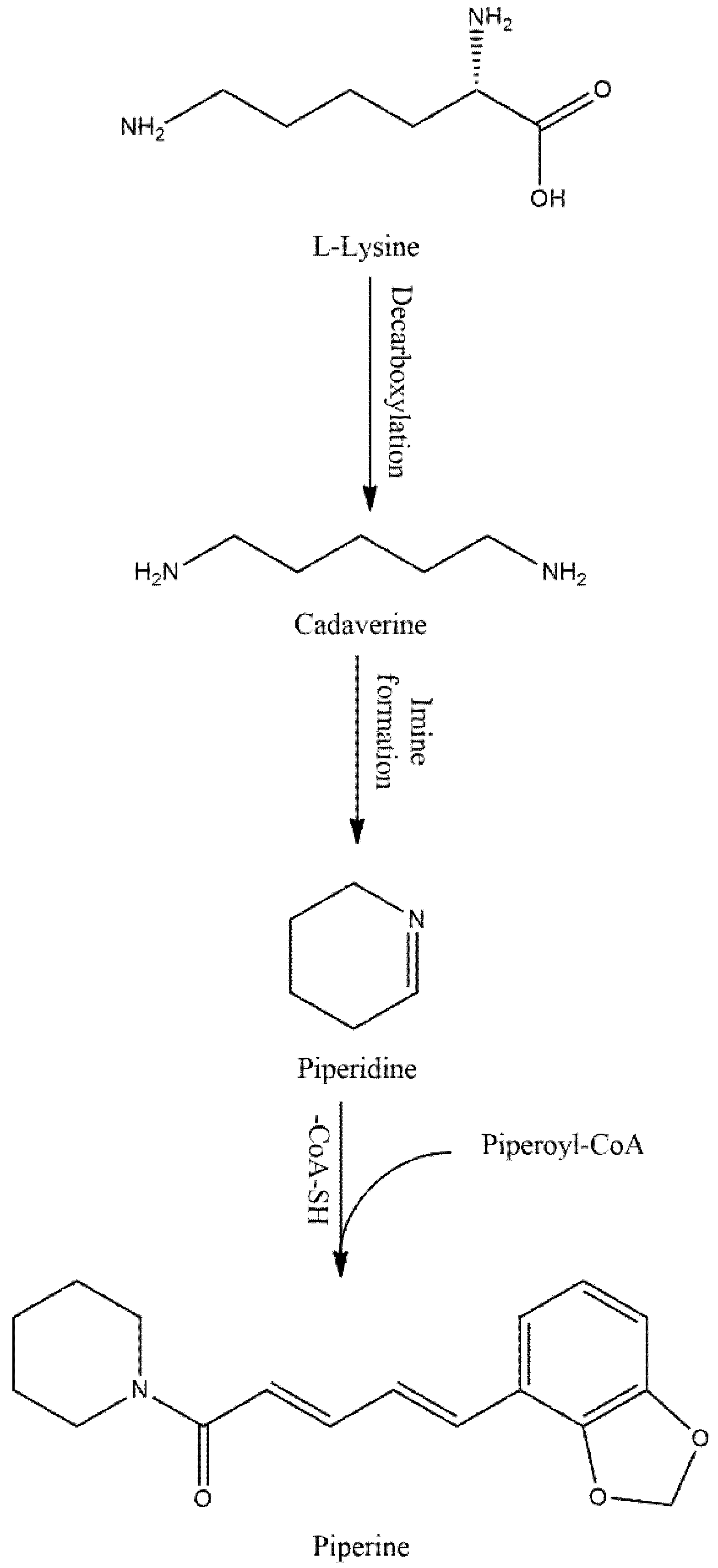
Piperine (PIP) is an active alkaloid of black and long peppers. An increasing amount of evidence is suggesting that PIP and its metabolite’s could be a potential therapeutic to intervene different disease conditions including chronic inflammation, cardiac and hepatic diseases, neurodegenerative diseases, and cancer. In addition, the omnipresence of PIP in food and beverages made this compound an important investigational material. It has now become essential to understand PIP pharmacology and toxicology to determine its merits and demerits, especially its effect on the central nervous system (CNS). Although several earlier reports documented that PIP has poor pharmacokinetic properties, such as absorption, bioavailability, and blood–brain barrier permeability. However, its interaction with metabolic enzyme cytochrome P450 superfamily and competitive hydrophobic interaction at Monoamine oxide B (MAO-B) active site have made PIP both a xenobiotics bioenhancer and a potential MAO-B inhibitor. Moreover, recent advancements in pharmaceutical technology have overcome several of PIP’s limitations, including bioavailability and blood–brain barrier permeability, even at low doses. Contrarily, the structure activity relationship (SAR) study of PIP suggesting that its several metabolites are reactive and plausibly responsible for acute toxicity or have pharmacological potentiality.
