TBangladeshe African catfish Clarias gariepinus has been introduced for has substantially increased aquaculture production over the last few decades, and the exotic species share a significant portion of the total fish production. Although exotic species are contributing to aquaculture in Bangladesh due to the scarcity of indigenousproduction, a few of them are causing biodiversity loss and genetic erosion C. batrachusof fnatingerlingve species. The African catfish Clarias gariepinus is a highly carnivorous species and predates small indigenous freshwater fishes when escaping into natural water bodies. However, the government of Bangladesh has banned the farming of C. gariepinus due to the carnivorous nature of this species. The introduction of C. gariepinus caused native biodiversity loss due to its predatory nature.
- catfish
- Claris batrachus
- C. gariepinus
- aquaculture
- Bangladesh
1. Introduction
2. Sequence Analysis of COI and Cytb Genes
2. Sequence Analysis of COI and Cytb Genes
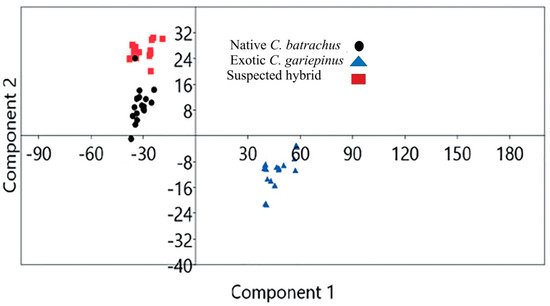
Validation of morphological identification by geometric morphometry: Based on geometric morphometry, the scattered plot of the landmark points showed that native C. batrachus, suspected hybrid and exotic C. gariepinus were morphologically distinguishable (Figure 1). The principal component 1 (PCA 1)_and PCA 2 of landmark points showed that native C. batrachus, suspected hybrid exotic C. gariepinus formed a separate cluster.
3. Sequence Analysis of COI and Cytb Genes
| No | Species Name | 1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|---|
| 1 | ||||||||
| 1.440 | ||||||||
| 1.235 | ||||||||
4. Transition/Transversion Bias
| A | T | C | G | |
|---|---|---|---|---|
| A |
and C. gariepinus; the C. gariepinus from Nigeria, Indonesia and C. batrachus from India, Indonesia, and the Philippines; the sequence of suspected hybrid and native C. batrachusand exotic C. gariepinus. The sequences of the COI gene in native C. batrachus evolved with the same pattern of substitution like the species originating from other counties (
4. The p-values were larger than 0.05 between sequence pairs of reference C. batrachus from the Philippines and Indonesia; the reference C. batrachus from India and Bangladesh relative to the reference from the Philippines and Indonesia; the Bangladeshi sequence of native C. batrachus and C. gariepinus; the C. gariepinus from Nigeria, Indonesia and C. batrachus from India, Indonesia, and the Philippines; the sequence of suspected hybrid and native C. batrachus and exotic C. gariepinus. The sequences of the COI gene in native C. batrachus evolved with the same pattern of substitution like the species originating from other counties (Table 3).
4).| Sl.No | Species Name | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. batrachus | 1 (MG988399) | - | |||||||||||||
| 6.9605% | |||||||||||||||
| 1 | C. batrachus | 6.2622% | 1 (MG988399) | 8.7613% | |||||||||||
| 0.189 | 0.691 | 0.000 * | 0.000 * | 0.318 | 0.146 | 0.000 * | 2 | C. batrachus (Philiphine) | 0.189 | ||||||
| 3 | C. batrachus (Indonesia) | 0.691 |
Table 4). The pairwise genetic distances based on nucleotide sequence divergences are presented in
5). The pairwise genetic distances based on nucleotide sequence divergences are presented inTable 4.
5.| No | Species Name | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | C. batrachus 1 (MG988399) | |||||||||||||||||
| T | 5.9107% | - | 15.3969% | |||||||||||||||
| 2 | ||||||||||||||||||
| 2 | C. batrachus (Philiphine) | 0.194 | 4.5926% | |||||||||||||||
| 0.000 * | 0.000 * | 1.328 | 0.000 * | C. batrachus | 0.000 * | (Philiphine) | 4.931 | 1.258 | C | 5.9107% | 17.1140% | |||||||
| 3 | C. batrachus | 0.000 | ||||||||||||||||
| - | 4.5926% | |||||||||||||||||
| G | 11.2759% | 6.9605% | 6.2622% | - |
5. Homogeneity of Substitution Patterns of COI Sequences
The estimated p values smaller than 0.05 (considered significant and marked with an asterisk) are presented above the diagonal in
The estimated p values smaller than 0.05 (considered significant and marked with an asterisk) are presented above the diagonal inTable 3. The estimated disparity index per site is presented for each sequence pair above the diagonal in
