Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Yvaine Wei and Version 3 by Sunmin Park.
Skeletal muscle mass plays a critical role in a healthy lifespan by helping to regulate glucose homeostasis. As seen in sarcopenia, decreased skeletal muscle mass impairs glucose homeostasis, but it may also be caused by glucose dysregulation. Gut microbiota modulates lipopolysaccharide (LPS) production, short-chain fatty acids (SCFA), and various metabolites that affect the host metabolism, including skeletal muscle tissues, and may have a role in the sarcopenia etiology. The evidence presented in this entry suggests that loss of muscle mass and function are not an inevitable consequence of the aging process, and that dietary and lifestyle interventions may prevent or delay sarcopenia.
- skeletal muscle mass
- L-leucine
- glucose metabolism
- gut microbiome
- short-chain fatty acids
1. Introduction
Sarcopenia is a progressive loss of muscle mass and function associated with aging or immobility [1]. More specifically, sarcopenia is defined as low muscle strength, reduced quantity and quality of muscle mass, and decreased physical performance [2]. Stated another way, “Sarcopenia is a progressive, generalized skeletal muscle disorder involving the combination of loss of muscle mass and loss of muscle function and/or muscle strength, as well as loss of muscle performance” [3]. Age-related muscle wasting diseases can be categorized as either sarcopenia or cachexia. Cachexia is muscle wasting that is associated with disease processes such as cancer. Sarcopenia is muscle loss due to the aging process and is perhaps classified as lean or obese sarcopenia depending on the fat mass of the sarcopenic individual [4]. Sarcopenia appears to be inextricably intertwined with glucose metabolism, and the maintenance of muscle mass in addition to the balance of insulin sensitivity and insulin secretion plays a critical role in maintaining glucose homeostasis [1]. However, insulin resistance is modulated by complex interactions with various factors, including body composition, gut microbiota, and various nutrient intakes. Increasing insulin resistance due to aging, obesity, inflammation, and oxidative stress elevates insulin secretion, eventually leading to chronic hyperglycemia and decreased lifespan [5].
Many scientists have investigated which tissues are primarily responsible for insulin resistance. Skeletal muscle has been identified as one of the responsible target tissues. Skeletal muscle tissues are insulin-dependent, and they are the most significant contributor to insulin-dependent glucose disposal [5]. Consequently, decreased skeletal muscle mass and impaired muscle glucose metabolism are possible contributors to type 2 diabetes and vice versa [5][6]. Skeletal muscle is believed to begin during the third decade of life and gradually decreases the capacity for muscle glucose utilization, increasing susceptibility to type 2 diabetes [7]. Type 2 diabetes increases sarcopenia risk and has a bidirectional relationship [5]. The balance between anabolism and catabolism regulates skeletal muscle mass. Anabolism is stimulated by growth factors, including androgens, insulin-like growth factor-I (IGF-I), insulin, and some myokines such as irisin, myonectin, decorin, and fibroblast growth factor (FGF)-21 [8]. Catabolism-related signaling pathways counterbalance anabolism in response to stimuli such as glucocorticoids and proinflammatory cytokines (Figure 1) [9]. These endogenous factors influence the Akt/mammalian target of rapamycin (mTOR), SMAD, autophagy, and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling pathways to modulate mitochondriogenesis, myogenesis, and muscle atrophy [10]. Among those pathways, IGF-1/insulin signaling is directly involved in insulin sensitivity, and it indirectly affects the NF-κB signaling pathways to modulate skeletal muscle mass [11]. Therefore, the IGF-1/insulin and NF-κB signaling pathways are closely associated with maintaining skeletal muscle mass.
