The vines and leaves of Momordica charantia L. are used as herbal medicines to treat inflammation-related disorders. However, their safety profile remains uncharacterized, and the constituents in their extracts that exert anti-inflammatory and adverse effects remain unclear. This study isolated the characteristic cucurbitane-type triterpenoid species in the vines and leaves of M. charantia L. and analyzed their cytotoxicity, anti-inflammatory effects, and underlying mechanisms. Four structurally related triterpenoids—momordicines I, II, IV, and (23E) 3β,7β,25-trihydroxycucurbita-5,23-dien-19-al (TCD)—were isolated from the triterpenoid-rich fractions of extracts from the vines and leaves of M. charantia. Momordicine I was cytotoxic on normal cells, momordicine II exerted milder cytotoxicity, and momordicine IV and TCD had no obvious adverse effects on cell growth. TCD had anti-inflammatory activity both in vivo and in vitro. In lipopolysaccharide-stimulated RAW 264.7 cells, TCD inhibited the inhibitor kappa B kinase/nuclear factor-κB pathway and enhanced the expression of nuclear factor erythroid 2-related factor 2, heme oxygenase-1, and glutamate-cysteine ligase modifier subunit through the extracellular signal-regulated kinase1/2 and p38. Thus, the vines and leaves of M. charantia should be used with caution. An extraction protocol that can enrich TCD but remove momordicine I would likely enhance the safety of the extract.
- Momordica charantia
- triterpenoid
- anti-inflammation
- antioxidant enzymes
- Nrf2
- heme oxygenase-1
- cytotoxicity
1. Introduction
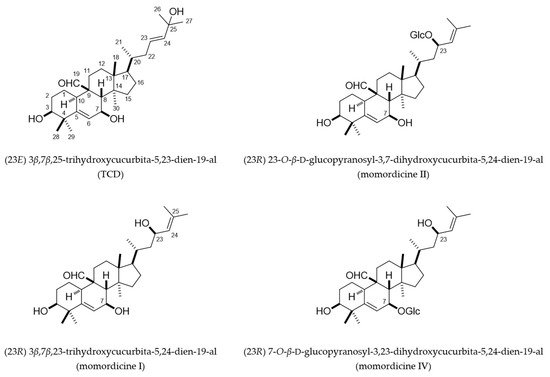
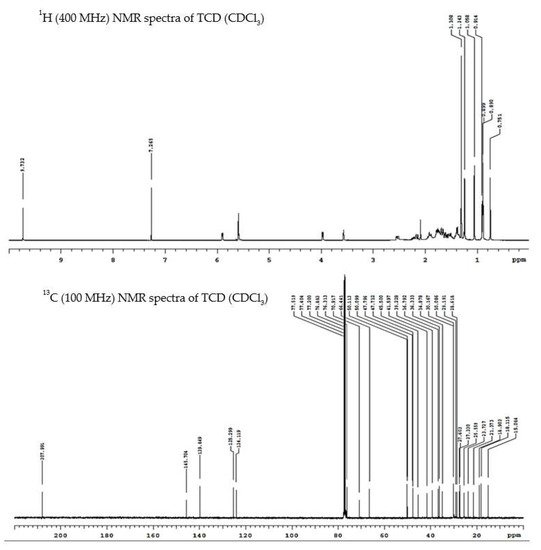
2. Cucurbitane-Type Triterpenoids Isolated from the Vines and Leaves of M. charantia
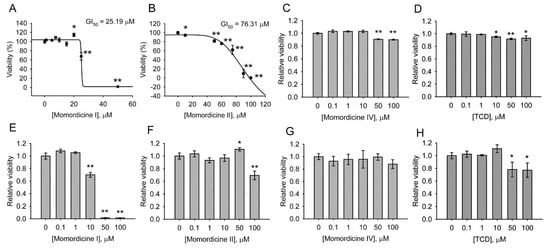
3. Cytotoxicity of Triterpenoids
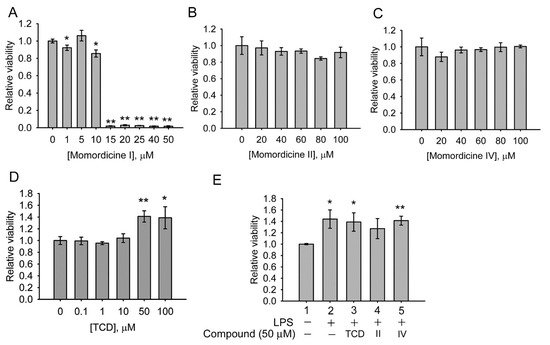
4. Anti-Inflammatory Effects of Triterpenoids
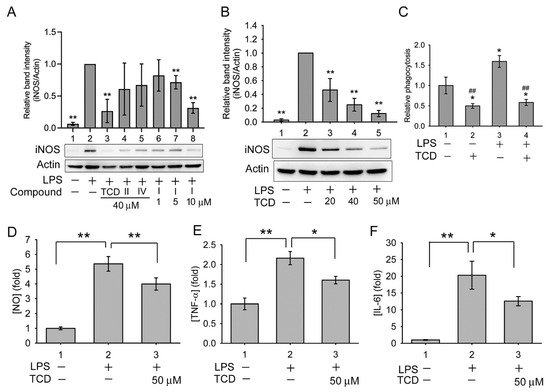
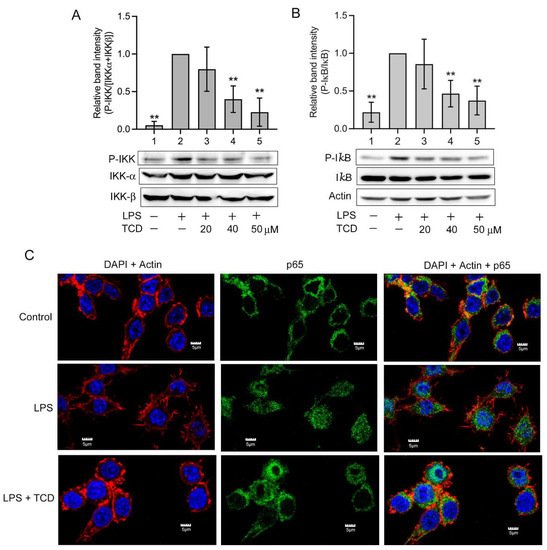
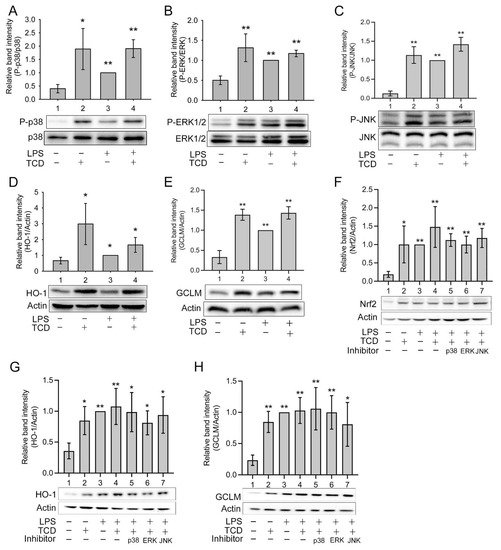
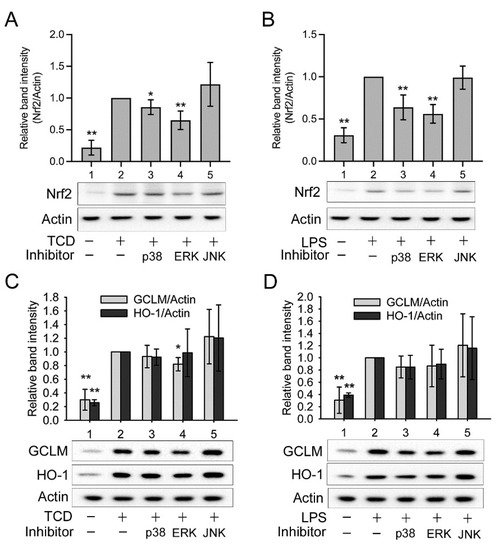
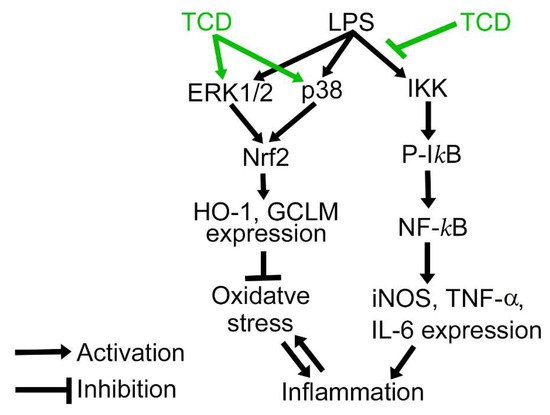
5. Anti-Inflammatory Mechanism of TCD
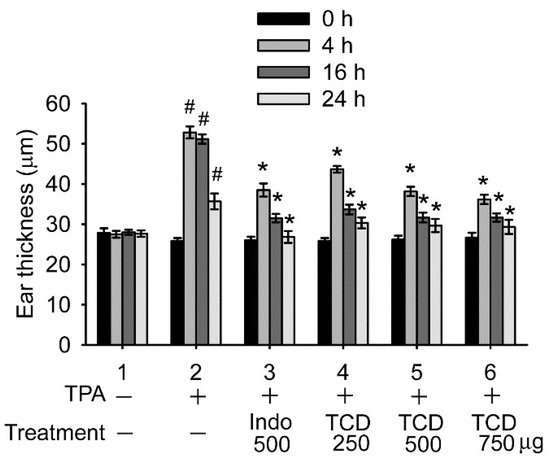
6. Discussion
References
- Jia, S.; Shen, M.; Zhang, F.; Xie, J. Recent advances in Momordica charantia: Functional components and biological activities. Int. J. Mol. Sci. 2017, 18, 2555.
