Antibody ribosome display remains one of the most successful in vitro selection technologies for antibodies fifteen years after it was developed. The unique possibility of direct generation of whole proteins, particularly single-chain antibody fragments (scFvs), has facilitated the establishment of this technology as one of the foremost antibody production methods. Ribosome display has become a vital tool for efficient and low-cost production of antibodies for diagnostics due to its advantageous ability to screen large libraries and generate binders of high affinity. The remarkable flexibility of this method enables its applicability to various platforms.
- ribosome display
- • combinatorial libraries
- • biopanning
- • selection
- scFv
- diagnostics
- therapeutics
1. Introduction
2. In Vitro Ribosome Display
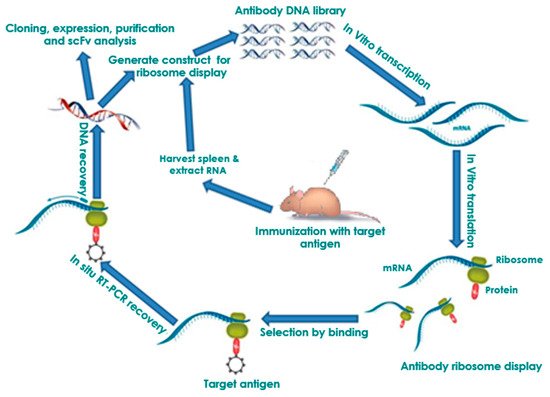
Ribosome display technology (RDT) is a potent in in vitrovitro, cell-free system that overcomes many limitations of cell-based methods by producing in vitro protein–mRNA complexes. There are several advantages of ribosome display compared to cell-based methods. First, the method is more efficient in the screening of large libraries without compromising the library size by transformation efficiency, selecting high-affinity combining sites, and eukaryotic cell-free systems, which are capable of post-translational modifications. Furthermore, it is quick and efficient as no cell culture is involved [13][14]. On the other hand, a noted limitation in ribosome display is the accessible, functional ribosome levels in the reaction for the library, which depends on the library size. RDT produces stable protein (antibody)–ribosome–mRNA (PRM) complexes to link individual antibody fragments to their corresponding mRNA [15]. The PRM complexes are formed through the deletion of the terminal stop codon from the mRNA, which causes stalling of the translating ribosome at the end of mRNA with the nascent polypeptide not released. The protein–mRNA linkage allows the simultaneous isolation of the mRNA and desirable proteins (antibodies) through an affinity for an immobilized ligand. The protein–mRNA complex that binds tightly to the ligand is subjected to in situ reverse transcription-PCR (RT-PCR) to recover the DNA encoding protein sequence [16] and amplified in a PCR reaction to generate a template for further manipulation and protein expression or panned for 3–5 additional cycles to obtain antibody leads. RDT allows the screening of libraries, with up to 1012–15-member antibody libraries in a single reaction [17]. Ribosome display has been extensively used in eukaryotic and prokaryotic translation systems. This was first demonstrated through a selection of peptide ligands using an E. coli extract by Mattheakis et al. [9][18]. This group showed the selection of peptide ligands that are similar to known peptide epitopes of a given antibody, using the antibody as a selection substrate. The peptide ligands of high-affinity were bound to the prostate-specific antigen and were identified through polysome selection from peptide libraries using a wheat germ extract translation system [19]. The selection of functional antibody fragments was reported using an E. coli translation system designed to increase the yield of ternary complexes and to allow disulfide bond formation [410]. This experimental set up was used to select antibodies from a murine library, and it was shown that the maturation of affinity occurs during the selection process. This is due to the combined effect of PCR errors and selection. An scFv fragment with a dissociation constant of about 10−11 M was obtained [20]. Specific antibody enrichment from mixed populations using rabbit reticulocyte extracts has also been demonstrated [410][21]. There was another study where scFv–ribosome–mRNA complexes were produced using a rabbit reticulocyte lysate system. This was then panned against the terminal protein (TP)-peptide of hepatitis B virus (HBV) DNA polymerase [22]. He and Taussig [14][16][23] also described the step-by-step procedure to perform eukaryotic ribosome display methodology. This has the distinctive feature of an in-situ RT-PCR procedure for DNA recovery from ribosome-bound mRNA. Another group [24] reported that a pseudoknot (originating from the genomic RNA of infectious bronchitis virus (IBV), a member of the positive-stranded coronavirus group) improves the selection efficiency in eukaryotic rabbit reticulocyte ribosome display. Qi et al. [25] selected antisulfadimidine-specific scFvs from a hybridoma cell through eukaryotic ribosome display. Kastelic and He [26] described the ribosome display of antibodies through the use of a eukaryotic rabbit reticulocyte system with an in-situ single-primer DNA recovery method. Edwards and He [27] also described the use of the eukaryotic rabbit reticulocyte ribosome display method to isolate variants of V(H) antibody fragments with improved affinities. Douthwaite [28] developed an optimized methodology for the use of rabbit reticulocyte lysate for ribosome display selections. Tang et al. [29] validated a novel in-vitro method for the rapid generation of human scFv monoclonal antibodies against recombinant gp120K530 from patient libraries using eukaryotic ribosome display.2.1. Selection of Antibodies by Panning
2.2. Affinity Maturation and Modification of Ribosome Display Antibodies
2.3. Ribosome Display Antibody Gene Libraries
3. Ribosome Display Technology in Disease Diagnostics and Control
Production of antibodies, especially scFvs, has been dramatically accelerated by in-vitro selection systems such as ribosome display technology. Within the past two decades, antibodies have gradually become very essential tools in the fields of biological sciences, agriculture, and medicine for basic research, disease diagnostics, and therapy.3.1. Human Infectious Diseases
Significant spike mutations (D614G, E484K, K417N/T, N501Y, L452R, T478K) are found associated with different clinical consequences throughout the globe [1104][110]. Scientists observed successful therapeutics from the significant clinical trials, including small antiviral molecules such as remdesivir or antibody-based therapeutics against SARS-CoV-2 [1115][111]. Several antibodies have shown significant neutralization activity against the virus. Some antibodies have received EUA (Emergency Use Authorization) for the treatment of this virus. Most of the antibodies are designed against the S-glycoprotein of this virus. Therefore, any S-glycoprotein mutations can trigger the antibody escapes/antibody resistance in SARS-CoV-2 variants and hinder the antibody-based therapeutic strategies against the virus [1104][110]. To avoid escape mutants, ribosome display technology could possibly be applied to target highly conserved epitopes to produce neutralizing scFv antibody cocktails targeting simultaneously non-RBD and RBD epitopes. Neutralizing antibody cocktails against SARS-CoV-2 would help in the development of diagnostics and treatment for COVID-19.
3.2. Cancer
3.3. Acquired Immunodeficiency Syndrome (AIDS)
3.4. Plant Disease: Pierce’s Disease
3.5. Pain
scFv antibodies are opening a new era of therapeutics, pharmacology, and pathophysiology research [1536][153]. These technologies have overcome previous challenges of providing therapeutic applications for G-protein-coupled receptors (GPCRs). More importantly, these small, brain penetrant antibodies are praised as having promising biotherapeutic applications for the nervous and immune systems, now recognized as interactive in chronic pain. scFvs are being investigated as therapeutics for arthritis, Creutzfeldt-Jakob, and Huntington’s disease due to their solubility, small size, and ability to cross the blood-brain barrier compared to mAbs available for migraine (Galcanezumab, Erenumab) [1547][1558][1569][154-156]. Despite the popularity of scFvs generated by ribosome display for chemotherapy, obtaining high-affinity scFvs from ribosome display libraries remains a challenging task [410][157].