4. The estimated disparity index per site is presented for each sequence pair above the diagonal inTable 3. The p-values were larger than 0.05 between sequence pairs of reference C. batrachus from the Philippines and Indonesia; the reference C. batrachus from India and Bangladesh relative to the reference from the Philippines and Indonesia; the Bangladeshi sequence of native C. batrachus
| (Indonesia) | |||||||||||||||||||||
| 0.108 | 1.000 | 0.000 * | 1.961 | 0.000 * | 0.000 * | 1.909 | |||||||||||||||
| 3 | C. batrachus (Indonesia) | 3.283 | 3.110 | 4 | C. batrachus (India) | 0.000 | 0.000 | 0.000 | 4 | ||||||||||||
| C. batrachus | (India) | 1.000 | 1.000 | 1.000 | 0.900 | 0.000 * | 0.000 * | 0.923 | |||||||||||||
| 4 | C. batrachus (India) | 4.122 | 2.137 | 2.302 | 5 | C. gariepinus (MG988400) | 0.000 | 5 | C. gariepinus | 1.328 | (MG988400) | 1.0001.961 | 0.900 | 0.044 | |||||||
| 0.012 | 0.100 | ||||||||||||||||||||
| 5 | C. gariepinus | ((MG988400) | 1.370 | 1.068 | 0.000 * | ||||||||||||||||
| 0.339 | 5.005 | 4.400 | 6 | C. gariepinus (Indonesia) | 0.318 | 0.000 | 0.000 | 0.000 | 1.370 | ||||||||||||
| 4.635 | 6 | C. gariepinus (Indonesia) | 0.234 | 1.000 | 1.000 | 1.000 | 0.032 | ||||||||||||||
| 6 | 0.000 * | 1.440 | C. gariepinus (Indonesia) | 2.837 | 4.987 | 4.638 | 6.238 | 2.594 | 7 | C. gariepinus (Nigeria) | 0.146 | 0.000 | 0.000 | 0.000 | 1.068 | ||||||
| 7 | C. gariepinus (Nigeria) | 0.332 | 1.000 | 1.000 | 1.000 | 0.000 | |||||||||||||||
| 0.050 | 1.000 | 1.235 | 8 | Suspected hybrid (MG988401) | 0.000 | 1.258 | |||||||||||||||
| 8 | Suspected hybrid (MG988401) | 1.000 | 1.909 | 0.923 | 0.024 | 0.022 | 0.0820.000 | 1.000 | 0.026 | 0.046 |
6. Genetic Distance
The lowest genetic distance was found between suspected hybrid and C. gariepinus (0.295). The highest genetic distance (6.238) was found between C. gariepinus from Indonesia and Bangladeshi C. batrachus . Low levels of genetic divergence (0.295 -0.339) were found among sequenced native Clarias batrachus, suspected hybrid and exotic C. gariepinus ( in Bangladesh, whereas interspecies genetic divergence was found between C. batrachus and C. gariepinus from other countries (
The lowest genetic distance was found between suspected hybrid and C. gariepinus (0.295). The highest genetic distance (6.238) was found between C. gariepinus from Indonesia and Bangladeshi C. batrachus. Low levels of genetic divergence (0.295–0.339) were found among sequenced native Clarias batrachus, suspected hybrid and exotic C. gariepinus (in Bangladesh, whereas interspecies genetic divergence was found between C. batrachus and C. gariepinus| 7 | ||||||||
| C. gariepinus | ||||||||
| (Nigeria) | ||||||||
| 4.055 | 4.636 | 5.095 | 3.147 | 4.025 | 2.315 | |||
| 8 | Suspectred hybrid (MG988401)) | 0.311 | 4.706 | 4.297 | 4.763 | 0.295 | 2.631 | 4.629 |
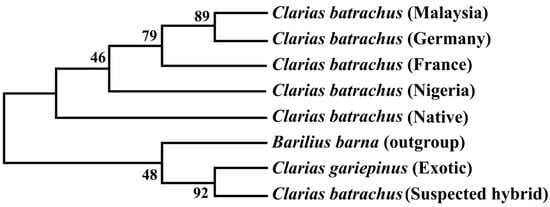
7. Phylogenetic Tree Using COI Nucleotide Sequences by Maximum Likelihood Methods
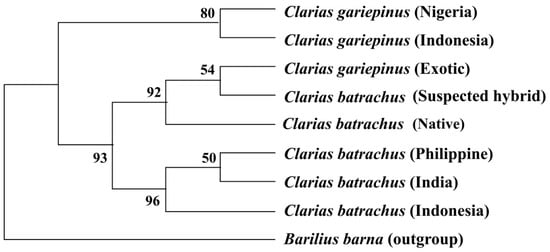
8. Phylogenetic Tree Using Cytb Nucleotide Sequences by Maximum Likelihood Method
The phylogenetic tree based on the Cytb nucleotide sequences was used to validate the phylogenetic tree inferred from the COI nucleotide sequence (
The phylogenetic tree based on the Cytb nucleotide sequences was used to validate the phylogenetic tree inferred from the COI nucleotide sequence (Figure 3). The results showed that sequenced suspected hybrid formed sister taxa with the sequenced C. gariepinus. The native C. batrachus formed a clade with C. batrachus of originated from different countries (Nigeria, France, Germany, and Malaysia) except suspected hybrid and C. gariepinus. The formation of sister clade of suspected hybrid with native C. batrachus and C. gariepinus confirmed the occurrence of hybridization of C. batrachus and C. gariepinus in Bangladesh, which validated the results based on COI gene.
5). The results showed that sequenced suspected hybrid formed sister taxa with the sequenced C. gariepinus. The native C. batrachus formed a clade with C. batrachus of originated from different countries (Nigeria, France, Germany, and Malaysia) except suspected hybrid and C. gariepinus. The formation of sister clade of suspected hybrid with native C. batrachus and C. gariepinus confirmed the occurrence of hybridization of C. batrachus and C. gariepinus in Bangladesh, which validated the results based on COI gene.