- Bortolotti, M.; Mercatelli, D.; Polito, L. Momordica charantia, a nutraceutical approach for inflammatory related diseases. Front. Pharmacol. 2019, 10, 486.
- Minihane, A.M.; Vinoy, S.; Russell, W.R.; Baka, A.; Roche, H.M.; Tuohy, K.M.; Teeling, J.L.; Blaak, E.E.; Fenech, M.; Vauzour, D.; et al. Low-grade inflammation, diet composition and health: Current research evidence and its translation. Br. J. Nutr. 2015, 114, 999–1012.
- Chang, C.I.; Chen, C.R.; Liao, Y.W.; Cheng, H.L.; Chen, Y.C.; Chou, C.H. Cucurbitane-type triterpenoids from Momordica charantia. J. Nat. Prod. 2006, 69, 1168–1171.
- Cheng, H.L.; Huang, H.K.; Chang, C.I.; Tsai, C.P.; Chou, C.H. A cell-based screening identifies compounds from the stem of Momordica charantia that overcome insulin resistance and activate AMP-activated protein kinase. J. Agric. Food Chem. 2008, 56, 6835–6843.
- Nagarani, G.; Abirami, A.; Siddhuraju, P. Food prospects and nutraceutical attributes of Momordica species: A potential tropical bioresources—a review. Food Sci. Hum. Well. 2014, 3, 117–126.
- Yasuda, M.; Iwamoto, M.; Okabe, H.; Yamauchi, T. Structures of Momordicines I, II and III, the bitter principles in the leaves and vines of Momordica charantia L. Chem. Parm. Bull. 1984, 32, 2044–2047.