- piperine
- biosynthesis
- metabolites
- Alzheimer’s disease
- Parkinson’s disease
1. Introduction
| Sources | Plant Parts | Estimated Amount (%) |
|---|---|---|
| Piper nigrum [2] | Fruit | 1.7–7.4 |
| Piper longum [3] | Spike and root | 5.9 |
| Fruit | 0.03 | |
| Piper chaba [3][4] | Fruit | 0.95–1.32 |
| Piper guineense [5] | Fruit | 0.23–1.1 |
| Piper sarmentosum [6] | Root | 0.20 |
| Stem | 1.59 | |
| Leaf | 0.104 | |
| Fruit | 2.75 |
2. PIP in Neurological and Psychiatric Diseases
2.1. AD
2.2. PD
2.3. Huntington’s Disease
2.4. Epilepsy
2.5. Other Neurological Diseases
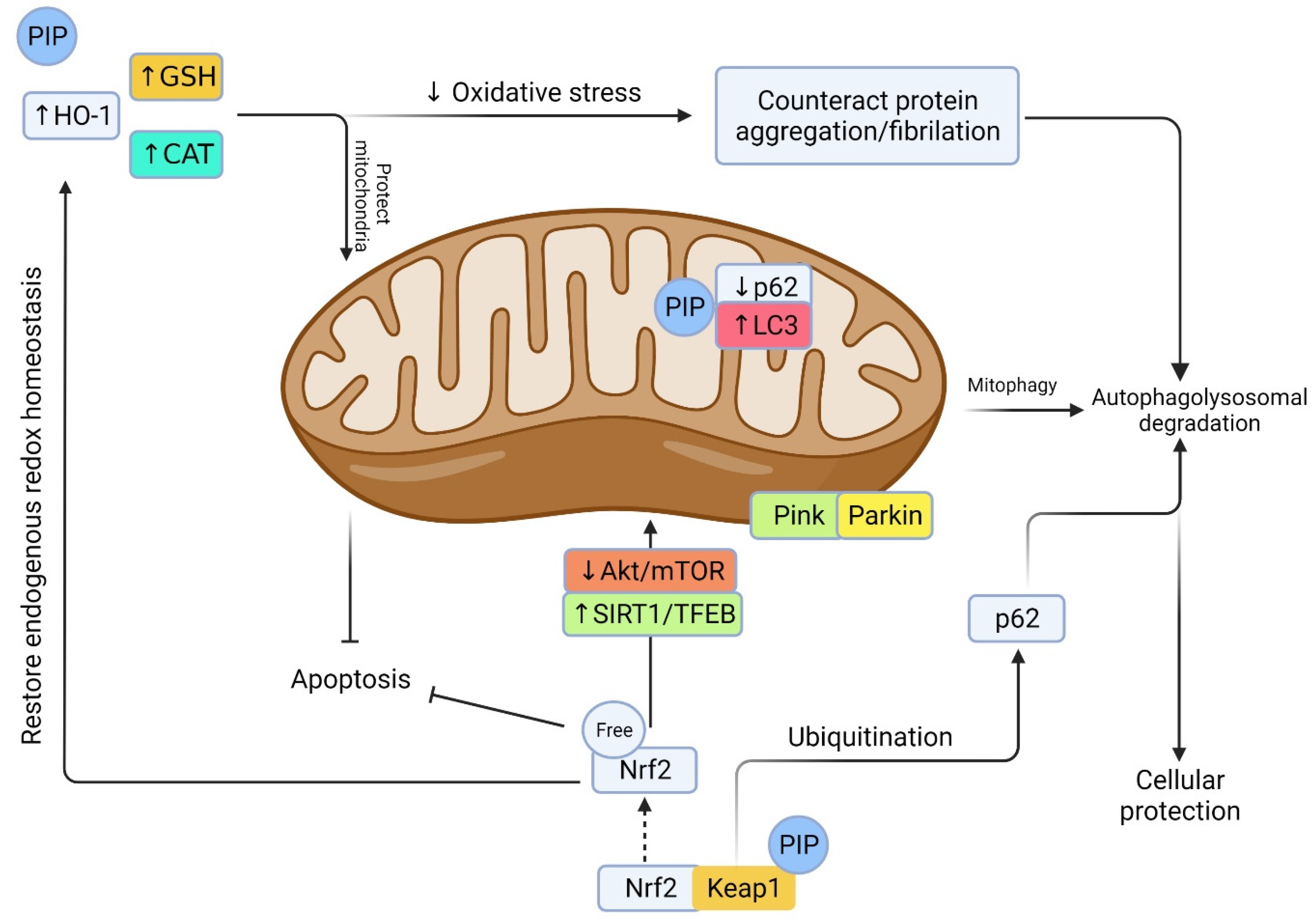
3. Therapeutic Index and Future Perspective
Abbreviations
References
- Khan, A.; Jahan, S.; Imtiyaz, Z.; Alshahrani, S.; Antar Makeen, H.; Mohammed Alshehri, B.; Kumar, A.; Arafah, A.; Rehman, M.U. Neuroprotection: Targeting Multiple Pathways by Naturally Occurring Phytochemicals. Biomedicines 2020, 8, 284.
- Mohapatra, M.; Basak, U. Evaluation of Piperine Content from Roots of Piper Longum Linn., Originated from Different Sources with Comparison of Zonal Variation in Odisha, India. Int. J. Pharma Res. Rev. 2015, 4, 1–8.
- Rameshkumar, K.B.; Aravind, A.P.A.; Mathew, P.J. Comparative Phytochemical Evaluation and Antioxidant Assay of Piper longum L. and Piper chaba Hunter Used in Indian Traditional Systems of Medicine. J. Herbs Spices Med. Plants 2011, 17, 351–360.
- Khan, M. Comparative Physicochemical Evaluation of Fruits and Anti depressant Potential of volatile oils of fruits of Local Piper Species. Orient. J. Chem. 2015, 31, 541–545.
