Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Dean Liu and Version 1 by Antonieta Guerrero-Plata.
Viruses are recognized by several Toll-like receptors (TLRs), including TLR8, which is known to bind ssRNA structures. However, the similarities between TLR8 and TLR7 have obscured the distinctive characteristics of TLR8 activation and its importance in the immune system.
- TLRs
- TLR8
- TLR7
- virus
- viral infections
- TLR agonists
1. Activation and Regulation of TLR8
Human TLR8 is expressed in monocytes, macrophages, neutrophils, myeloid dendritic cells [3,6,8,9[1][2][3][4][5],10], and regulatory T (Treg) cells [11][6] (Figure 1a). TLR8 is a type I transmembrane receptor characterized by three structural components: an extracellular domain-containing leucine-rich repeats (LRRs), a transmembrane domain, and a cytoplasmic Toll-interleukin 1 receptor (TIR) domain [12][7]. Its extracellular domain is integrated by approximately 800 amino acids and 26 LRR modules and allows TLR8 to recognize ssRNA. In the absence of a ligand, TLR8 forms a dimer that suffers a conformational change upon activation by recognition of its target [13][8]. Different from other TLRs, TLR8 forms a ring-shaped structure [12][7], which requires proteolytic cleavage at the Z-loop region [14][9]. Between species, the ectodomain (LRRs) determines whether signaling is initiated in response to a ligand stimulation, and within the ectodomain the RQSYA motif has shown to be essential for the TLR8 activation [15][10]. Although the sequence-specific recognition of RNA by TLR8 has not been fully established, it is known that TLR8 senses ssRNA throughout its ability to form secondary structures [16][11] and by recognizing ssRNA AU- and GU-rich sequences [17][12]. That differentiates it from TLR7, which is activated by GU-rich sequences [18][13]. In both cases, TLR7 and TLR8 use the same ligand-binding site, but with different amino acid composition [12][7]. Apparently, TLR8 is able to distinguish the host RNA by nucleoside modifications and only activates the signaling response when a non-modified RNA enters the cell [19][14]. TLR8 is also suggested to be a vita-PAMP receptor that is able to recognize microbial structures from viable microbes [20][15], poly(A)/T sequences [21][16] or even small antiviral molecules [12][7]. Overall, there are several agonists identified to specifically activate TLR8 or TLR7, and those that can activate both receptors (Table 1).
Activation of TLR8 can be triggered by multiple known ligands such as viral ssRNA [4][17], miRNAs [16,22,23][11][18][19] and some agonists included in Table 1. That activation can promote co-stimulation and MHC class II expression to induce proliferation of naïve CD4 T cells [24][20] and Th1 differentiation [25][21]. In human regulatory T cells (Treg), TLR8 agonists can mediate the reversal of the suppressive function of Treg cells through the TLR8-MyD88-IRAK4 signaling pathway [11][6]. Additionally, TLR8 has proven to be an important driver of T follicular helper (Tfh) cell differentiation [20][15]. Interestingly, experimental evidence suggests that TLR8 may also have a regulatory effect on other endosomal TLRs. In fact, TLR8 is able to inhibit both TLR7 and TLR9 in in vitro cells [26][22]. This observation was reproduced in Tlr8−/− mice, where the absence of TLR8 led to higher levels of expression of TLR7 and interferon-stimulated genes (ISGs), an effect that may account for an increased antiviral immunity in the infected mice [27][23].
However, whether the inhibition of TLR7 and TLR9 by TLR8 is through direct or indirect physical contact is still unknown. Furthermore, the regulation of TLR8 by other TLRs has also been suggested. In THP1 cells, a human monocyte cell line, the addition of both TLR7 and TLR8 ligands has shown an apparent inhibition of TLR8-induced cytokine expression, suggesting that TLR7 could have a modulatory effect on TLR8 responsiveness [28][24]. The impact of other TLRs on TLR8 expression and activation is not yet well defined, as some contradictions exist. Studies in HEK cells suggest that TLR9 inhibits TLR7, but neither TLR7 nor TLR9 inhibit TLR8 [26,29][22][25]. However, studies in mice indicate that TLR7 may negatively regulate TLR8, where the absence of TLR7 led to an upregulation of TLR8 expression, suggesting a compensatory mechanism in the immune response [30][26]. Future research will define the contribution of other endosomal TLRs on TLR8 function.