Chronic pain frequently evokes anxiety, depression, disability, and diminishes quality of life. It is known that cholecystokinin (CCK) evokes anxiety/panic attacks in healthy subjects depending on dosage, and it is 103 times more abundant than any other neuropeptide in the nervous system. Selective antagonists of the CCKB receptor (CCKBR) enhance morphine analgesia and prevent tolerance without worsening respiratory depression in non-human primates or side effects other than orthostatic dizziness in placebo-controlled trials. We have generated a scFv biological that targets mouse CCKBR using ribosome display [15711][158]. The small CCKBR scFv is ~1/6 the size of a monoclonal antibody thus can access the CCKBR biodistribution to positively impact pain circuitry neurons. Its high affinity binding permanently reverses chronic pain-, cognitive-, anxiety-, and depression-related behaviors.
A serious consequence of nerve injury pain or “neuropathic pain” is the transition to chronic pain that remains a significant clinical challenge with a treatment response rate of only 11% [15812][15913][159, 160]. While decades of study have been devoted to acute “nociceptive” mechanisms, it is clear that complex, multifactorial mechanisms are responsible for maintaining neuropathic pain long term, referred to as the “chronification” of pain. Current understanding is pain chronification causes physiological, molecular, epigenetic, and brain circuitry changes. While most studies are done in acute pain models, we utilize clinically relevant models of chronic neuropathic pain and find significantly reduced pain related behaviors after a single treatment with our scFv antibody targeting the P2X4 receptor (P2X4R) using ribosome display [16014][161]. P2X4R upregulation occurs in chronic pain, attributed to microglia in males and to T cells in females [16115][162]. These data provide support for pursuit of P2X4R scFvs as translational therapy for pain relief. We hope to develop non-opioid therapies to treat chronic pain using small protein, brain penetrant, scFv ingle chain Fragment variable (scFv) antibody therapies. These have the potential to reverse chronic neuropathic pain, associated pain-related behaviors and depression.
4. Future Perspectives
Ribosome display has proven to be a robust procedure, used now in academic and industrial laboratories, which comes rather close to experimental protein evolution in test tubes. Undoubtedly, the procedure will be further improved and applied to many new targets and selection goals. Together with the introduction of new technologies like next-generation sequencing, robotics, and nanotechnology, high-throughput screening of ribosome display libraries for rapid antibody generation is now a reality. Moreover, new molecular-based techniques for library generation and panning strategies will set the tone for the constant improvement of ribosome display in antibody generation for human infectious diseases, plant diseases, cancer, chronic pain and many more. Without a doubt, this technology will continue to evolve and play a bigger role in the coming decade within research, therapeutic, and diagnostic markets.
References
- Nina E. Weisser; J. Christopher Hall; Applications of single-chain variable fragment antibodies in therapeutics and diagnostics. BChiranjib Chakraborty; Manojit Bhattacharya; Ashish Ranjan Sharma; Present variants of concern and variants of interest of severe acute respiratory syndrome coronavirus 2: Their significant mutations in S‐glycoprotein, infectivity, re‐infectivity, immune escape and vaccines activity. Reviews iotn Mechndical Virology Advances 2009, 27, 502-520, 10.1016/j.biotechadv.2009.04.004.21, 0, e2270, 10.1002/rmv.2270.
- Marc A van Dijk; Jan G.J van de Winkel; Human antibodies as next generation therapeutics. Current Opinion in Chemical Biology 2001, 5, 368-374, 10.1016/s1367-5931(00)00216-7.Aleem, A.; AB, A.S.; Slenker A.K.. Emerging variants of SARS-CoV-2 and novel therapeutics against coronavirus (COVID-19); In: StatPearls, Eds.; StatPearls Publishing : Treasure Island (FL), 2022; pp. PMID: 34033342.