- Fatope, M.O.; Takeda, Y.; Yamashita, H.; Okabe, H.; Yamauchi, T. New cucurbitane triterpenoids from Momordica-Charantia. J. Nat. Prod. 1990, 53, 1491–1497.
- Mekuria, D.B.; Kashiwagi, T.; Tebayashi, S.; Kim, C.S. Cucurbitane glucosides from Momordica charantia leaves as oviposition deterrents to the leafminer, Liriomyza trifolii. Z. Naturforsch. C 2006, 61, 81–86.
- Mohan, S.; Gupta, D. Crosstalk of toll-like receptors signaling and Nrf2 pathway for regulation of inflammation. Biomed. Pharmacother. 2018, 108, 1866–1878.
- Tajalli-Nezhad, S.; Karimian, M.; Beyer, C.; Atlasi, M.A.; Azami Tameh, A. The regulatory role of Toll-like receptors after ischemic stroke: Neurosteroids as TLR modulators with the focus on TLR2/4. Cell. Mol. Life Sci. 2019, 76, 523–537.
- Liu, L.; Guo, H.; Song, A.; Huang, J.; Zhang, Y.; Jin, S.; Li, S.; Zhang, L.; Yang, C.; Yang, P. Progranulin inhibits LPS-induced macrophage M1 polarization via NF-κB and MAPK pathways. BMC Immunol. 2020, 21, 32.
- Gong, J.; Li, J.; Dong, H.; Chen, G.; Qin, X.; Hu, M.; Yuan, F.; Fang, K.; Wang, D.; Jiang, S.; et al. Inhibitory effects of berberine on proinflammatory M1 macrophage polarization through interfering with the interaction between TLR4 and MyD88. BMC Complement. Altern. Med. 2019, 19, 314.
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and -independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717.
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2-an update. Free Radic. Biol. Med. 2014, 66, 36–44.
- Yu, C.; Xiao, J.H. The Keap1-Nrf2 system: A mediator between oxidative stress and aging. Oxid. Med. Cell Longev. 2021, 2021, 6635460.
- Rushworth, S.A.; MacEwan, D.J.; O'Connell, M.A. Lipopolysaccharide-induced expression of NAD(P)H:quinone oxidoreductase 1 and heme oxygenase-1 protects against excessive inflammatory responses in human monocytes. J. Immunol. 2008, 181, 6730–6737.
- Tran, P.L.; Tran, P.T.; Tran, H.N.K.; Lee, S.; Kim, O.; Min, B.S.; Lee, J.H. A prenylated flavonoid, 10-oxomornigrol F, exhibits anti-inflammatory effects by activating the Nrf2/heme oxygenase-1 pathway in macrophage cells. Int. Immunopharmacol. 2018, 55, 165–173.
- Paine, A.; Eiz-Vesper, B.; Blasczyk, R.; Immenschuh, S. Signaling to heme oxygenase-1 and its anti-inflammatory therapeutic potential. Biochem. Pharmacol. 2010, 80, 1895–1903.
- Yang, C.H.; Yen, T.L.; Hsu, C.Y.; Thomas, P.A.; Sheu, J.R.; Jayakumar, T. Multi-targeting andrographolide, a novel NF-kappaB inhibitor, as a potential therapeutic agent for stroke. Int. J. Mol. Sci. 2017, 18, 1638.
- Xu, Y.; Yuan, H.; Luo, Y.; Zhao, Y.J.; Xiao, J.H. Ganoderic acid D protects human amniotic mesenchymal stem cells against oxidative stress-induced senescence through the PERK/NRF2 signaling pathway. Oxid. Med. Cell. Longev. 2020, 2020, 8291413.
- Song, C.; Heping, H.F.; Shen, Y.S.; Jin, S.X.; Li, D.Y.; Zhang, A.G.; Ren, X.L.; Wang, K.L.; Zhang, L.; Wang, J.D.; et al. AMPK/p38/Nrf2 activation as a protective feedback to restrain oxidative stress and inflammation in microglia stimulated with sodium fluoride. Chemosphere 2020, 244, 125495.