2. TLR8 and Viral Infections
The strategic localization of TLR8 in the endosomes allows for the recognition of several viruses, mainly because it recognizes ssRNAs. Viral ssRNA entering the cell would colocalize into early endosomes around 15 to 20 min after infection, where the RNA could bind to TLR8 [31][27].
Nevertheless, in the case of human immunodeficiency virus (HIV), the protein SNAPIN inhibits the colocalization of TLR8 with HIV[32][28]. Interestingly, studies with vaccinia virus (VV) infection in murine plasmacytoid dendritic cells (pDCs) indicate that poly(A)/T-rich DNA could also be recognized by TLR8, upregulating the expression of IFN-α and IFN-β [21][16]. Another example of TLR8 activation by viruses is provided by Coxsackie B virus (CBV), which induced an inflammatory response mediated mostly through TLR8 and, to a lesser extent, through TLR7 [4][17], similar effect is observed in Influenza A Virus (H3N2) [33][29]. In West Nile virus (WNV)-infected bone-marrow-derived dendritic cells from tlr8−/− mice, the lack of TLR8 resulted in an improved antiviral response due to an increase of TLR7 expression, likely as a result of a compensatory effect [27][23].
TLR8 expression has been documented to increase around 2 to 6 h after some viral infections such as human parechovirus 1 (HPEV-1) [31][27] and influenza virus (H5N1) [49][45]. However, the response against viral infection after TLR8 activation highly depends on the type of cell infected. For instance, in human primary CD4+ T cells, human immunodeficiency virus 1 (HIV-1) is endocytosed by CD4 T cells and activates TLR8. This response triggers a cytokine response that helps the viral replication and reactivates latent HIV-1 through pro-inflammatory stimuli [50][46]. However, in human monocytes, HIV-1 exposure initially induces autophagy through a TLR8- and BECN1-dependent mechanism, an event that is eventually modulated by the productive infection of the virus [51][47].
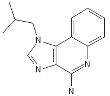
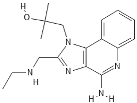
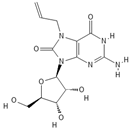
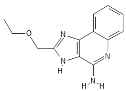
Table 2. Effect of viruses on TLR8 response.
| DNA Virus | |||||||
|---|---|---|---|---|---|---|---|
| Virus | Effect on TLR8 Activation | Cells | |||||
| DNA Viruses | |||||||
| Vaccinia virus | Induces the expression of IFN-α, IFN-β | pDCs, HEK cells [21][16] | |||||
| Hepatitis B virus | HBV infection inhibits innate immunity by decreasing TLR8 levels | PBMC [52][48] | [38,39,40,41][34][35][36][37] | ||||
| Isatoribine | 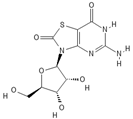 |
Nucleoside analogue | [42][38] | ||||
| RNA Viruses | TLR8 | TL8-506 | 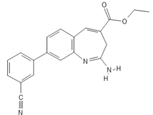 |
Benzoazepine compound | [43][39 | ||
| Influenza A virus (H3N2) | ] | ||||||
| Induces the expression of | IL-8 |
HEK cells, neutrophils [33][29] | Selgantolimod | 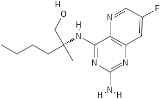 |
Nucleotide analogue | ||
| Coxsackie B virus | Induces the expression of IFN-β, IL-6 | Human cardiac cells [4][17] | [44,45][40][41] | ||||
| TLR7/TLR8 | Resiquimod | 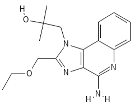 |
Imidazoquinoline compound derivative | Induces the expression of IL-6 and IL-1β |
Human primary CD4+ T cells [50][46][24,38,40][20][34][36] | ||
| HIV-1 | CL075 | 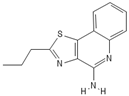 |
Thiazoloquinolone derivative | ||||
| Colocalization of HIV and TLR8 is decreased | Dendritic cells [32][28] | [ | 29,46,47,48 | ||||
| West Nile virus | TLR8 favors the infection in infected mice | Bone-marrow-derived dendritic cells (BMDCs) [27][23] | |||||
| Zika virus | TLR8 and MyD88 expression decreases | Peripheral blood [53][49] | |||||
3. Signaling Pathways Associated with TLR8 and Its Differences with TLR7
Although both TLR7 and TLR8 recognize ssRNA, they activate different signaling pathways to promote the expression of inflammatory cytokines and interferons used in the defense against viruses (Figure 1b). Both TLR7 and TLR8 signaling pathways are mediated by the adaptor molecule MyD88, which is expressed ubiquitously in the cell cytoplasm. After a viral infection, there is a rapid redistribution of MyD88 to the endosomal compartment, where the TIR domains of TLR8 phosphorylate MyD88 [4,12][17][7]. Activation of MyD88 pathway elicits a robust inflammatory and interferon induction by the activation of TRAF6, NF-kB, IRF-7, IRF-3, AP-1, and p38 MAPK [4,25][17][21].