- Michael J Taussig; Oda Stoevesandt; Carl A K Borrebaeck; Andrew R Bradbury; Dolores Cahill; Christian Cambillau; Antoine De Daruvar; Stefan Dübel; Jutta Eichler; Ronald Frank; et al.Toby J GibsonDavid GloriamLarry GoldFriedrich W HerbergHenning HermjakobJörg D HoheiselThomas O JoosOlli KallioniemiManfred KoeglZoltán KonthurBernhard KornElisabeth KremmerSylvia KrobitschUlf LandegrenSilvère Van Der MaarelJohn McCaffertySerge MuyldermansPer-Åke NygrenSandrine PalcyAndreas PlückthunBojan PolicMichael PrzybylskiPetri SavirantaAlan SawyerDavid J ShermanArne SkerraMarkus TemplinMarius UeffingMathias Uhlén ProteomeBinders: planning a European resource of affinity reagents for analysis of the human proteome. William T. Harvey; Alessandro M. Carabelli; Ben Jackson; Ravindra K. Gupta; Emma C. Thomson; Ewan M. Harrison; Catherine Ludden; Richard Reeve; Andrew Rambaut; Sharon J. Peacock; et al.David L. Robertson SARS-CoV-2 variants, spike mutations and immune escape. Nature Reviews Methods icrobiology 2006, 4, 13-17, 10.1038/nmeth0107-13.21, 19, 409-424, 10.1038/s41579-021-00573-0.
- Zuhaida Asra Ahmad; Swee Keong Yeap; Abdul Manaf Ali; Wan Yong Ho; Noorjahan Banu Mohamed Alitheen; Muhajir Hamid; scFv Antibody: Principles and Clinical Application. ClinicKaiming Tao; Philip L. Tzou; Janin Nouhin; Ravindra K. Gupta; Tulio de Oliveira; Sergei L. Kosakovsky Pond; Daniela Fera; Robert W. Shafer; The biological and clinical significance of emerging SARS-CoV-2 variants. Nalture and Developmental Immunology Reviews Genetics 20212, , 2012, 1-15, 10.1155/2012/980250., 757-773, 10.1038/s41576-021-00408-x.
- Safar Farajnia; Vahideh Ahmadzadeh; Asghar Tanomand; Kamal Veisi; Shiva Ahdi Khosroshahi; Leila Rahbarnia; Development trends for generation of single-chain antibody fragments. ImmuChiranjib Chakraborty; Ashish Ranjan Sharma; Manojit Bhattacharya; Govindasamy Agoramoorthy; Sang-Soo Lee; The Drug Repurposing for COVID-19 Clinical Trials Provide Very Effective Therapeutic Combinations: Lessons Learned From Major Clinical Studies. Frontiers inop Pharmacology and Immunotoxicology 20214, 36, 297-308, 10.3109/08923973.2014.945126., 12, 704205, 10.3389/fphar.2021.704205.
- Winter, G.; Griffiths, A.D.; Hawkins, R.E.; Hoogenboom, H.R. Making antibodies by phage display technology. Annu. Rev. Immunol. 1994, 12, 433–455. [Google Scholar] [CrossRef]Mohammed Akli Ayoub; Pascale Crépieux; Markus Koglin; Marc Parmentier; Jean-Philippe Pin; Anne Poupon; Eric Reiter; Martine Smit; Jan Steyaert; Hervé Watier; et al.Trevor Wilkinson Antibodies targeting G protein-coupled receptors: Recent advances and therapeutic challenges. Platform development for expression and purification of stable isotope labeled monoclonal antibodies in Escherichia coli 2017, 9, 735-741, 10.1080/19420862.2017.1325052.
- George Georgiou; Christos Stathopoulos; Patrick S. Daugherty; Amiya R. Nayak; Brent L. Iverson; Roy Curtiss Iii; Display of heterologous proteins on the surface of microorganisms: From the screening of combinatorial libraries to live recombinant vaccines. NatuDavid C. Butler; Julie A. McLear; Anne Messer; Engineered antibody therapies to counteract mutant huntingtin and related toxic intracellular proteins. Progress Biotechnin Neurobiology 2011996, 15, 29-34, 10.1038/nbt0197-29., 97, 190-204, 10.1016/j.pneurobio.2011.11.004.