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Markelova, N.Y.; Penkov, N.V.; Shakurova, E.R.; Belosludtsev, K.N.; Parfenova, L.V. New quaternized pyridinium derivatives of betulin: Synthesis and evaluation of membranotropic properties on liposomes, pro- and eukaryotic cells, and isolated mitochondria. Chem. Biol. Interact. 2021, 349, 109678.
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Penkov, N.V.; Nedopekina, D.A.; Sharapov, V.A.; Khoroshavina, E.I.; Davletshin, E.V.; Belosludtseva, N.V.; Spivak, A.Y.; et al. Mitochondria-targeted prooxidant effects of betulinic acid conjugated with delocalized lipophilic cation F16. Free Radic. Biol. Med. 2021, 168, 55–69.
- Dubinin, M.V.; Semenova, A.A.; Ilzorkina, A.I.; Mikheeva, I.B.; Yashin, V.A.; Penkov, N.V.; Vydrina, V.A.; Ishmuratov, G.Y.; Sharapov, V.A.; Khoroshavina, E.I.; et al. Effect of betulin and betulonic acid on isolated rat liver mitochondria and liposomes. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183383.
- Javadov, S.; Jang, S.; Agostini, B. Crosstalk between mitogen-activated protein kinases and mitochondria in cardiac diseases: Therapeutic perspectives. Pharmacol. Therapeut. 2014, 144, 202–225.
- Belosludtsev, K.N.; Belosludtseva, N.V.; Kosareva, E.A.; Talanov, E.Y.; Gudkov, S.V.; Dubinin, M.V. Itaconic acid impairs the mitochondrial function by the inhibition of complexes II and IV and induction of the permeability transition pore opening in rat liver mitochondria. Biochimie 2020, 176, 150–157.
- Kodr, D.; Stankova, J.; Rumlova, M.; Dzubak, P.; Rehulka, J.; Zimmermann, T.; Krizova, I.; Gurska, S.; Hajduch, M.; Drasar, P.B.; et al. Betulinic acid decorated with polar groups and blue emitting BODIPY dye: Synthesis, cytotoxicity, cell-cycle analysis and anti-HIV profiling. Biomedicines 2021, 9, 1104.
- Kimura, O.; Ohta, C.; Koga, N.; Haraguchi, K.; Kato, Y.; Endo, T. Carrier-mediated uptake of nobiletin, a citrus polymethoxyflavonoid, in human intestinal Caco-2 cells. Food Chem. 2014, 154, 145–150.
- Harinantenaina, L.; Tanaka, M.; Takaoka, S.; Oda, M.; Mogami, O.; Uchida, M.; Asakawa, Y. Momordica charantia constituents and antidiabetic screening of the isolated major compounds. Chem. Pharm. Bull. 2006, 54, 1017–1021.
- Chang, C.I.; Chou, C.H.; Liao, M.H.; Chen, T.M.; Cheng, C.H.; Anggriani, R.; Tsai, C.P.; Tseng, H.I.; Cheng, H.L. Bitter melon triterpenes work as insulin sensitizers and insulin substitutes in insulin-resistant cells. J. Funct. Foods 2015, 13, 214–224.
- Tsai, T.H.; Huang, W.C.; Ying, H.T.; Kuo, Y.H.; Shen, C.C.; Lin, Y.K.; Tsai, P.J. Wild bitter melon leaf extract inhibits Porphyromonas gingivalis-induced inflammation: Identification of active compounds through bioassay-guided isolation. Molecules 2016, 21, 454.
- Chuang, L.T.; Huang, W.C.; Hou, Y.C.; Chyuan, J.H.; Chang, H.; Chang, C.I.; Tsai, T.H.; Tsai, P.J. Suppressive effect of two cucurbitane-type triterpenoids from Momordica charantia on Cutibacterium acnes-induced inflammatory responses in human THP-1 monocytic cell and mouse models. Molecules 2021, 26, 579.