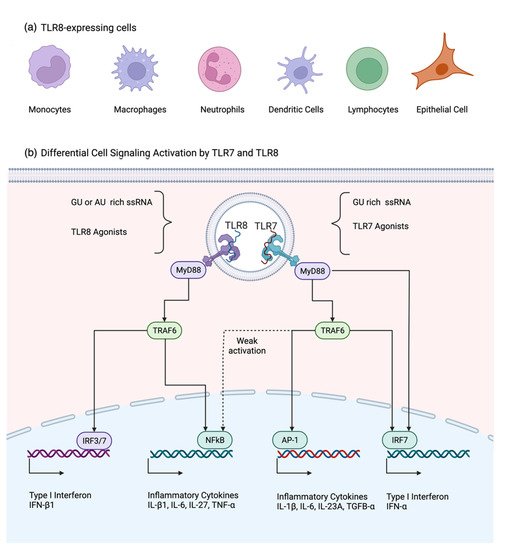
Figure 1. (a) Cells that are reported to express TLR8 under basal conditions; (b) TLR7 and TLR8 signaling pathways have the common adaptor protein MyD88. IFN-α expression after TLR7 can be achieved by forming the complex MyD88-IRF7 or by the activation of IRF7 via TRAF6. TRAF6 also activates AP-1 and weakly NF-kB, promoting inflammatory cytokine expression. TLR8 induces a more potent inflammatory cytokine response than TLR7 via the activation of TRAF6-NF-kB pathway, while the induction of IFN-b by TLR8 is achieved through the activation of the IRF3/7 pathway. Created with BioRender.com. (Accessed on 17 December 2021.)
Although MyD88 has been seen as a common adaptor molecule to TLR7 and TLR8, the activation of this protein in monocytes and macrophages modulates different immune responses with different signature profiles of pro-inflammatory cytokines and type I interferon in a cell-dependent fashion [8,9,25][3][4][21]. In the case of pro-inflammatory cytokines, TLR7 induces more IL-1β, IL-6, IP-10 and IL-23, whereas TLR8 agonists induce more IL-1α/β, IL-6, IL-8, TNF-α, IL-12β, IL-27 and MIP-1α in myeloid dendritic cells [25,48,56][21][44][50]. Moreover, TLR7 agonist stimulation in monocytes fails to induce a robust NF-kB. However, it induces the activation of AP-1, which may explain the differential cytokine profile elicited by TLR8 and TLR7 agonists [25][21]. A similar phenomenon is observed in the production of IFN by the activation of these two receptors. While TLR7 activation induces the expression of IFN-α1 and IFN-α2 in monocytes and pDCs [56,57][50][51], TLR8 activation in monocytes and myeloid DCs expresses more IFN-β1 [8,9,25,56][3][4][21][50]. After TLR7 ligand activation, the adaptor molecule MyD88 can stimulate IRF through two different mechanisms. First, MyD88 can phosphorylate IRF7, forming a complex that promotes the IFN response. Second, MyD88 activates TRAF6, which binds to IRF7, inducing the expression of IFN-α [58][52]. This experimental evidence illustrates the unique characteristics of TLR8 in triggering the inflammatory and antiviral host responses.
