinary Journas in Applied Microbiol 1977, 5ogy 2000, 3, 542-544, 10.1111/j.1751-0813.1977.tb07942.x.1, 251-254, 10.1046/j.1365-2672.2000.00809.x.
- J. Manes; M.A. Fiorentino; G. Kaiser; F. Hozbor; R. Alberio; E. Sanchez; F. Paolicchi; Changes in the aerobic vaginal flora after treatment with different intravaginal devices in ewes. SmG. J. Sawyer; OBSERVATIONS ON THE BACTERIAL POPULATION OF THE OS CERVIX OF THE EWE BEFORE AND AFTER EMBRYO DEATH. Australl Ruminant Research 20ian Veterinary Journal 10, 94, 201-204, 10.1016/j.smallrumres.2010.07.021.977, 53, 542-544, 10.1111/j.1751-0813.1977.tb07942.x.
- Raúl Miranda-CasoLuengo; Junnan Lu; Erin J. Williams; Aleksandra A. Miranda-CasoLuengo; Stephen D. Carrington; Alexander C. O. Evans; Wim G. Meijer; Delayed differentiation of vaginal and uterine microbiomes in dairy cows developing postpartum endometritis. PLoJ. Manes; M.A. Fiorentino; G. Kaiser; F. Hozbor; R. Alberio; E. Sanchez; F. Paolicchi; Changes in the aerobic vaginal flora after treatment with different intravaginal devices in ewes. Small ONE Ruminant Research 2019, 10, 94, e0200974, 10.1371/journal.pone.0200974., 201-204, 10.1016/j.smallrumres.2010.07.021.
- Silvia Giannattasio-Ferraz; Mateus Laguardia-Nascimento; Marcela Ribeiro Gasparini; Laura Rabelo Leite; Flávio Marcos Gomes Araujo; Anna Christina De Matos Salim; André Penido De Oliveira; Jacques Robert Nicoli; Guilherme Corrêa De Oliveira; Flavio Guimarães Da Fonseca; et al.Edel Figueiredo Barbosa-Stancioli A common vaginal microbiota composition among breeds of Bos taurus indicus (Gyr and Nellore). Brazilian JRaúl Miranda-CasoLuengo; Junnan Lu; Erin J. Williams; Aleksandra A. Miranda-CasoLuengo; Stephen D. Carrington; Alexander C. O. Evans; Wim G. Meijer; Delayed differentiation of vaginal and uterine microbiomes in dairy cows developing postpartum endometritis. PLournalS of Microbiology ONE 2019, 50, 1115-1124, 10.1007/s42770-019-00120-3., 14, e0200974, 10.1371/journal.pone.0200974.
- George I. Gorodeski; Ulrich Hopfer; Chung Chiun Liu; Ellen Margles; Estrogen Acidifies Vaginal pH by Up-Regulation of Proton Secretion via the Apical Membrane of Vaginal-Ectocervical Epithelial Cells. ESilvia Giannattasio-Ferraz; Mateus Laguardia-Nascimento; Marcela Ribeiro Gasparini; Laura Rabelo Leite; Flávio Marcos Gomes Araujo; Anna Christina De Matos Salim; André Penido De Oliveira; Jacques Robert Nicoli; Guilherme Corrêa De Oliveira; Flavio Guimarães Da Fonseca; et al.Edel Figueiredo Barbosa-Stancioli A common vaginal microbiota composition among breeds of Bos taurus indicus (Gyr and Nellore). Brazilian Journdal ocrinf Microbiology 2005, 146, 816-824, 10.1210/en.2004-1153.19, 50, 1115-1124, 10.1007/s42770-019-00120-3.
- G.A. Perry; B.L. Perry; Effect of preovulatory concentrations of estradiol and initiation of standing estrus on uterine pH in beef cows. Domestic Animal George I. Gorodeski; Ulrich Hopfer; Chung Chiun Liu; Ellen Margles; Estrogen Acidifies Vaginal pH by Up-Regulation of Proton Secretion via the Apical Membrane of Vaginal-Ectocervical Epithelial Cells. Endocrinology 2008, 35, 14, 333-338, 10.1016/j.domaniend.2007.09.003.6, 816-824, 10.1210/en.2004-1153.
- Pawel Gajer; Rebecca M. Brotman; Guoyun Bai; Joyce Sakamoto; Ursel M. E. Schütte; Xue Zhong; Sara S. K. Koenig; Li Fu; Zhanshan (Sam) Ma; Xia Zhou; et al.Zaid AbdoLarry J. ForneyJacques Ravel Temporal Dynamics of the Human Vaginal Microbiota. ScG.A. Perry; B.L. Perry; Effect of preovulatory concentrations of estradiol and initiation of standing estrus on uterine pH in beef cows. Domestience Translational Medicine Animal Endocrinology 2012, 08, 34, 132ra52-132ra52, 10.1126/scitranslmed.3003605., 333-338, 10.1016/j.domaniend.2007.09.003.
