Camelina (Camelina sativa (L.) Crantz) is a protein and oilseed crop belonging to the family Brassicaceae.
- camelina
- poultry
- n-3 fatty acids
- meat quality
1. Introduction
About 75% of protein feeds required for the balance of livestock feed rations is imported into the EU. Thus, the sustainable production strategy can be implemented by reducing the dependency on critical feed materials, for example, soya, which is grown on deforested land. One of the ways to solve the problem would be growing alternative protein crop varieties that are well-adapted to certain rural area conditions, require less fertilizers and are resistant to diseases and agricultural pests.
Currently, human nutrition is deficient of n-3 polyunsaturated fatty acids (PUFA). Epidemiologic studies indicate that such deficiency could be the cause of coronary heart disease, cancer and depression. One of the solutions to the problem could be poultry production with increased n-3 PUFA content by supplementing poultry with diets rich in n-3 PUFA feeds as, for example, camelina seed, cake or oil.
Camelina (Camelina sativa (L.) Crantz) is a protein and oilseed crop belonging to the family Brassicaceae. Numerous archaeobotanical studies indicate that camelina has a long history of its cultivation in Europe and Asia Minor. In many European countries camelina was grown as an agricultural crop until the mid-20th century.
Camelina has recently attracted great interest as an oil crop for biodiesel, jet fuel and oil production with a low production cost. Large quantities of by-products (cake, meal) are left after oil extraction. Camelina expeller, or cake, is a product of oil extraction obtained by pressing camelina seed. Cake contains from 6.4 to 22.7% residual oil.
These products have good potential to be used as a cheap alternative protein feedstuff and a valuable source in animal nutrition. Camelina by-products are comparatively cheaper than other sources, especially soybean.
The objective of this review is to discuss chemical, amino and fatty acid composition of various feed ingredients produced from camelina and their effects on meat type poultry growth performance, carcass traits, meat chemical composition and tissue fatty acid profile.
2. Camelina Chemical Composition
Camelina is a valuable source of protein. Different camelina feed components contain different amounts of protein. Camelina seed contains 24.78% protein, while seed by-products, such as cake, have higher protein content, respectively, 30.33–39.80% .
The crude protein content in camelina cake is close to that found in rapeseed meal (29.69–39.89%), but lower than in soybean meal (43.0–56.3%).
The crude fat content found in camelina seed is high (36.84%). However, the crude fat content in cake is in the range of 6.44–22.71%. The crude fat content in camelina cake is higher than in soybean (0.55–3.3%), and rapeseed (1.4–10.50%) meals.
Methionine with cystine and lysine are the first limiting acids in poultry nutrition. The content of methionine with cystine (3.80–5.17% of total protein) in camelina cake is higher than soybean meal (2.61–3.27% of total protein) and similar to that in rapeseed meal (4.12–5.01% of total protein). The content of lysine in camelina is 1.55–2.02 and 1.02–1.56 times lower than that in soybean meal and rapeseed meal, respectively.
The content of arginine, valine, alanine and proline in camelina are lower than in soybean meal, but by total protein content, the difference is insignificant. In comparison with rapeseed meal, camelina cake has a similar content of alanine, asparagine, glycine, isoleucine, leucine, phenylalanine, serine, tryptophan, tyrosine and valine.
2.1. Camelina Fatty Acid Composition
The percentage of highly important n-3 ALA is usually low in the main feed ingredients of plant origin. Thus, the amount of ALA usually found in wheat is 0.06–0.14%, in corn 0.48–0.50%, in sunflower 0.15–0.27% and in barley 0.35%.
Camelina seed, oil and its processing products have a much higher ALA content (25.88–36.67%) than other common feed components, i.e., soybean meal (7.21–8.58%), hempseed cake (15.85–24.7%), and rapeseed cake (10.60–13.05%), but lower than that found in linseed cake (51.5%). Consequently, it may be maintained that camelina is the second highest, by ALA content, plant growing in the northern hemisphere, and suitable as a feed component.