4. Future Perspectives for Therapeutics with TLR8 Agonist for Viral Infections
Commercially available TLR8 agonists have demonstrated the capability to selectively activate TLR8. There are many properties of these molecules that make them attractive candidates for use as vaccine adjuvants or antiviral activity [6,59][2][53]. These molecules can be administered through multiple methods including intravenously, oral [44][40], inhaled, or topically applied, which make them good candidates as therapeutical options for the future. Currently, TLR8 agonists are being investigated as vaccine adjuvants and in some other ailments such as allergy or asthma [60][54]. One example is that of HIV Gag protein, which was conjugated to TLR7/8 agonists as an effective way to elicit a broad-based adaptive immunity in nonhuman primates [61][55]. Moreover, recent work has demonstrated that the TLR7/8 agonist imidazoquinoline coupled to a novel amphiphilic carrier enhances vaccine efficiency by inducing a robust Th1 skewed antibody response in mice treated with a single shot with spike protein for SARS-CoV-2 or seasonal quadrivalent inactivated influenza virus vaccine [62][56]. Selgantolimod (GS-9688), also known as oral (R)-7, is a TLR8 agonist that has a good absorption properties, and is currently in phase 2 clinical trial (NCT04891770) for the treatment of chronic hepatitis B [44][40]. Some progress has also been done on the use of TLR8 agonists in HIV infection, where it is shown that T CD4+ TLR8 promoted Tfh differentiation towards Tfh1 and Tfh17 during HIV infection. Such differentiation and the cytokine secretion from TCD4+ via TLR8 could be exploited as a potential therapeutic target and vaccine development [50][46].
TLR8 agonists in combination with other ligands such as fms-like tyrosine kinase (Flt3L) have demonstrated to prime Ag-specific CD8+ T cells, suggesting that TLR8 ligands could also be used as potent adjuvants to prime functionally superior Ag-specific human CD8+ T cells and improve the response to viral infections [63][57].
References
- Kolli, D.; Velayutham, T.S.; Casola, A. Host-Viral Interactions: Role of Pattern Recognition Receptors (PRRs) in Human Pneumovirus Infections. Pathogens 2013, 2, 232–263.
- Tomai, M.A.; Vasilakos, J.P. Chapter 8 - Toll-Like Receptor 7 and 8 Agonists for Vaccine Adjuvant Use. In Immunopotentiators in Modern Vaccines, 2nd ed.; Schijns, V.E.J.C., O’Hagan, D.T., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 8; pp. 149–162. ISBN 978-0-12-804019-5.
- Bender, A.T.; Tzvetkov, E.; Pereira, A.; Wu, Y.; Kasar, S.; Przetak, M.M.; Vlach, J.; Niewold, T.B.; Jensen, M.A.; Okitsu, S.L. TLR7 and TLR8 Differentially Activate the IRF and NF-ΚB Pathways in Specific Cell Types to Promote Inflammation. ImmunoHorizons 2020, 4, 93–107.
- Eng, H.-L.; Hsu, Y.-Y.; Lin, T.-M. Differences in TLR7/8 Activation between Monocytes and Macrophages. Biochem. Biophys. Res. Commun. 2018, 497, 319–325.
- Maris, N.A.; Dessing, M.C.; de Vos, A.F.; Bresser, P.; van der Zee, J.S.; Jansen, H.M.; Spek, C.A.; Poll, T. van der Toll-like Receptor MRNA Levels in Alveolar Macrophages after Inhalation of Endotoxin. Eur. Respir. J. 2006, 28, 622–626.
- Peng, G.; Guo, Z.; Kiniwa, Y.; Voo, K.S.; Peng, W.; Fu, T.; Wang, D.Y.; Li, Y.; Wang, H.Y.; Wang, R.-F. Toll-Like Receptor 8-Mediated Reversal of CD4+ Regulatory T Cell Function. Science 2005, 309, 1380–1384.
- Ohto, U.; Tanji, H.; Shimizu, T. Structure and Function of Toll-like Receptor 8. Microbes Infect. 2014, 16, 273–282.
- Zhu, J.; Brownlie, R.; Liu, Q.; Babiuk, L.A.; Potter, A.; Mutwiri, G.K. Characterization of Bovine Toll-like Receptor 8: Ligand Specificity, Signaling Essential Sites and Dimerization. Mol. Immunol. 2009, 46, 978–990.
