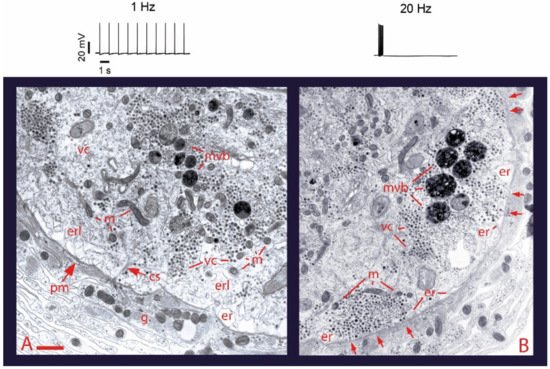
Somatic exocytosis of serotonin can be evoked in Retzius cells in a frequency-dependent manner by trains of action potentials but not by individual action potentials
[32][40][32,40]. Exocytosis on stimulation trains of ten impulses can be conveniently visualized from the formation of fluorescent spots of FM1-43 right inside the plasma membrane
[32]. Each spot forms upon the accumulation of dye inside a cluster of vesicles that undergo exo-endocytosis cycles. Although 1 Hz stimulation fails to evoke exocytosis, a number of fluorescent spots are produced by constitutive exocytosis. Such staining occurs equally well in the absence of stimulation, on 1 Hz stimulation, or on 20 Hz stimulation under experimental conditions that eliminate calcium entry
[32]. Above 5 Hz, the number of fluorescent spots increases with the stimulation frequency
[40].
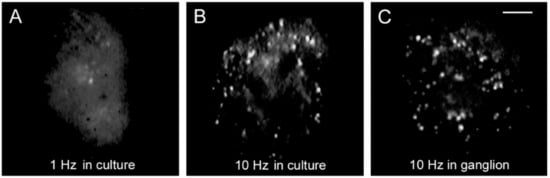
Figure 3 compares the FM1-43 staining patterns of cultured neurons, one stimulated with 1 Hz train (
Figure 3A) and another stimulated with 10 Hz train (
Figure 3B). The staining pattern produced in culture by 10 Hz stimulation reproduces that of neurons stimulated in the ganglion (
Figure 3C), therefore, the culture conditions provide an accurate bioassay to follow the sequence of events that led to serotonin exocytosis
[32]. A plateau in the amount of release at 20 Hz in Retzius neurons, in lobster serotonergic neurons and in mammalian peptidergic neurons
[40][41][42][40,41,42], suggests a generalized peak of efficiency for the coupling between electrical activity, calcium and exocytosis. For the reasons above, the experiments to be described next were carried out by stimulating neurons with 1 Hz or 20 Hz trains in combination with different experimental manipulations.
Figure 3. Somatic exocytosis as measured from FM1-43 fluorescence staining. (
A) The fluorescent FM1-43 spots after stimulation with a 1 Hz train correspond to constitutive exocytosis. (
B) Increase in the number of fluorescent spots after a 10 Hz train. (
C) Similar staining pattern in the soma of a neuron stimulated in the ganglion. Scale bar = 10 μm. Obtained with permission from
[32].
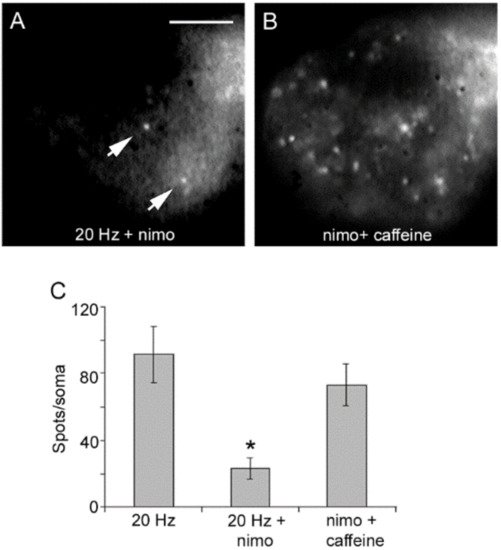
Blockade of L-type channels through the incubation of neurons with nimodipine abolishes exocytosis upon 20 Hz stimulation without affecting the constitutive exocytosis (
Figure 4A)
[32][40][32,40]. Subsequent addition of caffeine to such same neurons to promote release of intracellular calcium through ryanodine receptors
[43][44][45][46][43,44,45,46] in the absence of electrical stimulation produced a bulk of exocytosis, seen as an increased number of fluorescent spots (
Figure 4B). Such results clarified that calcium entry upon activation of L-type channels promotes both vesicle transport and exocytosis. Moreover, they suggested that calcium-induced calcium release contributes to link excitation and exocytosis.
Figure 4. Activation of L-type calcium channels mediate somatic exocytosis. (
A) Incubation with nimodipine (nimo) abolishes evoked but not constitutive exocytosis upon 20 Hz stimulation. The spots pointed by the arrow remained in B. (
B) Caffeine in the presence of nimodipine evokes vesicle transport and exocytosis in the absence of electrical stimulation. Images are selected areas of the soma. (
C) Exocytosis quantified as the number of fluorescent FM1-43 spots per soma. Bars are the S.E.M.; the * indicates a
p < 0.05 T test significance with respect to the 20 Hz and nimo+caffeine groups. The 20 Hz and nimo+caffeine groups were similar (
p > 0.05). Reproduced with permission from the publisher of
[32].