The aquaporins (AQPs) are a family of small integral membrane proteins that facilitate the bidirectional transport of water across biological membranes in response to osmotic pressure gradients as well as enabling the transmembrane diffusion of small neutral solutes (such as urea, glycerol, and hydrogen peroxide) and ions. AQPs are expressed throughout the human body.
1. Introduction
Aquaporins (AQPs) have diverse roles in mammals, ranging from fluid homeostasis, glandular secretions, barrier function, immunity and inflammation, cell migration, and angiogenesis to signal transduction and sensation. It is now clear that AQP functions are more complex than simply mediating the passive flow of water across biological membranes
[1]. Understanding their underlying regulatory mechanisms along with the discovery and development of small-molecule AQP inhibitors for use in research and therapeutic development is expected to lead to new insights into the basic biology of and novel treatments for the wide range of AQP-associated disorders.
2. Distribution and Classification of AQPs in the Human Body
The essential role of membrane intrinsic protein channels in the regulation of water transport and homeostasis was discovered in 1986
[2][3][2,3]. The molecular characterization of the archetypal water channel protein, aquaporin-1 (AQP1), launched the research field in the early 1990s with the cloning and characterization of CHIP28 (later renamed AQP1
[4][5][4,5]). Recognizing earlier work, the lens major intrinsic protein (MIP), cloned in 1984
[6], was renamed AQP0 on the basis that it formed ion channels in lipid bilayers
[7] and transported water, albeit at a considerably slower rate than AQP1
[8]. Additional members of the mammalian AQP family were cloned and characterized over the next decade; 13 paralogs have currently been identified in higher mammals, showing differential patterns of tissue and cell expression throughout the body (
Figure 1). Their diverse functions include the transport of water, glycerol, gases, hydrogen peroxide (H
2O
2), ammonia, and ions
[9][10][11][12][9,10,11,12].
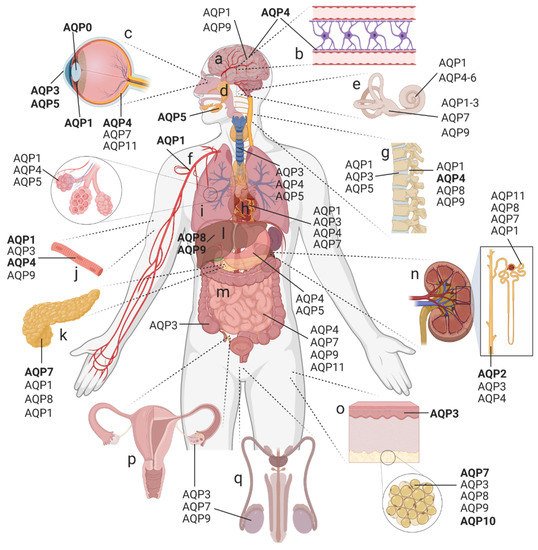
Figure 1. AQP distribution in the human body. Expression of AQP paralogs in the (
a) brain, (
b) blood–brain barrier, (
c) eye, (
d) exocrine glands, (
e) inner ear, (
f) cardiovascular system, (
g) spine, (
h) heart, (
i) respiratory tract (trachea and lung; inset showing alveoli), (
j) skeletal muscle, (
k) pancreas, (
l) liver, (
m) gastrointestinal tract, (
n) kidney, (
o) skin (inset showing adipose tissue), and (
p) female as well as (
q) male reproductive tracts. This summary is not comprehensive; minor AQP subtypes are omitted for clarity.
Bold text is used to highlight the major AQPs studied in the selected tissues
. [12]Created with BioRender.com; adapted from Day et al., 2014 [12].
The traditional categorization of aquaporins (AQPs) into orthodox (water-selective) and glycerol-permeable subtypes, with a third poorly understood ‘super’ or ‘subcellular’ group, no longer encompasses the expanding appreciation of AQPs as multi-functional channels. A broad repertoire of permeable substrates is evident not only in mammalian classes but in AQP classes across the kingdoms of life
[13]. In the traditional scheme, orthodox mammalian AQPs comprise AQP0, 1, 2, 4, 5, and 6. This group includes the classic water channels, but interestingly also includes all the mammalian AQPs known thus far to have dual roles as water and ion channels (which are AQP0, 1, and 6). No ion channel function has yet been detected for AQP4 and AQP5
[14], though it is possible that the key stimuli remain to be identified, a concept to be considered for any AQPs that appear to be nonfunctional in experimental assays. When a multiple-sequence alignment analysis is run for human AQPs combined with known non-mammalian AQP ion channels, AQP8 falls on a distant branch of the orthodox group alongside the soybean AQP channel nodulin 26, which conducts water, glycerol, ammonia, and ions
[1][15][16][1,15,16]. Although AQP8 in other alignments has been assigned to the ‘super-aquaporin’ group with AQP11 and AQP12
[17], its capacity for conducting ammonia
[18] presents an interesting similarity with nodulin 26.