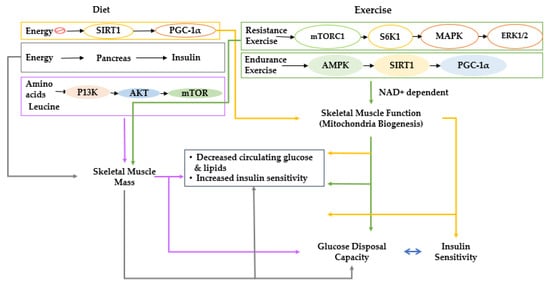
Figure 1. Proposed interactions of diet and lifestyle with skeletal muscle and glucose metabolism. Effects of diet and lifestyle on skeletal muscle mass and function and the impact on blood glucose regulation are explained; diet and lifestyle are shown to exert effects on muscle mass and function, which have interactive effects on glucose metabolism. Sirt 1, NAD-dependent deacetylase sirtuin-1; PGC-1α, peroxisome proliferator-activated receptor-gamma coactivator 1α; mTORC1, mechanistic target of rapamycin C1; S6K1, ribosomal protein S6 kinase beta-1; MAPK, mitogen-activated protein kinase; ERK1/2, extracellular signal-regulated kinases; AMPK, AMP-activated protein kinase; PI3K, phosphoinositide 3-kinase; AKT; NAD+, nicotinamide adenine dinucleotide.
2. Overall View of Muscle Metabolism and the Factors That Control Muscle Growth
As the population ages, there is an increasing burden of frailty characterized by bone loss, often resulting in osteoarthritis and fractures and muscle loss, sometimes progressing to a clinical diagnosis of sarcopenia [12]. The mechanical loads imposed on bone due to muscle contraction are critical for stimulating bone growth and maybe a propounding factor in developing degenerative bone diseases of aging as people become more sedentary [12]. The decreased physical activity is also believed to be an essential factor in the lifestyle changes that lead to sarcopenia [13]. Furthermore, it has been recently observed that low numbers of circulating osteoprogenitor cells, an indicator of mesenchymal stem cells in bone marrow, are associated with frailty and sarcopenia [14]. Therefore, it is likely that both muscle loss and bone loss exacerbate each other and are crucial factors in developing frailty during aging.
Skeletal muscle plays a critical role not only in glucose and lipid metabolism but also in endocrine and paracrine activities [15]. Genetic and environmental factors and disease status are involved in maintaining skeletal muscle mass, which interacts with various organs, including bone, adipose tissue, liver, heart, and brain [15][16]. Skeletal muscle mass is a net result of both catabolic and anabolic metabolisms of myocytes. Muscle loss is related to reduced satellite cell recruitment and anabolic hormonal signaling, protein oxidation, inflammation, and developmental factors [17]. Muscle breakdown is involved in elevated oxidative stress, degenerative neuromuscular junction, and hyperglycemia, in addition to muscle mass loss [17][18]. However, the molecular mechanisms have not been clearly characterized [16].
3. Age-Related Interactive Effects of Impaired Glucose Metabolism and Sarcopenia
3.1. Skeletal Muscle Mass Loss during Aging
Skeletal muscle mass has been reported to be positively associated with a metabolically healthy phenotype in a large population study [19]. The benefits were valid for both skeletal muscle mass and muscle quality and were associated with having less than two components of metabolic syndrome [19][20]. Unfortunately, one of the most consistent physiological features of the aging process is a gradual but progressive loss of skeletal muscle mass and function [13]. Age-related loss of skeletal muscle mass begins at around age 30 and is a consistent feature of the aging process [21]. However, the rapidity and severity of the progression of muscle are highly variable and may be modulated by dietary and lifestyle factors. It is often difficult to precisely define the degree of muscle loss and compare the differences among people due to how muscle loss presents among individuals and differences in assessment methods [22]. However, the most severe muscle loss is termed sarcopenia and has become officially recognized as a disease state in recent years, and was assigned an International Diagnostic Classification Code (ICD-10-CM code M62.84) [23].
The progression of muscle loss is usually accompanied by various metabolic changes that manifest during different stages of the aging process and may be involved in changes in body composition. Sarcopenia is strongly linked to metabolic diseases and frailty [24].