- Juliani, H.R.; Koroch, A.R.; Giordano, L.; Amekuse, L.; Koffa, S.; Asante-Dartey, J.; Simon, J.E. Piper guineense (Piperaceae): Chemistry, Traditional Uses, and Functional Properties of West African Black Pepper. In African Natural Plant Products Volume II: Discoveries and Challenges in Chemistry, Health, and Nutrition; American Chemical Society: Washington, DC, USA, 2013; Volume 1127, pp. 33–48.
- Hussain, K.; Ismail, Z.; Sadikun, A.; Ibrahim, P. Antioxidant, anti-TB activities, phenolic and amide contents of standardised extracts of Piper sarmentosum Roxb. Nat. Prod. Res. 2009, 23, 238–249.
- Nazifi, M.; Oryan, S.; Esfahani, D.E.; Ashrafpoor, M. The functional effects of piperine and piperine plus donepezil on hippocampal synaptic plasticity impairment in rat model of Alzheimer’s disease. Life Sci. 2021, 265, 118802.
- Wang, C.; Cai, Z.; Wang, W.; Wei, M.; Kou, D.; Li, T.; Yang, Z.; Guo, H.; Le, W.; Li, S. Piperine attenuates cognitive impairment in an experimental mouse model of sporadic Alzheimer’s disease. J. Nutr. Biochem. 2019, 70, 147–155.
- Khalili-Fomeshi, M.; Azizi, M.G.; Esmaeili, M.R.; Gol, M.; Kazemi, S.; Ashrafpour, M.; Moghadamnia, A.A.; Hosseinzadeh, S. Piperine restores streptozotocin-induced cognitive impairments: Insights into oxidative balance in cerebrospinal fluid and hippocampus. Behav. Brain Res. 2018, 337, 131–138.
- Hsieh, T.Y.; Chang, Y.; Wang, S.J. Piperine-mediated suppression of voltage-dependent Ca(2+) influx and glutamate release in rat hippocampal nerve terminals involves 5HT(1A) receptors and G protein βγ activation. Food Funct. 2019, 10, 2720–2728.