- Eric V Shusta; Jennifer Vanantwerp; K Dane Wittrup; Biosynthetic polypeptide libraries.. Current OpNives Škrlj; Marko Dolinar; New engineered antibodies against prions.. Binion in Biotechnology engineered 201999, 10, 117-122, 10.1016/s0958-1669(99)80020-2.3, 5, 10-4, 10.4161/bioe.26069.
- L. C. Mattheakis; R. R. Bhatt; W. J. Dower; An in vitro polysome display system for identifying ligands from very large peptide libraries.. PAlessandro Angelini; Yoshishige Miyabe; Daniel Newsted; Byron H. Kwan; Chie Miyabe; Ryan L. Kelly; Misha N. Jamy; Andrew D. Luster; K. Dane Wittrup; Directed evolution of broadly crossreactive chemokine-blocking antibodies efficacious in arthritis. Naturoceedings of the National Academy of Science Communications 201994, 8, 91, 9022-9026, 10.1073/pnas.91.19.9022., 1461, 10.1038/s41467-018-03687-x.
- Jozef Hanes; Andreas Plückthun; In vitro selection and evolution of functional proteins by using ribosome display. ProceedZuhaida Asra Ahmad; Swee Keong Yeap; Abdul Manaf Ali; Wan Yong Ho; Noorjahan Banu Mohamed Alitheen; Muhajir Hamid; scFv Antibody: Principles and Clinical Application. Clingsical of the National Academand Developmental Immunology of Sciences 201997, 94, 4937-4942, 10.1073/pnas.94.10.4937.2, 2012, 1-15, 10.1155/2012/980250.
- Mingyue He; M J Taussig; Antibody-ribosome-mRNA (ARM) complexes as efficient selection particles for in vitro display and evolution of antibody combining sites. K.N. Westlund; M.A. Montera; A.E. Goins; S.R.A. Alles; M. Afaghpour-Becklund; R. Bartel; R. Durvasula; A. Kunamneni; Single-chain Fragment variable antibody targeting cholecystokinin-B receptor for pain reduction. Neucleic Acids Research robiology of Pain 2021997, 25, 5132-5134, 10.1093/nar/25.24.5132., 10, 100067, 10.1016/j.ynpai.2021.100067.
- Richard W. Roberts; Jack W. Szostak; RNA-peptide fusions for the in vitro selection of peptides and proteins. PYaron Haviv; Yehuda Zadik; Yair Sharav; Rafael Benoliel; Painful Traumatic Trigeminal Neuropathy: An Open Study on the Pharmacotherapeutic Response to Stepped Treatment. Jouroceedings of the National Academy of Sciencal of Oral & Facial Pain and Headaches 201997, 94, 12297-12302, 10.1073/pnas.94.23.12297.3, 28, 52-60, 10.11607/jop.1154.
- Mingyue He; Bryan M. Edwards; Damjana Kastelic; Michael J. Taussig; Eukaryotic Ribosome Display with In Situ DNA Recovery. AdvancLene Baad-Hansen; Rafael Benoliel; Neuropathic orofacial pain: Facts and fiction. Ced Structurphal Safety Studies algia 2011, 805, 75-85, 10.1007/978-1-61779-379-0_5.7, 37, 670-679, 10.1177/0333102417706310.
- Mingyue He; Michael J Taussig; Eukaryotic ribosome display with in situ DNA recovery. NKarin N. Westlund; Marena A. Montera; Aleyah E. Goins; Sascha R. A. Alles; Nikita Suri; Sabrina L. McIlwrath; Robyn Bartel; Ravi V. Durvasula; Adinarayana Kunamneni; Single-Dose P2 X4R Single-Chain Fragment Variable Antibody Permanently Reverses Chronic Pain in Male Mice. International Joure Methodnal of Molecular Sciences 2007, 4, 281-288, 10.1038/nmeth1001.21, 22, 13612, 10.3390/ijms222413612.