- Tanji, H.; Ohto, U.; Motoi, Y.; Shibata, T.; Miyake, K.; Shimizu, T. Autoinhibition and Relief Mechanism by the Proteolytic Processing of Toll-like Receptor 8. Proc. Natl. Acad. Sci. USA 2016, 113, 3012–3017.
- Liu, J.; Xu, C.; Hsu, L.-C.; Luo, Y.; Xiang, R.; Chuang, T.-H. A Five-Amino-Acid Motif in the Undefined Region of the TLR8 Ectodomain Is Required for Species-Specific Ligand Recognition. Mol. Immunol. 2010, 47, 1083.
- Pluta, L.; Yousefi, B.; Damania, B.; Khan, A.A. Endosomal TLR-8 Senses MicroRNA-1294 Resulting in the Production of NFḱB Dependent Cytokines. Front. Immunol. 2019, 10, 2860.
- Zhang, Y.; El-Far, M.; Dupuy, F.P.; Abdel-Hakeem, M.S.; He, Z.; Procopio, F.A.; Shi, Y.; Haddad, E.K.; Ancuta, P.; Sekaly, R.-P.; et al. HCV RNA Activates APCs via TLR7/TLR8 While Virus Selectively Stimulates Macrophages Without Inducing Antiviral Responses. Sci. Rep. 2016, 6, 29447.
- Forsbach, A.; Nemorin, J.-G.; Montino, C.; Müller, C.; Samulowitz, U.; Vicari, A.P.; Jurk, M.; Mutwiri, G.K.; Krieg, A.M.; Lipford, G.B.; et al. Identification of RNA Sequence Motifs Stimulating Sequence-Specific TLR8-Dependent Immune Responses. J Immunol. 2008, 180, 3729–3738.
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175.
- Ugolini, M.; Gerhard, J.; Burkert, S.; Jensen, K.J.; Georg, P.; Ebner, F.; Volkers, S.M.; Thada, S.; Dietert, K.; Bauer, L.; et al. Recognition of Microbial Viability via TLR8 Drives TFH Cell Differentiation and Vaccine Responses. Nat. Immunol. 2018, 19, 386–396.
- Martinez, J.; Huang, X.; Yang, Y. Toll-like Receptor 8-Mediated Activation of Murine Plasmacytoid Dendritic Cells by Vaccinia Viral DNA. Proc. Natl. Acad. Sci. USA 2010, 107, 6442–6447.
- Triantafilou, K.; Orthopoulos, G.; Vakakis, E.; Ahmed, M.A.E.; Golenbock, D.T.; Lepper, P.M.; Triantafilou, M. Human Cardiac Inflammatory Responses Triggered by Coxsackie B Viruses Are Mainly Toll-like Receptor (TLR) 8-Dependent. Cell. Microbiol. 2005, 7, 1117–1126.
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs Bind to Toll-like Receptors to Induce Prometastatic Inflammatory Response. Proc Natl Acad Sci USA 2012, 109, E2110–E2116.
- Bernard, M.A.; Han, X.; Inderbitzin, S.; Agbim, I.; Zhao, H.; Koziel, H.; Tachado, S.D. HIV-Derived SsRNA Binds to TLR8 to Induce Inflammation-Driven Macrophage Foam Cell Formation. PLoS ONE 2014, 9, e104039.
- Hackstein, H.; Knoche, A.; Nockher, A.; Poeling, J.; Kubin, T.; Jurk, M.; Vollmer, J.; Bein, G. The TLR7/8 Ligand Resiquimod Targets Monocyte-Derived Dendritic Cell Differentiation via TLR8 and Augments Functional Dendritic Cell Generation. Cell. Immunol. 2011, 271, 401–412.
- de Marcken, M.; Dhaliwal, K.; Danielsen, A.C.; Gautron, A.S.; Dominguez-Villar, M. TLR7 and TLR8 Activate Distinct Pathways in Monocytes during RNA Virus Infection. Sci. Signal. 2019, 12.