Essential n-6 PUFA linoleic (C18:2n-6; LA) fatty acid content, which should be as low as possible in poultry feed, amounts to 18.84–24.16% in camelina seed and cake, and this percentage is lower than that found in rapeseed cake (21.67–23.5%), soybean meal (52.43–55.20%) and hempseed cake (52.5–59.52%), but higher than that found in linseed cake (14.60%).
The content of total n-3 PUFA, which is always deficient in standard poultry feeding, is from 3.11 to 4.00, 2.05 to 3.96 and 1.0 to 2.16 times higher in camelina cake and seed, than that found in soybean meal, rapeseed cake and hempseed cake, respectively, but from 1.50 to 2.19 times lower than in linseed cake [50,52,59,60]. Camelina cake and oil has a lower n-6/n-3 PUFA ratio in comparison with soybean meal, rapeseed cake and hempseed cake, respectively, 0.60–0.99 vs. 6.10, 1.68, 4.08 [37,51,52]. The linoleic/α-linolenic ratio in camelina cake (0.72) was also lower compared with rapeseed cake (1.66), hempseed cake (3.76) and soybean meal (6.11) [51,52,53].
2.2. Antioxidant Content in Camelina
Camelina oil contains high levels of γ-tocopherol (710 mg/kg. Other tocopherols are α-tocopherol (28.07–41.8 mg/kg) and δ-tocopherol (12.3–20.47 mg/kg).
The total tocopherol content determined in fresh camelina oil amounted from 751 to 687 mg/kg and this amount was higher than in flax oil and rapeseed oil.
A high level of phenolic acid and flavonoids have also been found in camelina. The content of phenolic acid in camelina seed ranges from 2043.6 to 3704.7 mg/kg, in oil from 681.89 to 892.12 mg/L, and in cake from 1148.67 to 1413.76 mg/kg of dry matter.
The content of flavonoids in camelina seed, oil and cake have been found to be from 329.49 to 526.4 mg/kg, 266.01 to 435.32 mg/L, and 37.69 to 73.13 mg/g, respectively. Antioxidants present in camelina prolong the storage time for oil, seeds and cake because they reduce lipid oxidation.
2.3. Antinutritive Compounds in Camelina
The use of camelina feedstock in poultry nutrition is limited by plant secondary metabolites, i.e., glucosinolates, sinapine, phytic acid and condensed tannins that are ascribed to antinutritive compounds found in camelina.
Glucosinolate accumulation in camelina depends on many factors—genotype, climatic conditions, soil type, sulfur content in the soil, and fertilization [14,71]. Therefore, a wide range of glucosinolate content can be found in camelina. The content of glucosinolates in whole seed, as reported by Schuster and Friedt [71], varies from 13.2 to 36.2 μmol/g and the mean value is 24 μmol/g; while Matthäus and Zubr [14] indicated it ranges from 9 to 19 μmol/g. The amount of glucosinolates in camelina cake is from 14.5 to 44.9 μmol/g [14,36,46,72,73,74]. Matthäus and Zubr [14] indicated that glucosinolates are stored in the residue when oil is produced during seed pressing. Research data show that whole seed contains 14.1 μg/mg, seed meal 24.3 μg/mg, and defatted meal as much as 31.8 μg/mg glucosinolates [75].
Woyengo et al. [36] indicate that poultry can tolerate up to 2.0 μmol/g glucosinolates in rapeseed diets, while Tripathi and Mishra [68] increase the tolerance level to 5.6 μmol/g. No sufficient research data can be found to define the effects of camelina-specific glucosinolates and their metabolic products on poultry nutrition.
Since the amount of glycosinolates in camelina varieties varies widely, it indicates a high phenotypic variation, which is a prerequisite for successful selection. Currently, the major breeding objectives for camelina are to increase seed yield, seed oil and protein content, and resistance to abiotic stress, however, varieties with low glucosinolate levels have not been developed.