AQP0 was the first mammalian water channel suggested to mediate an ion conductance, in addition to its function as a water channel
[19][20][19,20]. AQP1′s function as a gated ion channel was proposed in 1996
[21], and has since been refined to clarify that AQP1 activation occurs via the direct binding of cyclic guanosine monophosphate (cGMP)
[22][23][24][22,23,24]. The subsequent identification of other members of the AQP family as dual water and ion channels has added the mammalian water channel AQP6, the insect water channel
Drosophila big brain (DmBIB), and plant membrane intrinsic proteins such as the
Arabidopsis plasma membrane intrinsic protein (PIP2;1) to the list
[14][25][26][14,25,26].
The aquaglyceroporin group (AQP3, 7, 9, and 10) captures well-characterized glycerol-permeable channels. The most-recently cloned AQPs from higher mammalian orders include AQP11 and 12
[27]. Initial tests found no water permeability for AQP11 expressed in
Xenopus oocytes
[28]; however, reconstituted in membrane vesicles, AQP11 showed a low but detectable water permeability that was sensitive to mercury
[29]. At positions which correspond to the conserved arginine aromatic barrier in most AQPs, residues in AQP11 and 12 are uniquely hydrophobic, lacking both the aromatic and basic elements of this functionally important site. Although this should not be compatible with the transport of water, alternative barrier sites have been proposed
[30].
3. Structural Biology of the AQP Family
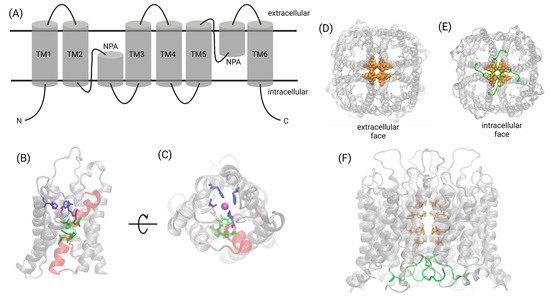
Owing to the relative ease of crystallizing AQP proteins, the structural biology of the transmembrane domains of the AQP family is well-established. A conserved signature fold consists of six transmembrane helices. Two re-entrant helix-forming loops stack one on top of the other, with the family’s signature asparagine–proline–alanine (NPA) motif present in both helices at their interface (
Figure 2A). In contrast to the transmembrane domains, little is known about the structures of the intracellular amino- and carboxy-termini of AQP proteins. These less-ordered regions are usually removed to facilitate crystallization. Complex formation by calmodulin (CaM) binding to the unstructured C-terminus of AQP4 causes it to adopt an α-helical conformation
[31]. The short C-terminal domain of AQP2 exemplifies this flexibility; the C-termini in each monomer adopted four different conformations within the tetrameric unit cell in an X-ray crystal structure
[32].
Figure 2. Structural biology of the AQP family. (A) The signature fold of the AQP family consists of six transmembrane helices and two helix-forming re-entrant loops containing the signature NPA motif. (B,C) Water transport and selectivity is facilitated by the NPA motifs (green) found at the interface of the two helical re-entrant loops (red) and the aromatic/arginine selectivity filter (blue). Water molecules (a single water oxygen at the selectivity filter is indicated by a purple sphere) traverse the pore in single-file. (D–F) The central pore formed at the fourfold axis of AQP1 contains two rings of bulky hydrophobic residues (orange) that prevent pore hydration in the absence of a cGMP signal. cGMP binding at loop D (green) activates the ion channel. Created with Biorender.com.
Following a pattern observed for cyclic-nucleotide-gated channels and potassium channels
[33], AQP tetramers have a central pore at the four-fold axis of symmetry
[34][35][36][34,35,36], which remains incompletely characterized for most AQP classes. Pharmacological and functional analyses have shown in AQP1 that ion and water transport occurs through independent parallel pathways
[11].