3.2. Sarcopenia and Glucose Metabolism
The factors responsible for age-related declines in muscle mass and function are not fully understood, but anabolic resistance seems to be one of the significant contributors to sarcopenia [25]. However, the nature of anabolic resistance is somewhat obscure, and there are profound differences of opinion about its causes and effects. Many investigators believe that the anabolic signals from the two major anabolic stimuli, resistance exercise, and amino acid (especially leucine) consumption, are suppressed [26] due to a lessened activation of phosphoinositide 3-kinase (PI3K) activation, in turn, decreased AKT signaling as well as mTOR. PI3K/AKT/mTOR signaling is a primary stimulatory regulator of anabolic signals for muscle growth and inhibitor of muscle anabolism (Figure 1) [26]. The impairment of anabolic signaling pathways is a commonly held paradigm of anabolic resistance in skeletal muscle. However, other researchers have demonstrated that older people do not have a blunted anabolic response to amino acids and resistance exercise [27][28].4. Glucose Metabolism and Gut Microbiota
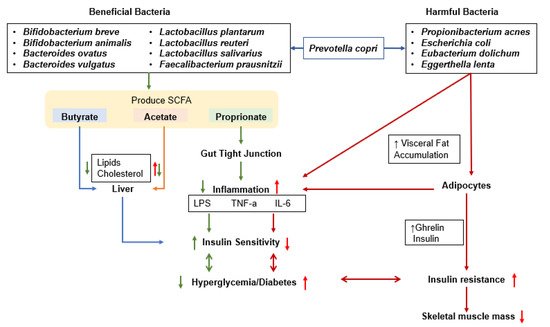
Genetic and environmental factors influence glucose metabolism, affecting both insulin secretion and insulin sensitivity [29][30]. Carbohydrate digestion and absorption, inflammation, nutrient intakes, and lifestyle factors such as alcohol consumption and exercise influence glucose homeostasis. The gut microbiome is an emerging factor that influences glucose metabolism by interacting with various factors and shows bidirectional symbiosis (Figure 2) [31]. Gut microbiota produces SCFA that may influence the host’s glucose metabolism by promoting the gut–liver, gut–skeletal muscle, gut–islet, and gut–brain axes [32]. These effects modulate insulin sensitivity, insulin secretion, and glucose disposal.Figure 2. Proposed interaction of gut bacteria with glucose metabolism. Effects of gut microbiota and their products on the liver, gut cell wall, adipocytes, and inflammation as regulators of glucose metabolism are explained; gut bacteria can increase and decrease inflammation, either improving or exacerbating insulin resistance and muscle loss. LPS, lipopolysaccharide; TNF-α, tumor necrosis factor-α; IL-6, interleukin-6.
Bacteria use glucose as an energy source, and glucose supply is a significant factor for commensalism with the host and bacteria colonization. Gut bacteria can be categorized as having mutualism, commensalism, and parasitism, all of which can affect the host’s glucose metabolism [33].
5. Effects of the Microbiome on Muscle Mass and the Development of Sarcopenia
5.1. Physical Activity, Sarcopenia, and Gut–Muscle Axis
The gut microbiome is a menagerie of bacteria, archaea, fungi, viruses, and eukaryotic microbes that live as independent life forms in humans and other mammalian hosts [34]. The microbiota making up the microbiome can profoundly affect the host, potentially affecting almost all physiological systems [35]. It seems counter-intuitive that the microorganisms living in the intestinal tract can regulate such seemingly disconnected systems as skeletal muscle mass and function. However, in a 2017 review article, Ticinesi et al. [36], outlined the current animal and human research supporting a bidirectional gut–muscle axis associated with sarcopenia. First of all, it had been well established that after age 65, there is a decline in the abundance and diversity of the gut microbiota, especially taxa believed to contribute health benefits [37]. Several studies have confirmed a strong association between exercise and fitness and a healthy robust microbiome, suggesting that exercise may stimulate the growth of healthy gut microbes in addition to muscle increment [38][39].