The content of sinapine varies markedly in camelina plants. The seed analysis of eight different camelina genotypes indicated the range of sinapine content to be from 2.8 to 7.8 mg/g, with an average of 4 mg/g [76]. Meanwhile, the analysis of 30 camelina cultivars from different European localities showed that sinapine concentration in oilseed cake is from 1.7 to 4.2 g/kg [14]. A similar mean sinapine content is found in camelina cake by other researchers, i.e., 2.32 g/kg [72], 2.57 g/kg [74] and 2.79 g/kg [73]. The content of sinapine in camelina is much lower than that found in other Brassicaceae family plants such as rape or mustard (7 and 13 mg/g, respectively) [14]. Feedstuffs with sinapine taste bitter, but as taste buds in birds are poorly developed [79], feed bitterness does not reduce voluntary feed intake in broilers [80]. However, if no more than 10% of camelina cake is used in meat poultry diets, no undesirable sinapine effect will be found, due to a low sinapine concentration.
The content of phytic acid in camelina seed was found to be 21 mg/g, and in cakes from 21.0 to 32.3 mg/g. The amount of phytic acid in camelina is similar to that in sunflower but 1.5 times higher than in rapeseed. Recent studies have shown that phytic acid also has a beneficial effect on health due to its antioxidative properties.
The average content of condensed tannins amounts to 1.1, and ranges from 1.0 to 2.4 mg/g in, respectively, camelina seed and cake. By the data from 12 camelina genotypes, the tannin amount varies from 1.92 to 4.39 g/kg, with an average content of 3.1 g/kg. The tannin content in the other study was found to be from 1.81 to 2.59 g/kg. The amounts of condensed tannins in camelina are relatively low and, therefore, there is either no, or a very insignificant, negative effect on poultry nutrition, as tannins show their toxicity only at over 1% amount in the diet. Meanwhile, even small amount of tannins might have a positive effect health as tannins possess antimicrobial as well as anticarcinogenic and antimutagenic properties.
−1 in the diets of monogastric animals. The US Food and Drug Administration approved inclusion of up to 10% of the weight of the total ration of the diets of beef cattle and poultry. In Canada and the USA, the standard for glucosinolate content in dried canola meal is set at a maximum of 30 μmol/g of dry matter, and in the EU this value should not exceed 20 μmol/g.
3. Influence of Camelina on Growth Performance
Different dietary camelina components and different amounts of their inclusion showed different effects on the growth performance of poultry.
Inclusion of 5% camelina seed in the diet had no effect on the growth performance of chickens, but feeding 10% seed resulted in a lower body weight (BW) and BW gain.
The amount of dietary camelina oil ranging from 2.5 to 4.07–6.91% also did not have any effect on the BW, BW gain, feed intake (FI) or feed conversion ratio (FCR) of chickens. This could be explained by a lower amount of antinutrients in oil in comparison with cake or seed.
The effects of dietary camelina cake on the growth performance of poultry are contradictory. Studies indicate that supplementation of the diets with 8 and 16% camelina cake increased BW and BW gain.
Other researchers who used from 5 to 10 and 24% cake in broiler chicken diets have not found any differences in the growth performance parameters.
Studies with quail (5 and 10% cake) and turkeys (5% cake) also indicated that dietary camelina had no influence on BW, FI and FCR data.
However, other authors indicated that camelina cake had a negative effect on the growth performance of poultry. Ryhänen et al. indicated that 5 and 10% camelina cake inclusion in chicken diets resulted in lower BW, FI (days 1–14) and higher FCR.
Supplementation of quail diets with a higher content of camelina cake (15–20%) resulted in a higher FCR.