- Andreza, S.F.; Matheus, G.d.O.; Giuliana, M.V.V.S.; Carlos, H.T.d.P.d.S.; Vinícius, B.d.S.; Carlton, A.T.; Gilberto, L.B.d.A. The Natural Alkaloid Piperine and its Acid and Ester Synthetic Derivatives are Acetylcholinesterase Inhibitors. Curr. Phys. Chem. 2015, 5, 294–300.
- Ravelli, K.G.; Rosário, B.D.; Camarini, R.; Hernandes, M.S.; Britto, L.R. Intracerebroventricular Streptozotocin as a Model of Alzheimer’s Disease: Neurochemical and Behavioral Characterization in Mice. Neurotox. Res. 2017, 31, 327–333.
- Murata, K.; Matsumura, S.; Yoshioka, Y.; Ueno, Y.; Matsuda, H. Screening of β-secretase and acetylcholinesterase inhibitors from plant resources. J. Nat. Med. 2015, 69, 123–129.
- Kumar, S.; Chowdhury, S.; Razdan, A.; Kumari, D.; Purty, R.S.; Ram, H.; Kumar, P.; Nayak, P.; Shukla, S.D. Downregulation of Candidate Gene Expression and Neuroprotection by Piperine in Streptozotocin-Induced Hyperglycemia and Memory Impairment in Rats. Front. Pharmacol. 2020, 11, 595471.
- Head, E.; Murphey, H.L.; Dowling, A.L.; McCarty, K.L.; Bethel, S.R.; Nitz, J.A.; Pleiss, M.; Vanrooyen, J.; Grossheim, M.; Smiley, J.R.; et al. A combination cocktail improves spatial attention in a canine model of human aging and Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2012, 32, 1029–1042.
- Elnaggar, Y.S.; Etman, S.M.; Abdelmonsif, D.A.; Abdallah, O.Y. Intranasal Piperine-Loaded Chitosan Nanoparticles as Brain-Targeted Therapy in Alzheimer’s Disease: Optimization, Biological Efficacy, and Potential Toxicity. J. Pharm. Sci. 2015, 104, 3544–3556.
- Yusuf, M.; Khan, M.; Khan, R.A.; Ahmed, B. Preparation, characterization, in vivo and biochemical evaluation of brain targeted Piperine solid lipid nanoparticles in an experimentally induced Alzheimer’s disease model. J. Drug Target. 2013, 21, 300–311.
- Etman, S.M.; Elnaggar, Y.S.R.; Abdelmonsif, D.A.; Abdallah, O.Y. Oral Brain-Targeted Microemulsion for Enhanced Piperine Delivery in Alzheimer’s Disease Therapy: In Vitro Appraisal, In Vivo Activity, and Nanotoxicity. AAPS PharmSciTech 2018, 19, 3698–3711.
- Gad, S.C.; Cassidy, C.D.; Aubert, N.; Spainhour, B.; Robbe, H. Nonclinical vehicle use in studies by multiple routes in multiple species. Int. J. Toxicol. 2006, 25, 499–521.
- Hong, R.; Li, X. Discovery of monoamine oxidase inhibitors by medicinal chemistry approaches. MedChemComm 2019, 10, 10–25.
- Al-Baghdadi, O.B.; Prater, N.I.; Van der Schyf, C.J.; Geldenhuys, W.J. Inhibition of monoamine oxidase by derivatives of piperine, an alkaloid from the pepper plant Piper nigrum, for possible use in Parkinson’s disease. Bioorg. Med. Chem. Lett. 2012, 22, 7183–7188.
- Shrivastava, P.; Vaibhav, K.; Tabassum, R.; Khan, A.; Ishrat, T.; Khan, M.M.; Ahmad, A.; Islam, F.; Safhi, M.M.; Islam, F. Anti-apoptotic and anti-inflammatory effect of Piperine on 6-OHDA induced Parkinson’s rat model. J. Nutr. Biochem. 2013, 24, 680–687.
- Liu, J.; Chen, M.; Wang, X.; Wang, Y.; Duan, C.; Gao, G.; Lu, L.; Wu, X.; Wang, X.; Yang, H. Piperine induces autophagy by enhancing protein phosphotase 2A activity in a rotenone-induced Parkinson’s disease model. Oncotarget 2016, 7, 60823–60843.