5.2. Gut Microbiota, Sarcopenia, and Gut–Muscle Axis
The changes in gut microbiota are involved with skeletal muscle mass by changing SCFA and inflammatory cytokines to modulate the gut–muscle axis. Qui et al. used an antibiotic cocktail to mostly eliminate the gut microbiota of 10-week-old C57BL/6 mice and determine the effects on body composition [40]. The antibiotic-treated mice experienced skeletal muscle atrophy linked to decreased ileal farnesoid X receptor (FXR)-FGF15 signaling and subsequently impaired skeletal muscle protein synthesis. Okamoto et al. investigated the effects of short-chain fatty acids produced by gut bacteria in mice [41]. They demonstrated that SCFA (acetate, butyrate, and propionate) were almost completely eliminated from fecal samples of antibiotic-treated mice, which had significantly decreased exercise endurance, but an infusion with acetate restored the exercise endurance in the mice.5.3. Myokines, Sarcopenia, and Gut–Liver–Muscle Axis
Myokines are released from myocytes in response to muscle contraction, and they are involved in muscle metabolism and other tissues, including adipocytes, liver, and brain, through their receptors [8]. Several known myokines, including myostatin, irisin, myonectin, FGF-21, decorin, IL-6, IL-15, and others, and their biological functions have not been well characterized. Some known characteristics involve myocyte proliferation, differentiation, growth, and atrophy. Irisin, myonectin, decorin, FGF-21, secreted protein acidic and cysteine-rich (SPARC), and brain-derived neurotrophic factor have a positive activity to increase muscle mass, but myostatin, IL-6, and IL-15 are involved in muscle atrophy [7]. Thus, the regulation of myokine activity can protect against sarcopenia. Gut microbiota is involved in regulating myokine function through changing SCFA, secondary bile acids, branched-chain amino acids (BCAA), endocannabinoids, and inflammatory cytokines [42].6. BCAA Effects on Metabolism and Sarcopenia Host through the Gut–Muscle Axis
Supplementing older adults with loss of muscle mass and function with BCAA and vitamin D for 8 weeks has improved muscle mass and function [43]. A Japanese study has found that supplementing BCAA to liver cirrhosis patients prevents sarcopenia and fat accumulation in skeletal muscle [44]. The apparent inconsistency between BCAA, insulin resistance, and type 2 diabetes may be related to BCAA utilization due to the attenuation of insulin resistance in the body. High serum BCAA concentrations are linked to insulin resistance, obesity, and type 2 diabetes, although BCAA supplementation is beneficial for increasing energy expenditure and skeletal muscle synthesis by activating mTOC1 [45][46][47]. Brown adipose tissues utilize BCAA catabolism in the mitochondria for thermogenesis, increasing energy expenditure [46]. However, a defect in the BCAA clearance mediated by SLC25A44 attenuates BCAA clearance to increase serum BACC concentration, suggesting inducing insulin resistance [46]. Gut microbiota is closely related to BCAA metabolism and is involved in their synthesis and degradation; however, their imbalance by gut microbiota raises serum BCAA concentrations, contributing to increased insulin resistance [45].7. The Modulation of Dietary Intake and Lifestyles and Gut Microbiome to Promote Skeletal Muscle Mass and Prevent Sarcopenia
7.1. Calorie and Fat Intake, Gut Microbiota, and Skeletal Muscle Mass
Research associating dietary intake with specific changes in the gut microbiota often produces inconsistent results. However, current research suggests that a dietary program that increases microbial diversity in the gut, particularly when rich in species that produce SCFA, is highly beneficial for healthy skeletal muscle aging [48]. Various dietary factors, including intakes of prebiotics, fermented foods, fat, protein, and carbohydrates, can modulate gut microbiota composition [49]. The changes in gut microbiota are associated with not only the availability of energy sources but also intestinal permeability, digestion capacity of carbohydrates and proteins, gastric acid, and bile acid secretion [50]. Muscle loss is linked to insulin resistance and inflammation associated with gut microbiota dysbiosis, contributing to increased intestinal barrier permeability, serum inflammatory cytokines, and insulin resistance [51].
7.2. Probiotic and Prebiotic Intakes, Gut Microbiota, and Skeletal Muscle Mass
Skeletal muscle mass and strength decrease from middle age to the elderly, and it is difficult to prevent the decrement of muscle mass and strength only with dietary intake. The effects of protein and amino acids, especially leucine, vitamin D, ω-3 fatty acids, antioxidants, magnesium, and probiotics, have been studied as nutritional interventions to prevent sarcopenia in the elderly [52]. However, their preventive activities are minimal, but supplementation with exercise may be beneficial [52].8. Summary and Conclusions
Age-associated physiological changes commonly include losses of muscle mass and function, declines in microbiome quality and diversity, and disruption of glucose regulation. The simultaneous occurrence of these changes is probably not coincidental since interactions among them can cause many of the underlying defects associated with each, and dietary and lifestyle changes that occur with aging can also contribute to each condition. Muscle mass and function can be improved by physiological stimuli such as load-bearing exercise, protein consumption, especially the amino acid leucine, and improving anabolic hormone signaling, including insulin and SIRT-1. Insulin resistance and inflammation are linked to muscle mass loss, but it is not yet known if loss of muscle mass is a consequence or cause of insulin resistance and/or inflammation. However, increased circulating lipids due to decreased uptake in muscle could lead to insulin resistance, and insulin resistance in muscle could decrease anabolic signaling by insulin/IGF-1. Increased proinflammatory cytokines reduce PGC-1α activation, increasing oxidative stress and NF-κB activation to elevate further inflammation linked to muscle degradation. The gut microbiome is also associated with increasing and maintaining muscle mass through modulating the gut microbiome-muscle axis by producing SCFAs and proinflammatory cytokines. Gut dysbiosis induces a leaky intestinal wall that allows toxic bacterial metabolites to enter the host metabolism. SCFA, especially propionate and butyrate, may act as AMPK activator to stimulate PGC-1α and insulin/IGF-1 signaling to promote muscle biogenesis.