- Mingyue He; Ribosome display: Cell-free protein display technology. BRobert E. Sorge; Josiane Mapplebeck; Sarah Rosen; Simon Beggs; Sarah Taves; Jessica K. Alexander; Loren Martin; Jean-Sebastien Austin; Susana G. Sotocinal; Di Chen; et al.Mu YangXiang Qun ShiHao HuangNicolas PillonPhilip J. BilanYu Shan TuAmira KlipRu-Rong JiJi ZhangMichael W. SalterJeffrey S. Mogil Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Naturiefings in Functional Genomics and Prot Neuroscienceomics 2001, 5, 1, 204-212, 10.1093/bfgp/1.2.204.8, 1081-1083, 10.1038/nn.4053.
- Mingyue He; Michael J. Taussig; Selection of Recombinant Antibodies by Eukaryotic Ribosome Display. Advanced Structural Safety Studies 2007, 484, 193-205, 10.1007/978-1-59745-398-1_13.He, M.; Taussig, M.J. Selection of recombinant antibodies by eukaryotic ribosome display. Methods Mol. Biol. 2008, 484, 193–205.
- Ryan L. Stafford; Marissa Matsumoto; Gang Yin; Q. Cai; Juan Jose Fung; Heather Stephenson; Avinash Gill; Monica You; Shwu-Hwa Lin; Willie D. Wang; et al.Mary Rose MasikatXiaofan LiKalyani PentaAlex R. SteinerRamesh BaligaChristopher J. MurrayChristopher D. ThanosTrevor J. HallamAaron K. Sato In vitro Fab display: a cell-free system for IgG discovery. Protein Engineering Design and Selection 2014, 27, 97-109, 10.1093/protein/gzu002.Stafford, R.; Matsumoto, M.; Yin, G.; Cai, Q.; Fung, J.J.; Stephenson, H.; Gill, A.; You, M.; Lin, S.-H.; Wang, W.; et al. In vitro Fab display: A cell-free system for IgG discovery. Protein Eng. Des. Sel. Peds 2014, 27.
- Larry C. Mattheakis; Jennifer M. Dias; William J. Dower; Cell-free synthesis of peptide libraries displayed on polysomes. Methods in Enzymology 1995, 267, 195-207, 10.1016/s0076-6879(96)67013-x.Mattheakis, L.C.; Dias, J.M.; Dower, W.J. Cell-free synthesis of peptide libraries displayed on polysomes. Methods Enzym. 1996, 267, 195–207.
- Gersuk, G.M.; Corey, M.J.; Corey, E.; Stray, J.E.; Kawasaki, G.H.; Vessella, R.L. High-affinity peptide ligands to prostate-specific antigen identified by polysome selection. Biochem. Biophys. Res. Commun. 1997, 232, 578–582.
- Hanes, J.; Jermutus, L.; Weber-Bornhauser, S.; Bosshard, H.R.; Pluckthun, A. Ribosome display efficiently selects and evolves high-affinity antibodies in vitro from immune libraries. Proc. Natl. Acad. Sci. USA 1998, 95, 14130–14135.
- Hanes, J.; Jermutus, L.; Schaffitzel, C.; Pluckthun, A. Comparison of Escherichia coli and rabbit reticulocyte ribosome display systems. Febs Lett. 1999, 450, 105–110.
- Lee, M.S.; Kwon, M.H.; Kim, K.H.; Shin, H.J.; Park, S.; Kim, H.I. Selection of scFvs specific for HBV DNA polymerase using ribosome display. J. Immunol. Methods 2004, 284, 147–157.
- Mingyue He; Michael J. Taussig; Ribosome display of antibodies: expression, specificity and recovery in a eukaryotic system. Journal of Immunological Methods 2005, 297, 73-82, 10.1016/j.jim.2004.11.022.He, M.; Taussig, M.J. Ribosome display of antibodies: Expression, specificity and recovery in a eukaryotic system. J. Immunol. Methods 2005, 297, 73–82.
- Kim, J.M.; Shin, H.J.; Kim, K.; Lee, M.S. A pseudoknot improves selection efficiency in ribosome display. Mol. Biotechnol. 2007, 36, 32–37.