- Wang, J.; Shao, Y.; Bennett, T.A.; Shankar, R.A.; Wightman, P.D.; Reddy, L.G. The Functional Effects of Physical Interactions among Toll-like Receptors 7, 8, and 9*. J. Biol. Chem. 2006, 281, 37427–37434.
- Paul, A.M.; Acharya, D.; Le, L.; Wang, P.; Stokic, D.S.; Leis, A.A.; Alexopoulou, L.; Town, T.; Flavell, R.A.; Fikrig, E.; et al. TLR8 Couples SOCS-1 and Restrains TLR7-Mediated Antiviral Immunity, Exacerbating West Nile Virus Infection in Mice. J. Immunol. 2016, 197, 4425–4435.
- Odoardi, N.; Kourko, O.; Petes, C.; Basta, S.; Gee, K. TLR7 Ligation Inhibits TLR8 Responsiveness in IL-27-Primed Human THP-1 Monocytes and Macrophages. J. Innate Immun. 2021, 1–14.
- Wang, C.-H.; Eng, H.-L.; Lin, K.-H.; Chang, C.-H.; Hsieh, C.-A.; Lin, Y.-L.; Lin, T.-M. TLR7 and TLR8 Gene Variations and Susceptibility to Hepatitis C Virus Infection. PLOS ONE 2011, 6, e26235.
- Awais, M.; Wang, K.; Lin, X.; Qian, W.; Zhang, N.; Wang, C.; Wang, K.; Zhao, L.; Fu, Z.F.; Cui, M. TLR7 Deficiency Leads to TLR8 Compensative Regulation of Immune Response against JEV in Mice. Front. Immunol. 2017, 8, 160.
- Triantafilou, K.; Vakakis, E.; Orthopoulos, G.; Ahmed, M.A.E.; Schumann, C.; Lepper, P.M.; Triantafilou, M. TLR8 and TLR7 Are Involved in the Host’s Immune Response to Human Parechovirus 1. Eur. J. Immunol. 2005, 35, 2416–2423.
- Khatamzas, E.; Hipp, M.M.; Gaughan, D.; Pichulik, T.; Leslie, A.; Fernandes, R.A.; Muraro, D.; Booth, S.; Zausmer, K.; Sun, M.; et al. Snapin Promotes HIV-1 Transmission from Dendritic Cells by Dampening TLR8 Signaling. EMBO J. 2017, 36, 2998–3011.
- Wang, J.P.; Bowen, G.N.; Padden, C.; Cerny, A.; Finberg, R.W.; Newburger, P.E.; Kurt-Jones, E.A. Toll-like Receptor–Mediated Activation of Neutrophils by Influenza A Virus. Blood 2008, 112, 2028–2034.
- Larangé, A.; Antonios, D.; Pallardy, M.; Kerdine-Römer, S. TLR7 and TLR8 Agonists Trigger Different Signaling Pathways for Human Dendritic Cell Maturation. J. of Leukoc. Biol. 2009, 85, 673–683.
- Craft, N.; Bruhn, K.W.; Nguyen, B.D.; Prins, R.; Lin, J.W.; Liau, L.M.; Miller, J.F. The TLR7 Agonist Imiquimod Enhances the Anti-Melanoma Effects of a Recombinant Listeria Monocytogenes Vaccine. J. Immunol. 2005, 175, 1983–1990.
- Buitendijk, M.; Eszterhas, S.K.; Howell, A.L. Gardiquimod: A Toll-Like Receptor-7 Agonist That Inhibits HIV Type 1 Infection of Human Macrophages and Activated T Cells. AIDS Res. Hum. Retrovir. 2013, 29, 907–918.
- Enosi Tuipulotu, D.; Netzler, N.E.; Lun, J.H.; Mackenzie, J.M.; White, P.A. TLR7 Agonists Display Potent Antiviral Effects against Norovirus Infection via Innate Stimulation. Antimicrob. Agents and Chemother. 2018, 62, e02417-17.
- Jurk, M.; Kritzler, A.; Schulte, B.; Tluk, S.; Schetter, C.; Krieg, A.M.; Vollmer, J. Modulating Responsiveness of Human TLR7 and 8 to Small Molecule Ligands with T-Rich Phosphorothiate Oligodeoxynucleotides. Eur. J. Immunol. 2006, 36, 1815–1826.
