Research has indicated that zinc plays a consequential mechanistic role in the protection against oxidative stress as zinc is required for the proper functioning of the antioxidant system, the suppression of inflammatory mediators, and the modulation of zinc transporters. Recently, the mechanisms surrounding ZnT8, ZIP7, and metallothionein have shown to be of particular pathogenic importance and are considered as potential therapeutic targets in disease management. The literature has shown that zinc dysregulation is associated with diabetes and may be considered as a leading contributor to the deleterious vascular alterations exhibited by the disease. Although further investigation is required, studies have indicated the favorable use of zinc supplementation in the protection against and prevention of oxidative stress and its consequences over the course of the condition.
- oxidative stress
- ROS
- zinc homeostasis
- zinc transporters
1. Zinc Homeostasis
As Zn2+ plays a diverse role in cellular processes including cell signaling, enzymatic activity, and gene transcription, homeostatic mechanisms are required to tightly control Zn2+ absorption, distribution, intracellular availability, and excretion [1][2]. The cation Zn2+ cannot cross lipid bilayers and consequently physiological levels are maintained by three groups of proteins which regulate the inflow, outflow, and compartmentalization of Zn2+: the ZnT and ZIP families of Zn2+ transporters and the Zn2+-sensitive metallothioneins (MTs) [3][1][2][4]. The ZnT family (SLC30A) is a group of 10 (ZnT1-ZnT10) cation-diffusion facilitators that transport Zn2+ ions towards the extracellular space or from the cytosol into organelles [5][6]. The ZIP family (SLC39A), ZIP1-ZIP14, passes Zn2+ into the cytoplasm from the extracellular