Human aging is associated with impaired glucose utilization, decreased muscle mass and function, and lessened gut microbial diversity. There is increasing evidence that muscle mass decrement is accelerated by its interactions with dietary and lifestyle changes that impact it. It has been demonstrated that muscle mass and function remain near youthful levels with both load-bearing and endurance exercise as well as adequate intakes of protein and specific amino acids. Optimal muscle function improves glucose disposal in the muscle, which assists glucose management, as do signals from the gut microbiome. Finally, non-digestible carbohydrate intake results in SCFA production that stimulates muscle growth and improves glucose regulation. Therefore, this entry provides convincing evidence that a concerted program of specific dietary and lifestyle interventions can have synergistic effects on multiple mutually supportive physiological systems that may delay age-related physical deterioration and prevent sarcopenia in aging adults.
References
- Peterson, S.J.; Mozer, M. Differentiating Sarcopenia and Cachexia Among Patients With Cancer. Nutr. Clin. Pract. 2017, 32, 30–39.
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31.
- Lampignano, L.; Bortone, I.; Castellana, F.; Donghia, R.; Guerra, V.; Zupo, R.; De Pergola, G.; Di Masi, M.; Giannelli, G.; Lozupone, M.; et al. Impact of Different Operational Definitions of Sarcopenia on Prevalence in a Population-Based Sample: The Salus in Apulia Study. Int. J. Environ. Res. Public Health 2021, 18, 12979.
- Molino, S.; Dossena, M.; Buonocore, D.; Verri, M. Sarcopenic Obesity: An Appraisal of the Current Status of Knowledge and Management in Elderly People. J. Nutr. Health Aging 2016, 20, 780–788.
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161.
- Mesinovic, J.; Zengin, A.; De Courten, B.; Ebeling, P.R.; Scott, D. Sarcopenia and type 2 diabetes mellitus: A bidirectional relationship. Diab. Metab. Syndr. Obes. 2019, 12, 1057–1072.
- Mitchell, W.K.; Williams, J.; Atherton, P.; Larvin, M.; Lund, J.; Narici, M. Sarcopenia, dynapenia, and the impact of advancing age on human skeletal muscle size and strength; a quantitative review. Front. Physiol. 2012, 3, 260.
- Lee, J.H.; Jun, H.-S. Role of Myokines in Regulating Skeletal Muscle Mass and Function. Front. Physiol. 2019, 10, 42.
- Klein, G.L. The effect of glucocorticoids on bone and muscle. Osteoporos. Sarcopenia 2015, 1, 39–45.
- Ji, L.L.; Yeo, D. Mitochondrial dysregulation and muscle disuse atrophy. F1000Research 2019, 8, F1000.
- Schakman, O.; Dehoux, M.; Bouchuari, S.; Delaere, S.; Lause, P.; Decroly, N.; Shoelson, S.E.; Thissen, J.P. Role of IGF-I and the TNFα/NF-κB pathway in the induction of muscle atrogenes by acute inflammation. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E729–E739.
- Novotny, S.A.; Warren, G.L.; Hamrick, M.W. Aging and the muscle-bone relationship. Physiology 2015, 30, 8–16.
- Nair, K.S. Aging muscle. Am. J. Clin. Nutr. 2005, 81, 953–963.
- Al Saedi, A.; Phu, S.; Vogrin, S.; Gunawardene, P.; Duque, G. Association between Circulating Osteoprogenitor Cells and Sarcopenia. Gerontology 2021, 1–6.