- Dzopalic, T.; Dragicevic, A.; Vasilijic, S.; Vucevic, D.; Majstorovic, I.; Bozic, B.; Balint, B.; Colic, M. Loxoribine, a Selective Toll-like Receptor 7 Agonist, Induces Maturation of Human Monocyte-Derived Dendritic Cells and Stimulates Their Th-1- and Th-17-Polarizing Capability. Int. Immunopharmacol. 2010, 10, 1428–1433.
- Heil, F.; Ahmad-Nejad, P.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Gellert, T.; Dietrich, H.; Lipford, G.; Takeda, K.; Akira, S.; et al. The Toll-like Receptor 7 (TLR7)-Specific Stimulus Loxoribine Uncovers a Strong Relationship within the TLR7, 8 and 9 Subfamily. Eur. J. Immunol. 2003, 33, 2987–2997.
- Xu, N.; Yao, H.-P.; Lv, G.-C.; Chen, Z. Downregulation of TLR7/9 Leads to Deficient Production of IFN-α from Plasmacytoid Dendritic Cells in Chronic Hepatitis B. Inflamm. Res. 2012, 61, 997–1004.
- Horsmans, Y.; Berg, T.; Desager, J.-P.; Mueller, T.; Schott, E.; Fletcher, S.P.; Steffy, K.R.; Bauman, L.A.; Kerr, B.M.; Averett, D.R. Isatoribine, an Agonist of TLR7, Reduces Plasma Virus Concentration in Chronic Hepatitis C Infection. Hepatology 2005, 42, 724–731.
- Serrano, R.; Wesch, D.; Kabelitz, D. Activation of Human Γδ T Cells: Modulation by Toll-Like Receptor 8 Ligands and Role of Monocytes. Cells 2020, 9, 713.
- Mackman, R.L.; Mish, M.; Chin, G.; Perry, J.K.; Appleby, T.; Aktoudianakis, V.; Metobo, S.; Pyun, P.; Niu, C.; Daffis, S.; et al. Discovery of GS-9688 (Selgantolimod) as a Potent and Selective Oral Toll-Like Receptor 8 Agonist for the Treatment of Chronic Hepatitis B. J. Med. Chem. 2020, 63, 10188–10203.
- Amin, O.E.; Colbeck, E.J.; Daffis, S.; Khan, S.; Ramakrishnan, D.; Pattabiraman, D.; Chu, R.; Micolochick Steuer, H.; Lehar, S.; Peiser, L.; et al. Therapeutic Potential of TLR8 Agonist GS-9688 (Selgantolimod) in Chronic Hepatitis B: Remodeling of Antiviral and Regulatory Mediators. Hepatology 2021, 74, 55–71.
- Du, J.; Wu, Z.; Ren, S.; Wei, Y.; Gao, M.; Randolph, G.J.; Qu, C. TLR8 Agonists Stimulate Newly Recruited Monocyte-Derived Cells into Potent APCs That Enhance HBsAg Immunogenicity. Vaccine 2010, 28, 6273–6281.
- Chatillon, J.-F.; Hamieh, M.; Bayeux, F.; Abasq, C.; Fauquembergue, E.; Drouet, A.; Guisier, F.; Latouche, J.-B.; Musette, P. Direct Toll-Like Receptor 8 Signaling Increases the Functional Avidity of Human CD8+ T Lymphocytes Generated for Adoptive T Cell Therapy Strategies. Immun. Inflamm. Dis. 2015, 3, 1–13.
- Wu, J.; Li, S.; Li, T.; Lv, X.; Zhang, M.; Zang, G.; Qi, C.; Liu, Y.-J.; Xu, L.; Chen, J. PDC Activation by TLR7/8 Ligand CL097 Compared to TLR7 Ligand IMQ or TLR9 Ligand CpG. J. Immunol. Res. 2019, 2019, 1749803.
- Song, B.M.; Kang, Y.M.; Kim, H.S.; Seo, S.H. Induction of Inflammatory Cytokines and Toll-like Receptors in Human Normal Respiratory Epithelial Cells Infected with Seasonal H1N1, 2009 Pandemic H1N1, Seasonal H3N2, and Highly Pathogenic H5N1 Influenza Virus. Viral Immunol. 2011, 24, 179–187.