space or from intracellular organelles [5][7][6]. Zn2+ binds with MTs until homeostatic conditions change such that Zn2+ is required to be released and redistributed in the cells (e.g., in a state of oxidative stress, Zn2+ is released from its complex with MT for antioxidant purposes) [8][1]. Movement of the cation is facilitated within a bimodal framework of Zn2+ signaling. Early zinc signaling (EZS) is independent of gene transcription and results in a rapid fluctuation of intracellular Zn2+ levels via efflux from the organelles into the cytosol [7]. Late zinc signaling (LZS) is slower than the response of EZS because it consists of transcriptional changes in genes and includes the use of storage proteins or transporters. Together, both systems regulate processes involved in metabolism, cell differentiation, proliferation, and growth [7][9][10][6][1]. In the liver or muscle, evidence of Zn2+ on cellular signaling is exemplified through the inhibition of protein tyrosine phosphatase 1B (PTP1B). This protein negatively regulates insulin-signaling pathways whereas Zn2+ can extend the insulin signal through the insulin receptor via the inhibition of PTP1B [5][7]. The human MT family consists of 12 operational MTs, with MT1 and MT2 as the major isoforms in the pancreas; these molecules bind Zn2+ with high affinity, incorporating up to seven Zn2+ ions per molecule [11][3][12][13]. As MTs are equipped with reversible dissociation, they can act as Zn2+ donors or acceptors [11][3][12][13]. Downstream cell signaling is induced by releasing Zn2+ either via the oxidation of the sulfur donors in the MT molecule or through the interaction of the MT with nitric oxide (NO) [12].Zinc Distribution in the β-Cell
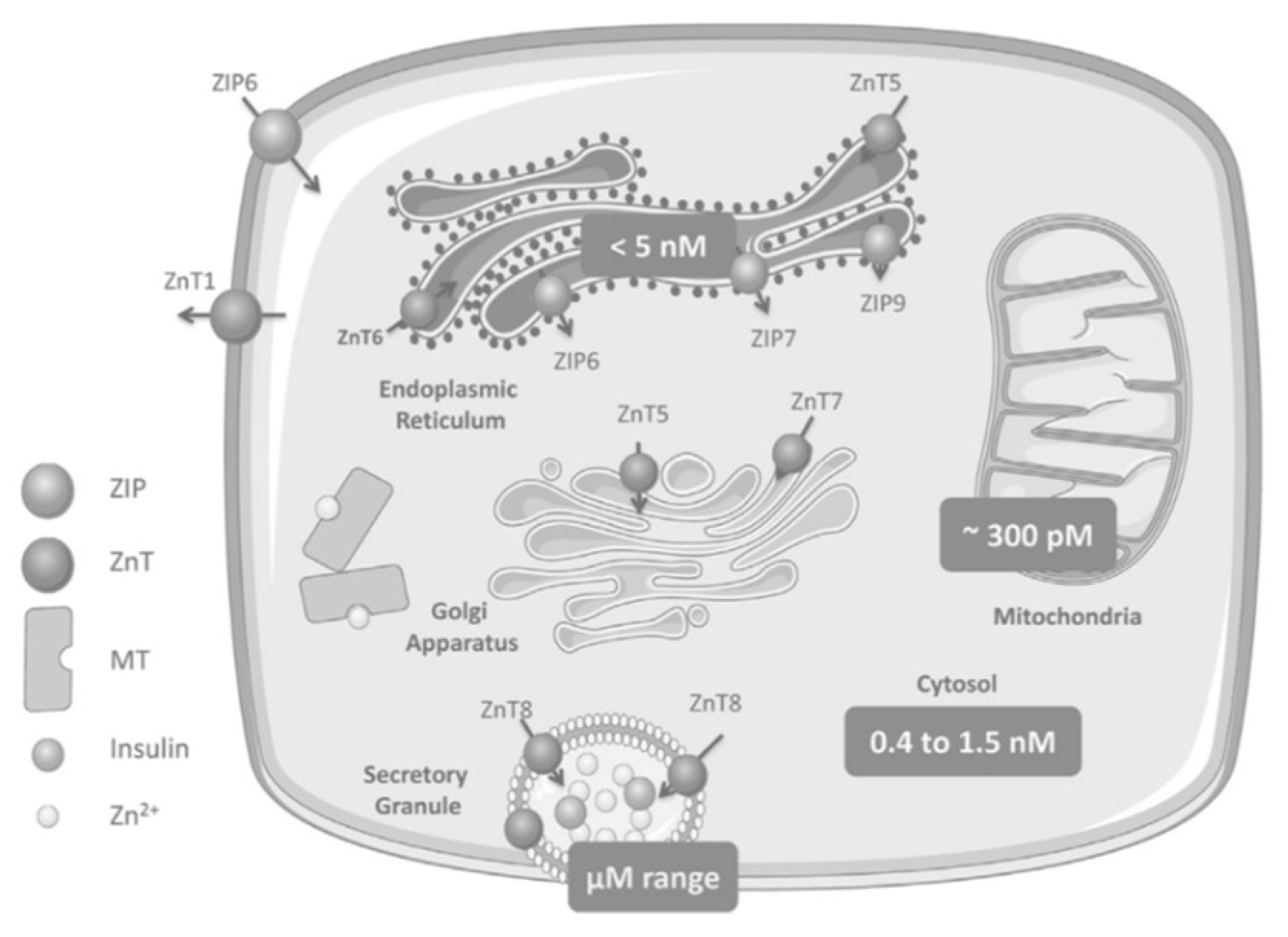
2. Zinc Transporters: Glucose Homeostasis, Insulin Resistance and Immunity