Studies also indicated that 15 to 20% camelina cake inclusion in the diets of turkey poults at the first starter phase of up to 4 weeks of age, and 10% cake inclusion in the diets of chickens up to 21 days of age, had a negative effect on BW and FI because chicks and poults do not have a fully developed digestive system and, therefore, lower ability to digest camelina cake.
Many researchers indicated that the reason for poorer growth performance was the presence of antinutrients, the amounts of which were also different in camelina cakes derived from different source materials. It was found that the glucosinolate content in camelina cake could be from 14.5 to 44.9 µmol/g and that might also influence the growth performance results . Toxic glucosinolate transformation products as thiocyanates and oxazolidinethiones disturb the thyroid function, negatively affect growth, fertility and reproduction and reduce feed conversion. Nitriles irritate the gastro-intestinal mucosa and cause local necroses and hepatotoxicity and nephrotoxicity effects.
Other authors found that increasing levels of dietary camelina cake from 3 to 15% in broiler chicken diets reduced the apparent total tract digestibility of dry matter, nitrogen and energy. Pekel et al. reported that camelina cake in broiler diets increased viscosity observed in jejunal digesta and, consequently, reduced utilization of energy and nitrogen.
Moreover, 2.24 to 5.44% fiber was found with 5 to 16% camelina cake inclusion in chicken diets, and in many trials, this amount of fiber was higher than recommended (2.41–2.56%) for Ross cross chicken nutrition management. Non-starch polysaccharides (NSP), which make the basis of fiber, are poorly digested by poultry due to nutrient encapsulating in cell walls. Research shows that when camelina cake amounts to 30%, fiber-degrading enzymes, i.e., carbohydrases, should be used to degrade NSP in order to reduce feed viscosity, improve nutrient utilization and, consequently, poultry growth and FCR.
4. Influence of Camelina on PUFA Composition in Breast, Leg Muscles and Liver
Poultry feeding with a standard compound feed results in a low content of the total n-3 PUFA (from 1.63 to 3.88%) and a comparatively high n-6/n-3 PUFA ratio (from 13.22 to 43.3) in breast and leg muscles. The n-6/n-3 PUFA ratio in the liver is also very high (9.50–12.72). This fatty acid composition is not beneficial to human nutrition.
Therefore, after Nguyen et al. had found linear correlations between the content of PUFA in feeds and in the tissues of monogastric animals offered PUFA-containing diets, researchers conducted numerous trials aiming for the modification of meat fatty acid composition so as to be beneficial to human health. In agreement with the above studies, Kanakri et al. indicated 0.999 Pearson correlations between n-3 PUFA levels in the diets and tissues of meat type chickens.
The diets for chickens and ducks could include different camelina components such as seed (5–10%), cake (2.5–24%) and oil (2.5–6.91%).
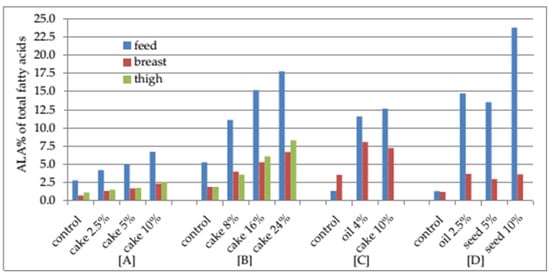
The total n-3 PUFA content in breast and leg muscles could be increased, respectively, from 1.48 to 2.83, and 1.32 to 3.73 times, and in the liver from 1.62 to 3.90 times. These changes have been mostly influenced by the increase in one of the main n-3 PUFA ALA in breast and leg muscles.
The least statistically significant ALA increases, by 1.78 and 1.32 times in, respectively, breast and leg muscles, were found after 2.5% cake inclusion in chicken diets. Meanwhile, supplementation of poultry diets with up to 25% camelina cake resulted in 3.9 and 4.37 times higher ALA content in, respectively, breast and leg muscles. The highest ALA increases in chicken breast and leg muscles by 7.23 and 6.60 times, respectively, were found after rapeseed oil had been replaced by camelina oil. The studies have indicated that the higher the ALA content was in a poultry diet, the higher was its concentration in the muscles.