- Kim, G.; Kim, J.H. Impact of Skeletal Muscle Mass on Metabolic Health. Endocrinol. Metab. 2020, 35, 1–6.
- Park, S. Association between polygenetic risk scores related to sarcopenia risk and their interactions with regular exercise in a large cohort of Korean adults. Clin. Nutr. 2021, 40, 5355–5364.
- Tournadre, A.; Vial, G.; Capel, F.; Soubrier, M.; Boirie, Y. Sarcopenia. Jt. Bone Spine 2019, 86, 309–314.
- Romanello, V.; Sandri, M. Mitochondrial qaulity control and muscle mass maintenance. Front. Physiol. 2016, 6, 422.
- Lee, M.J.; Kim, E.-H.; Bae, S.-J.; Choe, J.; Jung, C.H.; Lee, W.J.; Kim, H.-K. Protective role of skeletal muscle mass against progression from metabolically healthy to unhealthy phenotype. Clin. Endocrinol. 2019, 90, 102–113.
- Kim, H.K.; Lee, M.J.; Kim, E.H.; Bae, S.J.; Kim, K.W.; Kim, C.H. Comparison of muscle mass and quality between metabolically healthy and unhealthy phenotypes. Obesity 2021, 29, 1375–1386.
- Volpi, E.; Nazemi, R.; Fujita, S. Muscle tissue changes with aging. Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 405–410.
- Bosy-Westphal, A.; Müller, M.J. Identification of skeletal muscle mass depletion across age and BMI groups in health and disease--there is need for a unified definition. Int. J. Obes. 2015, 39, 379–386.
- Anker, S.D.; Morley, J.E.; von Haehling, S. Welcome to the ICD-10 code for sarcopenia. J. Cachexia Sarcopenia Muscle 2016, 7, 512–514.
- Buscemi, C.; Ferro, Y.; Pujia, R.; Mazza, E.; Boragina, G.; Sciacqua, A.; Piro, S.; Pujia, A.; Sesti, G.; Buscemi, S.; et al. Sarcopenia and Appendicular Muscle Mass as Predictors of Impaired Fasting Glucose/Type 2 Diabetes in Elderly Women. Nutrients 2021, 13, 1909.
- Haran, P.H.; Rivas, D.A.; Fielding, R.A. Role and potential mechanisms of anabolic resistance in sarcopenia. J. Cachexia Sarcopenia Muscle 2012, 3, 157–162.
- Lee, E.J.; Neppl, R.L. Influence of Age on Skeletal Muscle Hypertrophy and Atrophy Signaling: Established Paradigms and Unexpected Links. Genes 2021, 12, 688.
- Moro, T.; Brightwell, C.R.; Deer, R.R.; Graber, T.G.; Galvan, E.; Fry, C.S.; Volpi, E.; Rasmussen, B.B. Muscle Protein Anabolic Resistance to Essential Amino Acids Does Not Occur in Healthy Older Adults Before or After Resistance Exercise Training. J. Nutr. 2018, 148, 900–909.
- Paulussen, K.J.M.; McKenna, C.F.; Beals, J.W.; Wilund, K.R.; Salvador, A.F.; Burd, N.A. Anabolic Resistance of Muscle Protein Turnover Comes in Various Shapes and Sizes. Front. Nutr. 2021, 8, 615849.
- Dendup, T.; Feng, X.; Clingan, S.; Astell-Burt, T. Environmental Risk Factors for Developing Type 2 Diabetes Mellitus: A Systematic Review. Int. J. Environ. Res. Public Health 2018, 15, 78.
- Khodaeian, M.; Enayati, S.; Tabatabaei-Malazy, O.; Amoli, M.M. Association between Genetic Variants and Diabetes Mellitus in Iranian Populations: A Systematic Review of Observational Studies. J. Diabetes Res. 2015, 2015, 585917.
- Yu, L.; Zhou, X.; Duan, H.; Chen, Y.; Cui, S.; Guo, R.; Xue, Y.; Tian, F.; Zhao, J.; Zhang, H.; et al. Synergistic Protective Effects of Different Dietary Supplements Against Type 2 Diabetes via Regulating Gut Microbiota. J. Med. Food 2021, 24, 319–330.
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206.