2.1. ZnT8
2.2. ZIP7
While T2DM indicates major metabolic dysfunction and aberrant blood glucose levels, IR is the precursor to this disease and is marked by the body’s inability to respond properly or in a timely fashion [5]. IR is induced by the distortion of accurate cell signaling and effective glucose uptake into peripheral tissues [7]. ZIP7 is responsible for transporting Zn2+ out of the ER and Golgi into the cytosol; it also controls cell signaling pathways (of note, the insulin receptor substrate-phosphoinositide-3-kinase-protein kinase B (IRS-P13K-AKT) pathway) akin to insulin, which initiates glucose uptake in skeletal muscle [5][7]. Literature suggests that ZIP7, as linked with ER stress and cell-signaling pathways, is at the root of IR-associated distortions. As seen in Figure 2, ZIP7 action increases the cytosolic Zn2+ concentration, which participates in a signaling pathway that elicits glucose mobilization and metabolism [5][16]. In this regard, Zn2+ acts as an insulin mimetic through the phosphorylation of AKT and the resultant mobilization of Glut4 transporters to facilitate the influx of glucose in skeletal muscle [7]. In a Zn2+-depleted cytosolic environment, ER protein folding becomes compromised, thereby activating the unfolded protein response (UPR); the rate of folding is thus reduced, but if ER stress does not resolve, apoptosis is triggered [5].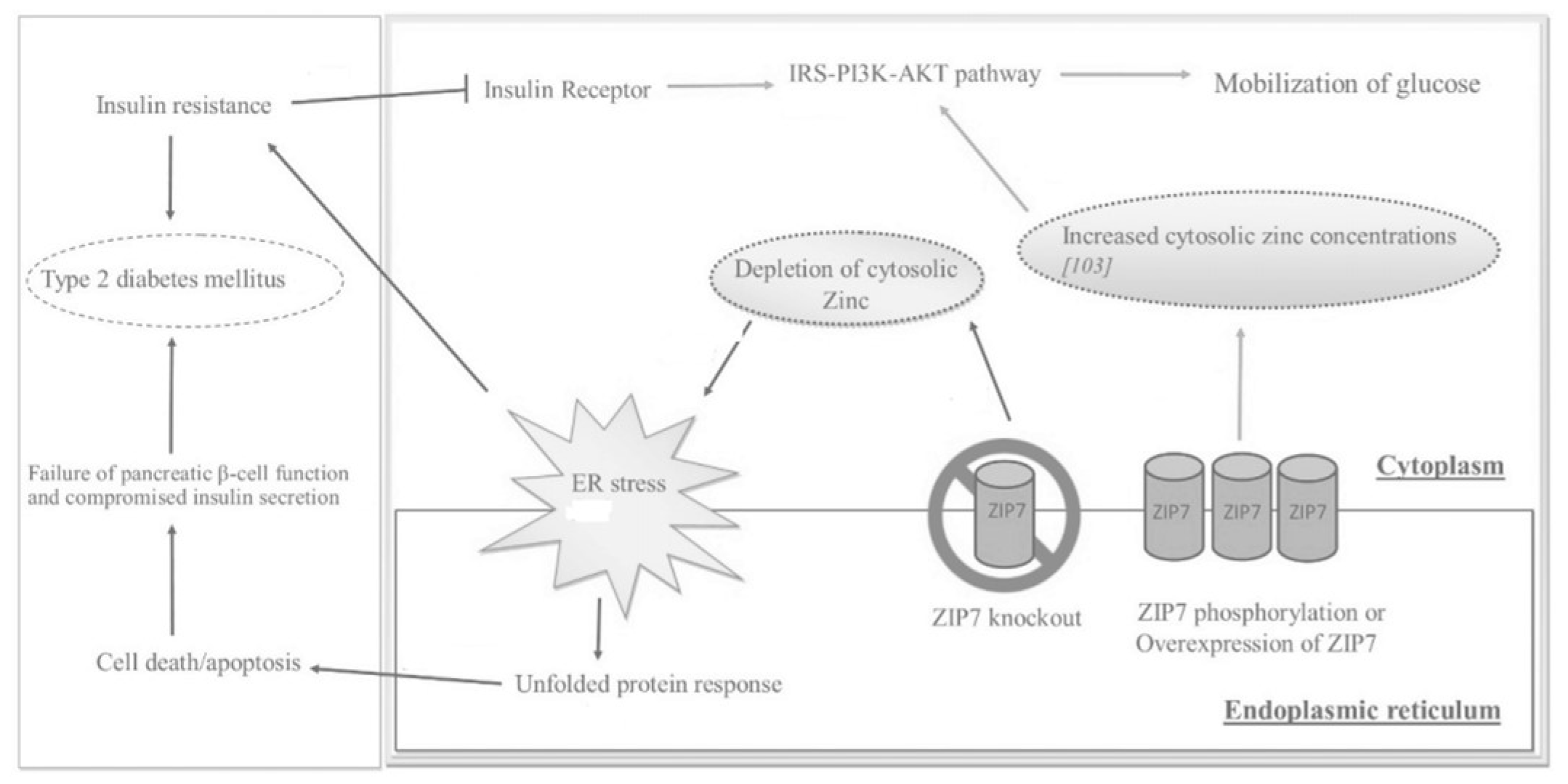 Figure 2. The effects of ZIP7 on glucose mobilization, ER stress, insulin resistance (IR), and T2DM as depicted through various regulatory pathways. Adapted from [5].
Figure 2. The effects of ZIP7 on glucose mobilization, ER stress, insulin resistance (IR), and T2DM as depicted through various regulatory pathways. Adapted from [5].