Long-chain (LC) n-3 fatty acids such as eicosapentaenoic (C20:5n-3; EPA) and docosahexaenoic (C22:6n-3; DHA) are very important in human nutrition. However, the conversion of these acids from ALA in adults is lower than 5% and, therefore, human nutrition should be provided with LC n-3 PUFA. Birds are known for considerably more effective synthesis of LC n-3 PUFA from ALA, and, therefore, LC n-3 PUFA accumulation in poultry tissues is preconditioned by ALA being the precursor of all LC n-3 PUFA.
The increase in eicosatrienoic fatty acid (C20:3n-3; ETE) in chicken and duck muscles and the liver was from 0.08 to 0.37% . Meanwhile, camelina seed, oil and cake feeding resulted in 0.15 to 1.08% higher EPA content in breast muscles though other researchers did not report any changes with 2.5 to 24% cake addition to the diet. This is in agreement with the statement by Rymer and Gyvens that there is at best a very weak relationship between dietary ALA content and tissue EPA content.
The amount of another LC n-3 PUFA, docosapentaenoic fatty acid (C22:5n-3; DPA), increased from 1.67 to 2.35, and 1.9 to 3.77 times in, respectively, leg muscles and the liver. Meanwhile, the addition of small amounts (2.5–10%) of camelina cake did not increase DPA content in breast muscles. However, supplementation of the diets with camelina seed, oil or a larger amount of cake (16–24%) resulted in 1.17 to 1.72 times higher tissue DPA content.
Bioconversion of dietary ALA is clearly indicated by tissue deposition of ETE and DPA which are transitional metabolites of ALA bioconversion to DHA.
The effect of camelina components in the feed on the increase in LC DHA in the muscles is controversial. Feeding with 2.5% oil, or 5 and 10% seed, or 5–25% cake resulted in 1.22 to 4.12 times higher DHA content in breast muscles, whereas DHA increase in leg muscles was from 2.67 to 3.67 times higher than compared with the control group. However, other authors have reported no DHA increase in the muscles when feeding similar amounts of cake (2.5–24%). Rymer and Givens have also indicated that there was no relationship between dietary ALA content and meat DHA content.
Feeding 5 to 24% cake resulted in 1.39 to 3.31 times higher DHA content in the liver. Liver n-3 PUFA profile was distinguished by DHA domination accounting from 29 to 66.44% of total n-3 PUFA, due to the greater ability of poultry liver to convert dietary ALA to DHA.
Studies indicate that the accumulation efficiency of n-3 PUFA was different for the different tissue types. Camelina inclusion in poultry diets showed that the ALA content accounted for 1.35 to 8.07%, and 1.53 to 9.96%, in the fatty acid profile of, respectively, breast and leg muscles. The preferential deposition of ALA in the leg muscles can be explained by the fact that triglycerides are dominant in the intramuscular fat in leg muscles. Meanwhile, DHA tends to be accumulated in phospholipids that are prevalent among breast tissue lipids. Therefore, the DHA content in the breast and leg fatty acid profile accounted for, respectively, 0.66 to 2.35%, and 0.4 to 0.55%.
Sensory analyses of cooked meat indicated that camelina had no influence on the organoleptic quality of meat. No differences were found at evaluation of flavor, tenderness and tastiness. Higher juiciness was reported in one of the studies with camelina oil.
The content of n-6 PUFA in camelina cake is comparatively high, but it is lower than that of n-3 PUFA, and the n-6/n-3 ratio is mostly 0.72. The effect of dietary camelina on n-6 PUFA changes in poultry tissues is not clear. The amount of essential n-6 PUFA LA increased from 1.08 to 2.10 times in chicken muscles after a dietary inclusion of 8 to 24% cake, or 6.91 to 4.07% oil.