- Wang, H.; Liu, J.; Gao, G.; Wu, X.; Wang, X.; Yang, H. Protection effect of piperine and piperlonguminine from Piper longum L. alkaloids against rotenone-induced neuronal injury. Brain Res. 2016, 1639, 214–227.
- Wang, L.; Cai, X.; Shi, M.; Xue, L.; Kuang, S.; Xu, R.; Qi, W.; Li, Y.; Ma, X.; Zhang, R.; et al. Identification and optimization of piperine analogues as neuroprotective agents for the treatment of Parkinson’s disease via the activation of Nrf2/keap1 pathway. Eur. J. Med. Chem. 2020, 199, 112385.
- Li, R.; Lu, Y.; Zhang, Q.; Liu, W.; Yang, R.; Jiao, J.; Liu, J.; Gao, G.; Yang, H. Piperine promotes autophagy flux by P2RX4 activation in SNCA/α-synuclein-induced Parkinson disease model. Autophagy 2021, 1–17.
- Huang, L.; Zhong, X.; Luo, Q.; Zhang, Q.; Deng, M. Autophagic activity of piperine on small intestine in dementia model mice with Parkinson’s disease. Zhongguo Zhong Yao Za Zhi Zhongguo Zhongyao Zazhi China J. Chin. Mater. Med. 2020, 45, 5238–5247.
- Salman, M.; Tabassum, H.; Parvez, S. Piperine mitigates behavioral impairments and provides neuroprotection against 3-nitropropinoic acid-induced Huntington disease-like symptoms. Nutr. Neurosci. 2020, 1–10.
- Pal, A.; Nayak, S.; Sahu, P.K.; Swain, T. Piperine protects epilepsy associated depression: A study on role of monoamines. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 1288–1295.
- da Cruz, G.M.; Felipe, C.F.; Scorza, F.A.; da Costa, M.A.; Tavares, A.F.; Menezes, M.L.; de Andrade, G.M.; Leal, L.K.; Brito, G.A.; da Graça Naffah-Mazzacoratti, M.; et al. Piperine decreases pilocarpine-induced convulsions by GABAergic mechanisms. Pharmacol. Biochem. Behav. 2013, 104, 144–153.
- Mishra, A.; Punia, J.K.; Bladen, C.; Zamponi, G.W.; Goel, R.K. Anticonvulsant mechanisms of piperine, a piperidine alkaloid. Channels 2015, 9, 317–323.
- Chen, C.Y.; Li, W.; Qu, K.P.; Chen, C.R. Piperine exerts anti-seizure effects via the TRPV1 receptor in mice. Eur. J. Pharmacol. 2013, 714, 288–294.
- Dong, Y.; Yin, Y.; Vu, S.; Yang, F.; Yarov-Yarovoy, V.; Tian, Y.; Zheng, J. A distinct structural mechanism underlies TRPV1 activation by piperine. Biochem. Biophys. Res. Commun. 2019, 516, 365–372.
- Ren, T.; Yang, M.; Xiao, M.; Zhu, J.; Xie, W.; Zuo, Z. Time-dependent inhibition of carbamazepine metabolism by piperine in anti-epileptic treatment. Life Sci. 2019, 218, 314–323.
- Pattanaik, S.; Hota, D.; Prabhakar, S.; Kharbanda, P.; Pandhi, P. Pharmacokinetic interaction of single dose of piperine with steady-state carbamazepine in epilepsy patients. Phytother. Res. PTR 2009, 23, 1281–1286.
- Verma, A.K.; Khan, E.; Mishra, S.K.; Jain, N.; Kumar, A. Piperine Modulates Protein Mediated Toxicity in Fragile X-Associated Tremor/Ataxia Syndrome through Interacting Expanded CGG Repeat (r(CGG)(exp)) RNA. ACS Chem. Neurosci. 2019, 10, 3778–3788.
- Mao, Q.Q.; Huang, Z.; Ip, S.P.; Xian, Y.F.; Che, C.T. Role of 5-HT(1A) and 5-HT(1B) receptors in the antidepressant-like effect of piperine in the forced swim test. Neurosci. Lett. 2011, 504, 181–184.
- Mao, Q.Q.; Huang, Z.; Zhong, X.M.; Xian, Y.F.; Ip, S.P. Brain-derived neurotrophic factor signalling mediates the antidepressant-like effect of piperine in chronically stressed mice. Behav. Brain Res. 2014, 261, 140–145.