- Meås, H.Z.; Haug, M.; Beckwith, M.S.; Louet, C.; Ryan, L.; Hu, Z.; Landskron, J.; Nordbø, S.A.; Taskén, K.; Yin, H.; et al. Sensing of HIV-1 by TLR8 Activates Human T Cells and Reverses Latency. Nat. Commun. 2020, 11, 147.
- Campbell, G.R.; Rawat, P.; Bruckman, R.S.; Spector, S.A. Human Immunodeficiency Virus Type 1 Nef Inhibits Autophagy through Transcription Factor EB Sequestration. PLoS Pathog. 2015, 11, e1005018.
- Deng, G.; Ge, J.; Liu, C.; Pang, J.; Huang, Z.; Peng, J.; Sun, J.; Hou, J.; Zhang, X. Impaired Expression and Function of TLR8 in Chronic HBV Infection and Its Association with Treatment Responses during Peg-IFN-α-2a Antiviral Therapy. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 386–398.
- da Silva, M.H.M.; Moises, R.N.C.; Alves, B.E.B.; Pereira, H.W.B.; de Paiva, A.A.P.; Morais, I.C.; Nascimento, Y.M.; Monteiro, J.D.; de Souto, J.T.; Nascimento, M.S.L.; et al. Innate Immune Response in Patients with Acute Zika Virus Infection. Med. Microbiol. Immunol. 2019, 208, 703–714.
- Gorden, K.B.; Gorski, K.S.; Gibson, S.J.; Kedl, R.M.; Kieper, W.C.; Qiu, X.; Tomai, M.A.; Alkan, S.S.; Vasilakos, J.P. Synthetic TLR Agonists Reveal Functional Differences between Human TLR7 and TLR8. J. Immunol. 2005, 174, 1259–1268.
- Hornung, V.; Guenthner-Biller, M.; Bourquin, C.; Ablasser, A.; Schlee, M.; Uematsu, S.; Noronha, A.; Manoharan, M.; Akira, S.; de Fougerolles, A.; et al. Sequence-Specific Potent Induction of IFN-α by Short Interfering RNA in Plasmacytoid Dendritic Cells through TLR7. Nat. Med. 2005, 11, 263–270.
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-α Induction through Toll-like Receptors Involves a Direct Interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004, 5, 1061–1068.
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Sousa, C.R. e Innate Antiviral Responses by Means of TLR7-Mediated Recognition of Single-Stranded RNA. Science 2004, 303, 1529–1531.
- Aryan, Z.; Holgate, S.T.; Radzioch, D.; Rezaei, N. A New Era of Targeting the Ancient Gatekeepers of the Immune System: Toll-Like Agonists in the Treatment of Allergic Rhinitis and Asthma. Int. Arch. Allergy Immunol. 2014, 164, 46–63.
- Wille-Reece, U.; Flynn, B.J.; Loré, K.; Koup, R.A.; Kedl, R.M.; Mattapallil, J.J.; Weiss, W.R.; Roederer, M.; Seder, R.A. HIV Gag Protein Conjugated to a Toll-like Receptor 7/8 Agonist Improves the Magnitude and Quality of Th1 and CD8+ T Cell Responses in Nonhuman Primates. Proc. Natl. Acad. Sci. USA 2005, 102, 15190–15194.
- Jangra, S.; De Vrieze, J.; Choi, A.; Rathnasinghe, R.; Laghlali, G.; Uvyn, A.; Van Herck, S.; Nuhn, L.; Deswarte, K.; Zhong, Z.; et al. Sterilizing Immunity against SARS-CoV-2 Infection in Mice by a Single-Shot and Lipid Amphiphile Imidazoquinoline TLR7/8 Agonist-Adjuvanted Recombinant Spike Protein Vaccine**. Angew. Chem. Int. Ed. 2021, 60, 9467–9473.
- Lissina, A.; Briceño, O.; Afonso, G.; Larsen, M.; Gostick, E.; Price, D.A.; Mallone, R.; Appay, V. Priming of Qualitatively Superior Human Effector CD8+ T Cells Using TLR8 Ligand Combined with FLT3 Ligand. J. Immunol. 2016, 196, 256–263.
More