However, other authors indicated no changes of LA content in breast muscles after poultry diet supplementation with 2.5–4% seed, 4% oil, or 2.5–25% cake. No changes of LA content in leg muscles were found with 2.5–20% cake in the diet.
It should be noted that LA accumulation in the muscles is lower than that of ALA, and the reason for this could be the assumption that higher dietary ALA causes competition for the same elongation–desaturation enzymes necessary for the synthesis of both n-3 and n-6 LC fatty acids, thus, resulting in a lower LA content.
Studies indicate that one in vivo PUFA metabolism regulating factors is dietary fatty acid composition. Jing et al.have found that the expression of FADS1, FADS2, ELOVL2 and ELOVL5 genes related with lipid metabolism in the liver of broiler chickens was higher when the linoleic/α-linolenic ratio in the diet was lower. Other studies indicated that ALA content in chicken diets also increased the expression of genes related with lipid metabolism (FADS1, FADS2, ELOVL2, ELOVL5) in breast muscles.
Feeding camelina resulted in higher n-3 PUFA amount in muscles and liver and, thus, the n-6/n-3 PUFA ratio decreased and was closer to that suitable for healthy human nutrition.
Inclusion of 5–25% cake or 4% oil in chicken and 15–20% cake in duck diets resulted in 1.57–2.86 n-6/n-3 ratio in poultry breast muscles. Similar changes were found in chicken and duck leg muscles when n-6/n-3 PUFA ratio decreased and ranged from 1.9 to 2.7 with 8 to 25% dietary camelina cake, or 4.07 to 6.91% oil inclusion. In the breast muscles the n-6/n-3 PUFA ratio decreased from 1.48 to 8.35 times, in the thigh muscles from 1.32 to 5.15 times, and in the liver from 1.32–3.29 times, in comparison with control diets. Enrichment of poultry diets with camelina seeds, cake and oil resulted in such fatty acid profile changes in poultry tissues, which allowed the production and supply of healthier poultry to consumers. According to European Commission Nutrition Claims, chicken breast and liver produced with, respectively 16–24% and 10% dietary camelina cake, can be labeled as “high in omega-3 fatty acids”, because the EPA and DHA content was higher than 80 mg/100 g. Meanwhile, the EPA and DHA content in the thigh and breast tissues of chickens fed, respectively, 16–24% and 8% cake, was higher than 40 mg/100 g, and these products could be labeled as “a source of omega-3 fatty acids”. The above results might arouse consumer interest and, consequently, lead to higher consumption of valuable n-3 LC PUFA.
5. Conclusions
Camelina seed and its by-product from oil or biodiesel production, such as cake, can be used for meat poultry feeding because they are a valuable feed rich in crude protein (25–40%), oil (6–37%) and antioxidant substances. The content of crude protein and composition of camelina amino acids is close to that of rapeseed meal.
Camelina is distinguished by a unique fatty acid composition, as ALA accounts for 25.88 to 36.67% of the total fatty acids.
However, camelina also contains antinutrients, especially glucosinolates, that prevent the use of seed and its by-products in poultry nutrition on a larger scale.
Addition of camelina seed, oil, cake to poultry diets results in 1.32 to 7.23 times higher ALA content in chicken muscles, in comparison with conventional chicken diets. Consequently, higher ALA content reduces the n-6/n-3 PUFA ratio from 1.32 to 8.35 times in muscles.
Poultry with a higher n-3 PUFA content is beneficial to consumers in their pursuit of healthier products, as such meat increases the consumption of currently deficient n-3 PUFAs and consequently lowers the risks of cardiovascular diseases. Moreover, the use of camelina cake in poultry diets lowers the cost price of poultry and enhances the sustainability of poultry growing and biofuel production. A wider use of camelina should reduce, at least partly, the dependence on imported non-sustainable soya bean meal, and induce its cultivation worldwide, thus, increasing the crop variety used in agriculture.