- Khom, S.; Strommer, B.; Schöffmann, A.; Hintersteiner, J.; Baburin, I.; Erker, T.; Schwarz, T.; Schwarzer, C.; Zaugg, J.; Hamburger, M.; et al. GABAA receptor modulation by piperine and a non-TRPV1 activating derivative. Biochem. Pharmacol. 2013, 85, 1827–1836.
- Wang-Sheng, C.; Jie, A.; Jian-Jun, L.; Lan, H.; Zeng-Bao, X.; Chang-Qing, L. Piperine attenuates lipopolysaccharide (LPS)-induced inflammatory responses in BV2 microglia. Int. Immunopharmacol. 2017, 42, 44–48.
- Vaibhav, K.; Shrivastava, P.; Javed, H.; Khan, A.; Ahmed, M.E.; Tabassum, R.; Khan, M.M.; Khuwaja, G.; Islam, F.; Siddiqui, M.S.; et al. Piperine suppresses cerebral ischemia-reperfusion-induced inflammation through the repression of COX-2, NOS-2, and NF-κB in middle cerebral artery occlusion rat model. Mol. Cell. Biochem. 2012, 367, 73–84.
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin–proteasome system upon ubiquitinated protein degradation. Cell. Mol. Biol. Lett. 2016, 21, 29.
- Wattanathorn, J.; Chonpathompikunlert, P.; Muchimapura, S.; Priprem, A.; Tankamnerdthai, O. Piperine, the potential functional food for mood and cognitive disorders. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2008, 46, 3106–3110.
- Li, S.; Wang, C.; Li, W.; Koike, K.; Nikaido, T.; Wang, M.W. Antidepressant-like effects of piperine and its derivative, antiepilepsirine. J. Asian Nat. Prod. Res. 2007, 9, 421–430.
- Gilhotra, N.; Dhingra, D. Possible involvement of GABAergic and nitriergic systems for antianxiety-like activity of piperine in unstressed and stressed mice. Pharmacol. Rep. 2014, 66, 885–891.
- Bajad, S.; Coumar, M.; Khajuria, R.; Suri, O.P.; Bedi, K.L. Characterization of a new rat urinary metabolite of piperine by LC/NMR/MS studies. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2003, 19, 413–421.
- Shwaireb, M.H.; Wrba, H.; el-Mofty, M.M.; Dutter, A. Carcinogenesis induced by black pepper (Piper nigrum) and modulated by vitamin A. Exp. Pathol. 1990, 40, 233–238.
- Unchern, S.; Saito, H.; Nishiyama, N. Death of cerebellar granule neurons induced by piperine is distinct from that induced by low potassium medium. Neurochem. Res. 1998, 23, 97–102.
- Chu, C.Y.; Chang, J.P.; Wang, C.J. Modulatory effect of piperine on benzopyrene cytotoxicity and DNA adduct formation in V-79 lung fibroblast cells. Food Chem. Toxicol. 1994, 32, 373–377.
- Dogra, R.K.; Khanna, S.; Shanker, R. Immunotoxicological effects of piperine in mice. Toxicology 2004, 196, 229–236.
- EFSA Panel on Food Contact Materials, Enzymes, Flavourings and Processing Aids (CEF). Scientific Opinion on Flavouring Group Evaluation 86, Revision 2 (FGE.86Rev2): Consideration of aliphatic and arylalkyl amines and amides evaluated by JECFA (65th meeting). EFSA J. 2015, 13, 3998.
- Haq, I.-U.; Imran, M.; Nadeem, M.; Tufail, T.; Gondal, T.A.; Mubarak, M.S. Piperine: A review of its biological effects. Phytother. Res. 2021, 35, 680–700.
- Elnaggar, Y.S.; Etman, S.M.; Abdelmonsif, D.A.; Abdallah, O.Y. Novel piperine-loaded Tween-integrated monoolein cubosomes as brain-targeted oral nanomedicine in Alzheimer’s disease: Pharmaceutical, biological, and toxicological studies. Int. J. Nanomed. 2015, 10, 5459–5473.
- Moradi, S.Z.; Momtaz, S.; Bayrami, Z.; Farzaei, M.H.; Abdollahi, M. Nanoformulations of Herbal Extracts in Treatment of Neurodegenerative Disorders. Front. Bioeng. Biotechnol. 2020, 8, 238.