- Kjersti Aagaard; Kevin Riehle; Jun Ma; Nicola Segata; Toni-Ann Mistretta; Cristian Coarfa; Sabeen Raza; Sean Rosenbaum; Ignatia Van Den Veyver; Aleksandar Milosavljevic; et al.Dirk GeversCurtis HuttenhowerJoseph PetrosinoJames Versalovic A Metagenomic Approach to Characterization of the Vaginal Microbiome Signature in Pregnancy. PLPawel Gajer; Rebecca M. Brotman; Guoyun Bai; Joyce Sakamoto; Ursel M. E. Schütte; Xue Zhong; Sara S. K. Koenig; Li Fu; Zhanshan (Sam) Ma; Xia Zhou; et al.Zaid AbdoLarry J. ForneyJacques Ravel Temporal Dynamics of the Human Vaginal Microbiota. Science TranslatioSnal ONE Medicine 2012, 7, e36466, 10.1371/journal.pone.0036466., 4, 132ra52-132ra52, 10.1126/scitranslmed.3003605.
- Giulia Esposito; Emiliano Raffrenato; Somwe D Lukamba; Mounir Adnane; Pete C Irons; Paul Cormican; Taurai Tasara; Aspinas Chapwanya; Characterization of metabolic and inflammatory profiles of transition dairy cows fed an energy-restricted diet. JKjersti Aagaard; Kevin Riehle; Jun Ma; Nicola Segata; Toni-Ann Mistretta; Cristian Coarfa; Sabeen Raza; Sean Rosenbaum; Ignatia Van Den Veyver; Aleksandar Milosavljevic; et al.Dirk GeversCurtis HuttenhowerJoseph PetrosinoJames Versalovic A Metagenomic Approach to Characterization of the Vaginal Microbiome Signature in Pregnancy. PLournalS of Animal Science ONE 20120, 98, 98, 10.1093/jas/skz391., 7, e36466, 10.1371/journal.pone.0036466.
- I. Martin Sheldon; Erin Williams; Aleisha N.A. Miller; Deborah Mary Nash; Shan Herath; Uterine diseases in cattle after parturition. The VeteGiulia Esposito; Emiliano Raffrenato; Somwe D Lukamba; Mounir Adnane; Pete C Irons; Paul Cormican; Taurai Tasara; Aspinas Chapwanya; Characterization of metabolic and inflammatory profiles of transition dairy cows fed an energy-restricted diet. Jourinary Journl of Animal Science 20208, 176, 115-121, 10.1016/j.tvjl.2007.12.031., 98, 98, 10.1093/jas/skz391.
- Federico Cunha; Soo Jin Jeon; Rodolfo Daetz; Achilles Vieira Neto; Jimena Laporta; K. Casey Jeong; Anthony F. Barbet; Carlos A. Risco; Klibs N. Galvão; Quantifying known and emerging uterine pathogens, and evaluating their association with metritis and fever in dairy cows. I. Martin Sheldon; Erin Williams; Aleisha N.A. Miller; Deborah Mary Nash; Shan Herath; Uterine diseases in cattle after parturition. The Veteriogenonary Journalogy 20018, 114, 25-33, 10.1016/j.theriogenology.2018.03.016.76, 115-121, 10.1016/j.tvjl.2007.12.031.
- M.L.S. Bicalho; V.S. Machado; G. Oikonomou; Robert Gilbert; R.C. Bicalho; Association between virulence factors of Escherichia coli, Fusobacterium necrophorum, and Arcanobacterium pyogenes and uterine diseases of dairy cows. VetFederico Cunha; Soo Jin Jeon; Rodolfo Daetz; Achilles Vieira Neto; Jimena Laporta; K. Casey Jeong; Anthony F. Barbet; Carlos A. Risco; Klibs N. Galvão; Quantifying known and emerging uterine pathogens, and evaluating their association with metritis and fever in dairy cows. Theriogenary Microbioology 2012, 8, 1157, 125-131, 10.1016/j.vetmic.2011.11.034.4, 25-33, 10.1016/j.theriogenology.2018.03.016.
- Rebecca M. Brotman; Michelle D. Shardell; Pawel Gajer; Doug Fadrosh; Kathryn Chang; Michelle I. Silver; Raphael P. Viscidi; Anne E. Burke; Jacques Ravel; Patti E. Gravitt; et al. Association between the vaginal microbiota, menopause status, and signs of vulvovaginal atrophy. MM.L.S. Bicalho; V.S. Machado; G. Oikonomou; Robert Gilbert; R.C. Bicalho; Association between virulence factors of Escherichia coli, Fusobacterium necrophorum, and Arcanobacterium pyogenes and uterine diseases of dairy cows. Veterinopause ary Microbiology 2018, 22, 15, 1321-1330, 10.1097/gme.0000000000001236.7, 125-131, 10.1016/j.vetmic.2011.11.034.
- N.Y. Shpigel; L. Adler-Ashkenazy; S. Scheinin; T. Goshen; A. Arazi; Z. Pasternak; Y. Gottlieb; Characterization and identification of microbial communities in bovine necrotic vulvovaginitis. ThRebecca M. Brotman; Michelle D. Shardell; Pawel Gajer; Doug Fadrosh; Kathryn Chang; Michelle I. Silver; Raphael P. Viscidi; Anne E. Burke; Jacques Ravel; Patti E. Gravitt; et al. Association between the vaginal microbiota, menopause status, and signs of vulvovaginal atrophy. Me Veterinary Journal opause 2017, 8, 219, 34-39, 10.1016/j.tvjl.2016.12.002.5, 1321-1330, 10.1097/gme.0000000000001236.
- N.Y. Shpigel; L. Adler-Ashkenazy; S. Scheinin; T. Goshen; A. Arazi; Z. Pasternak; Y. Gottlieb; Characterization and identification of microbial communities in bovine necrotic vulvovaginitis. The Veterinary Journal 2017, 219, 34-39, 10.1016/j.tvjl.2016.12.002.