2.3. Metallotionein
MT is a potent ROS scavenger that offers significant protection against DM and DM-induced cardiovascular injury [17]. Studies of Zn2+ supplementation in diabetic mice have shown that the expression of MT is significantly induced by cellular Zn2+ levels [18][17]. This mechanism has been further confirmed as Zn2+ supplementation up-regulates the expression of MT and consequently decreases diabetes-induced vascular complications [18]. Research has shown that MT single nucleotide polymorphisms are related to various pathological processes, three of which are linked with a significant increase in T2DM prevalence due to reduced MT antioxidant capabilities [11]. Zn2+ effectively determines MT levels through the stimulation of responsive metal transcription factor 1 (MTF-1); this transcription factor directly regulates the expression of MT [19].2.4. Other ZnT/SLC30A Transporters
2.5. Other ZIP/SLC39A Transporters
| Zinc Transporter | Regulators | Effect |
|---|---|---|
| ZnT1 | Metal-responsive mode of regulation; dietary intake of zinc | Efflux of zinc in smooth muscle cells |
| ZnT2 | Metal-responsive mode of regulation; dietary intake of zinc | Zinc transport in vesicles and lysosomes of pancreas, kidney, testis, epithelial cells, small intestine |
| ZnT3 | Glucose status | Transport of zinc to synaptic vesicles |
| ZnT4 | Unaffected by changes in dietary zinc uptake; regulated by extracellular zinc concentrations | Transport of zinc in the trans-Golgi network and in the cytoplasmic vesicular compartment |
| ZnT5 | Glucose status; zinc-responsive elements | Transport of zinc into Golgi lumen for storage |
| ZnT7 | Glucose status | Transport of zinc to Golgi apparatus in retina, liver, epithelial cells, small intestine; may play a redundant role of ZnT8 |
| ZnT8 | Glucose status | Regulation of zinc in the secretory vesicles of pancreatic β-cells |
| ZnT9 | Expressed in low levels in response to dietary intake of zinc | Export of zinc out of myocytes; efflux of zinc in smooth muscle cells |
| ZIP1 | Testosterone and prolactin | Uptake of zinc into cells |
| ZIP6 | Estrogen stimulation, glucose status | Down-regulation leads to poor insulin secretion |
| ZIP7 | Glucose status | Increases cytosolic zinc concentrations that participate in glucose mobilization and metabolism |
| ZIP8 | Glucose status, TNF-∝ in lung epithelial cells | Increases intracellular zinc levels |
| ZIP13 | Gene mutation leads to loss of function | Inhibition of adipocyte browning |
| ZIP14 | Acute phase response during inflammation; IL-6 | Rapid intake of plasma zinc into the organs |
3. Conclusions
References
- Myers, S.A.; Nield, A.; Myers, M. Zinc transporters, mechanisms of action and therapeutic utility: Implications for type 2 diabetes mellitus. J. Nutr. Metab. 2012, 2012, 173712.
- Woodruff, G.; Bouwkamp, C.G.; de Vrij, F.M.; Lovenberg, T.; Bonaventure, P.; Kushner, S.A.; Harrington, A.W. The Zinc Transporter SLC39A7 (ZIP7) Is Essential for Regulation of Cytosolic Zinc Levels. Mol. Pharmacol. 2018, 94, 1092–1100.
- Chabosseau, P.; Rutter, G.A. Zinc and diabetes. Arch Biochem Biophys. 2016, 611, 79–85.
- Zalewski, P.D.; Beltrame, J.F.; Wawer, A.A.; Abdo, A.I.; Murgia, C. Roles for endothelial zinc homeostasis in vascular physiology and coronary artery disease. Crit. Rev. Food Sci. Nutr. 2019, 59, 3511–3525.
- Adulcikas, J.; Sonda, S.; Norouzi, S.; Sohal, S.S.; Myers, S. Targeting the Zinc Transporter ZIP7 in the Treatment of Insulin Resistance and Type 2 Diabetes. Nutrients 2019, 11, 408.
- Fukunaka, A.; Fujitani, Y. Role of Zinc Homeostasis in the Pathogenesis of Diabetes and Obesity. Int. J. Mol. Sci. 2018, 19, 476.
- Norouzi, S.; Adulcikas, J.; Sohal, S.S.; Myers, S. Zinc transporters and insulin resistance: Therapeutic implications for type 2 diabetes and metabolic disease. J. Biomed. Sci. 2017, 24, 87.