Despite the ubiquity of tetramerization in the AQP family, the water pores reside and function within monomers, as evidenced by AQP structures in which permeable substrates have been co-crystallized in the intrasubunit pore in single-file
[34][37][38][39][34,37,38,39]. AQP1 fusion proteins, linking one functional and one non-water-conducting monomer into dimers, were used to demonstrate that each subunit contains an independent water pore pathway
[40]. In human AQP4, mutations in the loop D domain were shown to reduce oligomerization, impairing membrane trafficking and its responsiveness to osmotic stimuli
[41].
The molecular mechanisms of ion permeation and gating differ between AQP classes. In AQP1, cGMP-activated cation currents are thought to flow through the central pore
[36]. Molecular dynamic simulations and mutagenesis revealed that cGMP interacts with an arginine-rich region in loop D, causing loop displacement and conformational changes, and widening the central pore to enable hydration and then ion permeation
[24][42][43][24,42,43]. In the closed central pore, hydrophobic barriers restrict ion permeation but could allow the permeation of gases, such as CO
2 [44]. The loop D domain of AQP1, modeled as the gate for cGMP-dependent ion-channel-opening, might interact with the C-terminus, which modulates activation rates
[22][23][42][43][45][22,23,42,43,45].
Amino acid sequence similarities between the AQP1 C-terminus and other known cGMP-interacting proteins fit a proposed modulatory role for the C-terminal domain
[45]. The activation of ion conductance is impaired in AQP1 channels with truncated or mutated C-termini
[46]. In contrast, in AQP6 the Hg
2+-inducible permeation pathways for anions appear to reside in the intrasubunit (monomeric) pores, based on the Hg
2+-sensitive cysteine locations and the lack of allostery in Hg
2+-induced activation
[47]. AQP6 channel properties are affected by the conformational flexibility of transmembrane helices involving conserved glycines at the crossing point of transmembrane domains TM2 and TM5; the mutation of a glutamine uniquely present in rat AQP6 into a glycine typical of most AQPs (N60G) abolished anion conduction while increasing water permeability
[48]. The molecular basis of ion transport through AQP0 remains unexplored.
4. AQP Permeabilities: An Expanding Repertoire
In contrast to initial expectations, AQPs have turned out to be more than simple water channels, and display properties of a diverse multifunctional protein family that is still not fully characterized. The spectrum of roles now recognized for AQPs include water, glycerol, urea, ammonia, nitric oxide, and H
2O
2 transport; ion conductance; direct mediation of cell–cell adhesion; and regulation of the plasma membrane abundance of other membrane proteins (
Table 1). Emerging permeability properties do not segregate neatly into the traditional orthodox and aquaglyceroporin classification scheme. Classes of AQPs that facilitate H
2O
2 transport, described as peroxiporins, include AQP1
[49], AQP3
[50], AQP5
[51], AQP8
[50][52][53][50,52,53], and AQP9
[54], although earlier work on AQP1 did not find peroxiporin activity
[55]. AQP8 is expressed in the inner membrane of mitochondria and involved in H
2O
2 transport linked with the accumulation of reactive oxygen species (ROS)
[52], revealing an unexpected breadth of physiologically important roles for AQPs across phyla
[13][56][13,56] and highlighting many gaps that are yet to be addressed. Recent perspectives have provided comprehensive synopses of AQP-related diseases
[57][58][59][60][61][62][63][64][65][66][67][68][57,58,59,60,61,62,63,64,65,66,67,68].
Table 1. AQP classification with permeant substrates and main sites of expression. Chromosome location and water permeability data adapted from
[69]. Asterisks (*) highlight permeability results that are controversial or not yet accepted as readily reproducible.