- Anhê, F.F.; Barra, N.G.; Schertzer, J.D. Glucose alters the symbiotic relationships between gut microbiota and host physiology. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E111–E116.
- Berg, G.; Rybakova, D.; Fischer, D.; Cernava, T.; Vergès, M.-C.C.; Charles, T.; Chen, X.; Cocolin, L.; Eversole, K.; Corral, G.H.; et al. Microbiome definition re-visited: Old concepts and new challenges. Microbiome 2020, 8, 103.
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75.
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging Gut Microbiota at the Cross-Road between Nutrition, Physical Frailty, and Sarcopenia: Is There a Gut-Muscle Axis? Nutrients 2017, 9, 1303.
- Ragonnaud, E.; Biragyn, A. Gut microbiota as the key controllers of “healthy” aging of elderly people. Immun. Ageing 2021, 18, 2.
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920.
- Schroeder, B.O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089.
- Qiu, Y.; Yu, J.; Li, Y.; Yang, F.; Yu, H.; Xue, M.; Zhang, F.; Jiang, X.; Ji, X.; Bao, Z. Depletion of gut microbiota induces skeletal muscle atrophy by FXR-FGF15/19 signalling. Ann. Med. 2021, 53, 508–522.
- Okamoto, T.; Morino, K.; Ugi, S.; Nakagawa, F.; Lemecha, M.; Ida, S.; Ohashi, N.; Sato, D.; Fujita, Y.; Maegawa, H. Microbiome potentiates endurance exercise through intestinal acetate production. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E956–E966.
- Suriano, F.; Van Hul, M.; Cani, P.D. Gut microbiota and regulation of myokine-adipokine function. Curr. Opin. Pharmacol. 2020, 52, 9–17.
- Gkekas, N.K.; Anagnostis, P.; Paraschou, V.; Stamiris, D.; Dellis, S.; Kenanidis, E.; Potoupnis, M.; Tsiridis, E.; Goulis, D.G. The effect of vitamin D plus protein supplementation on sarcopenia: A systematic review and meta-analysis of randomized controlled trials. Maturitas 2021, 145, 56–63.
- Tajiri, K.; Shimizu, Y. Branched-chain amino acids in liver diseases. Transl. Gastroenterol. Hepatol. 2018, 3, 47.
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381.
- Yoneshiro, T.; Wang, Q.; Tajima, K.; Matsushita, M.; Maki, H.; Igarashi, K.; Dai, Z.; White, P.J.; McGarrah, R.W.; Ilkayeva, O.R.; et al. BCAA catabolism in brown fat controls energy homeostasis through SLC25A44. Nature 2019, 572, 614–619.
- Kamei, Y.; Hatazawa, Y.; Uchitomi, R.; Yoshimura, R.; Miura, S. Regulation of Skeletal Muscle Function by Amino Acids. Nutrients 2020, 12, 261.
- Ticinesi, A.; Mancabelli, L.; Tagliaferri, S.; Nouvenne, A.; Milani, C.; Del Rio, D.; Lauretani, F.; Maggio, M.G.; Ventura, M.; Meschi, T. The Gut-Muscle Axis in Older Subjects with Low Muscle Mass and Performance: A Proof of Concept Study Exploring Fecal Microbiota Composition and Function with Shotgun Metagenomics Sequencing. Int. J. Mol. Sci. 2020, 21, 8946.
- Jeong, D.Y.; Ryu, M.S.; Yang, H.J.; Park, S. γ-PGA-Rich Chungkookjang, Short-Term Fermented Soybeans: Prevents Memory Impairment by Modulating Brain Insulin Sensitivity, Neuro-Inflammation, and the Gut-Microbiome-Brain Axis. Foods 2021, 10, 221.
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 625913.
- Ticinesi, A.; Nouvenne, A.; Cerundolo, N.; Catania, P.; Prati, B.; Tana, C.; Meschi, T. Gut Microbiota, Muscle Mass and Function in Aging: A Focus on Physical Frailty and Sarcopenia. Nutrients 2019, 11, 1633.
- Cruz-Jentoft, A.J.; Dawson Hughes, B.; Scott, D.; Sanders, K.M.; Rizzoli, R. Nutritional strategies for maintaining muscle mass and strength from middle age to later life: A narrative review. Maturitas 2020, 132, 57–64.
More