- Do Marreiro, D.N.; Cruz, K.J.C.; Morais, J.B.S.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R.S. Zinc and Oxidative Stress: Current Mechanisms. Antioxidants 2017, 6, 24.
- Olechnowicz, J.; Tinkov, A.; Skalny, A.; Suliburska, J. Zinc status is associated with inflammation, oxidative stress, lipid, and glucose metabolism. J. Physiol. Sci. 2018, 68, 19–31.
- Farooq, M. Zinc Deficiency is Associated with Poor Glycemic Control. J. Coll. Physicians Surg. Pak. 2019, 29, 253–257.
- Zhao, T.; Huang, Q.; Su, Y.; Sun, W.; Huang, Q.; Wei, W. Zinc and its regulators in pancreas. Inflammopharmacology 2019, 27, 453–464.
- Choi, S.; Liu, X.; Pan, Z. Zinc deficiency and cellular oxidative stress: Prognostic implications in cardiovascular diseases. Acta Pharmacol. Sin. 2018, 39, 1120–1132.
- Mondragon, P.; Bergdahl, A. Metallothionein expression in slow- vs. fast-twitch muscle fibers following 4 weeks of streptozotocin-induced type 1 diabetes. Facets 2018, 3, 315–325.
- Williams, C.L.; Long, A.E. What has zinc transporter 8 autoimmunity taught us about type 1 diabetes? Diabetologia 2019, 62, 1969–1976.
- Huang, Q.; Du, J.; Merriman, C.; Gong, Z. Genetic, Functional, and Immunological Study of ZnT8 in Diabetes. Int. J. Endocrinol. 2019, 2019, 1524905.
- Anzilotti, C.; Swan, D.J.; Boisson, B.; Deobagkar-Lele, M.; Oliveira, C.; Chabosseau, P.; Engelhardt, K.R.; Xu, X.; Chen, R.; Alvarez, L.; et al. An essential role for the Zn2+ transporter ZIP7 in B cell development. Nat. Immunol. 2019, 20, 350–361.
- Miao, X.; Wang, Y.; Sun, J.; Sun, W.; Tan, Y.; Cai, L.; Zheng, Y.; Su, G.; Liu, Q.; Wang, Y. Zinc protects against diabetes-induced pathogenic changes in the aorta: Roles of metallothionein and nuclear factor (erythroid-derived 2)-like 2. Cardiovasc. Diabetol. 2013, 12, 54.
- Wang, J.; Song, Y.; Elsherif, L.; Song, Z.; Zhou, G.; Prabhu, S.D.; Saari, J.T.; Cai, L. Cardiac metallothionein induction plays the major role in the prevention of diabetic cardiomyopathy by zinc supplementation. Circulation 2006, 113, 544–554.
- Fiorentino, T.V.; Prioletta, A.; Zuo, P.; Folli, F. Hyperglycemia-induced oxidative stress and its role in diabetes mellitus related cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5695–5703.
- Liu, Y.; Batchuluun, B.; Ho, L.; Zhu, D.; Prentice, K.J.; Bhattacharjee, A.; Zhang, M.; Pourasgari, F.; Hardy, A.B.; Taylor, K.M.; et al. Characterization of Zinc Influx Transporters (ZIPs) in Pancreatic β Cells: Roles in regulating cytosolic zinc homeostasis and insulin secretion. J. Biol. Chem. 2015, 290, 18757–18769.
- Chabosseau, P.; Woodier, J.; Cheung, R.; Rutter, G.A. Sensors for measuring subcellular zinc pools. Metallomics 2018, 10, 229–239.
- Aydemir, T.B.; Cousins, R.J. The Multiple Faces of the Metal Transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184.
- Jarosz, M.; Olbert, M.; Wyszogrodzka, G.; Młyniec, K.; Librowski, T. Antioxidant and anti-inflammatory effects of zinc. Zinc-dependent NF-κB signaling. Inflammopharmacology 2017, 25, 11–24.
- Barman, S.; Pradeep, S.R.; Srinivasan, K. Zinc supplementation mitigates its dyshomeostasis in experimental diabetic rats by regulating the expression of zinc transporters and metallothionein. Metallomics 2017, 9, 1765–1777.