|
Aquaporin
|
Chromosome
|
Water Permeability (Pf)
[×10−14 cm3 s−1]
|
Permeability to Molecules Other Than Water
|
Main Expression Sites
|
|
Orthodox (classical) AQPs
|
|
AQP0
|
12q13
|
0.25
|
Ions [19][20][19,20]
|
Eye lens
|
|
AQP1
|
7p14
|
6.0
|
Monovalent cations [24][36][42][24,36,42], nitric oxide [70], H2O2 [49][55][49,55], and glycerol * [71]
|
Central nervous system (CNS), inner ear, eye, kidney, endothelium, lung, skeletal muscle, cartilage, and erythrocytes
|
|
AQP2
|
12q13
|
3.3
|
None known
|
Kidney, inner ear, and reproductive tract
|
|
AQP4
|
18q22
|
24
|
Nitric oxide [72]
|
CNS, inner ear, retina, kidney, gastrointestinal tract (GIT), lung, and skeletal muscle
|
|
AQP5
|
12q13
|
5.0
|
H2O2 [51]
|
Secretory glands, inner ear, eye, kidney, GIT, and lung
|
|
AQP6
|
12q13
|
Low; no quantitative data
|
Ammonia [73], glycerol, urea [74], nitrate [75], and anions (NO3−, Cl−) [76]
|
Inner ear, kidney
|
|
AQP8
|
16p12
|
No quantitative data
|
Urea, ammonia, and H2O2 [77]
|
Liver, kidney, adipose tissue, pancreas, GIT, and reproductive tract
|
|
Aquaglyceroporins
|
|
AQP3
|
9p13
|
2.1
|
Glycerol [78], H2O2 [9], urea * [78], and ammonia [79]
|
Skin, inner ear, eye, adipose tissue, kidney, GIT, heart, lung, reproductive tract, and cartilage
|
|
AQP7
|
9p13
|
No quantitative data
|
Arsenite [80], glyerol and urea [81], and ammonia [82]
|
Adipose tissue, pancreas, liver, kidney, inner ear, GIT, heart, reproductive tract
|
|
AQP9
|
15q22
|
No quantitative data
|
Arsenite [80], carbamides, polyols, purines, pyrimidines [83], ketone bodies [84], lactate [85], ammonia [86], glycerol, urea [83][87][88][83,87,88], and H2O2 [54]
|
Liver, adipose tissue, CNS (unclear for humans), inner ear, and reproductive tract
|
|
AQP10
|
1q21
|
No quantitative data
|
Glycerol [89]
|
Adipose tissue and reproductive tract
|
|
Unorthodox AQPs/S-aquaporins
|
|
AQP11
|
11q13
|
~2
|
Glycerol [29]90][91][29[,90,91]
|
Retina, kidney, GIT, and reproductive tract
|
|
AQP12
|
2q37
|
No quantitative data
|
Unknown
|
Pancreas
|
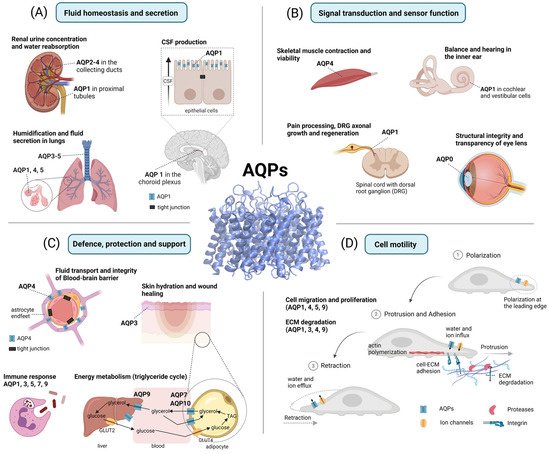
The diverse array of AQP functions in human physiology and pathophysiology has been divided into four overarching themes. These are: fluid homeostasis and secretion; signal transduction and sensory functions; defense and metabolism; and motility and cancer (Figure 3).
Figure 3. Functional roles of AQPs. (A) Fluid homeostasis and secretion: In the kidney, AQP1 regulates water reabsorption in the proximal tubules, while AQP2–4 are involved in urine concentration. In the central nervous system (CNS), AQP1 is involved in cerebrospinal fluid (CSF) production in the choroid plexus. In the lungs, AQPs facilitate transendothelial and transepithelial water flow. (B) Signal transduction and sensor function: AQP4 is involved in skeletal muscle contraction and viability. In the spinal cord, AQP1 is thought to contribute to pain processing and promote axonal growth as well as the regeneration of dorsal root ganglia (DRG). In the inner ear, AQPs are involved in balance and hearing. In the eye, AQP0 facilitates the structural integrity and transparency of the lens. (C) Defense, protection, and support: AQP4 is involved in blood–brain barrier (BBB) integrity, astrocyte plasticity, glial scar formation, and cerebral waste clearance. AQP3 supports skin hydration and wound healing. AQP1, 3, 5, 7, and 9 are involved in immune cell activation and pathogen elimination (phagocytosis). AQP7, 9, and 10 are involved in the glycerol transport that supports energy metabolism. (D) Cell motility: AQP1, 4, 5, and 9 are polarized at the leading edge of migrating cells and are thought to promote the cellular migration stages of polarization, protrusion, adhesion, and retraction. Additionally, AQP1, 3, 4, and 9 are assumed to enhance the degradation of the extracellular matrix (ECM). Created with BioRender.com.