- Qi, Y.; Wu, C.; Zhang, S.; Wang, Z.; Huang, S.; Dai, L.; Wang, S.; Xia, L.; Wen, K.; Cao, X.; et al. Selection of anti-sulfadimidine specific ScFvs from a hybridoma cell by eukaryotic ribosome display. PLoS ONE 2009, 4, e6427.
- Kastelic, D.; He, M. Ribosome display and screening for protein therapeutics. Methods Mol. Biol. 2012, 899, 61–72.
- Edwards, B.M.; He, M. Evolution of antibodies in vitro by ribosome display. Methods Mol. Biol. 2012, 907, 281–292.
- Douthwaite, J.A. Eukaryotic ribosome display selection using rabbit reticulocyte lysate. Methods Mol. Biol. 2012, 805, 45–57.
- Tang, J.; Wang, L.; Markiv, A.; Jeffs, S.A.; Dreja, H.; McKnight, A.; He, M.; Kang, A.S. Accessing of recombinant human monoclonal antibodies from patient libraries by eukaryotic ribosome display. Hum. Antibodies 2012, 21, 1–11.
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318.
- Bittner, M.; Kupferer, P.; Morris, C.F. Electrophoretic transfer of proteins and nucleic acids from slab gels to diazobenzyloxymethyl cellulose or nitrocellulose sheets. Anal. Biochem. 1980, 102, 459–471.
- Hawlisch, H.; Muller, M.; Frank, R.; Bautsch, W.; Klos, A.; Kohl, J. Site-specific anti-C3a receptor single-chain antibodies selected by differential panning on cellulose sheets. Anal. Biochem. 2001, 293, 142–145.
- Moghaddam, A.; Borgen, T.; Stacy, J.; Kausmally, L.; Simonsen, B.; Marvik, O.J.; Brekke, O.H.; Braunagel, M. Identification of scFv antibody fragments that specifically recognise the heroin metabolite 6-monoacetylmorphine but not morphine. J. Immunol. Methods 2003, 280, 139–155.
- Breitling, F.; Dubel, S.; Seehaus, T.; Klewinghaus, I.; Little, M. A surface expression vector for antibody screening. Gene 1991, 104, 147–153.
- Hust, M.; Maiss, E.; Jacobsen, H.J.; Reinard, T. The production of a genus-specific recombinant antibody (scFv) using a recombinant potyvirus protease. J. Virol. Methods 2002, 106, 225–233.
- Barbas, C.F., 3rd; Kang, A.S.; Lerner, R.A.; Benkovic, S.J. Assembly of combinatorial antibody libraries on phage surfaces: The gene III site. Proc. Natl. Acad. Sci. USA 1991, 88, 7978–7982.
- Sanna, P.P.; Williamson, R.A.; De Logu, A.; Bloom, F.E.; Burton, D.R. Directed selection of recombinant human monoclonal antibodies to herpes simplex virus glycoproteins from phage display libraries. Proc. Natl. Acad. Sci. USA 1995, 92, 6439–6443.
- Winter, G.; Milstein, C. Man-made antibodies. Nature 1991, 349, 293–299.
- Hoogenboom, H.R. Selecting and screening recombinant antibody libraries. Nat Biotechnol 2005, 23, 1105–1116.
- Hust, M.; Frenzel, A.; Schirrmann, T.; Dubel, S. Selection of recombinant antibodies from antibody gene libraries. Methods Mol Biol 2014, 1101, 305–320.
- Hust, M.; Dubel, S.; Schirrmann, T. Selection of recombinant antibodies from antibody gene libraries. Methods Mol. Biol. 2007, 408, 243–255.
- Hallborn, J.; Carlsson, R. Automated screening procedure for high-throughput generation of antibody fragments. Biotechniques 2002, 33 (Suppl. S6), 30–37.
- Konthur, Z.; Hust, M.; Dubel, S. Perspectives for systematic in vitro antibody generation. Gene 2005, 364, 19–29.
- Schirrmann, T.; Al-Halabi, L.; Dubel, S.; Hust, M. Production systems for recombinant antibodies. Front Biosci. 2008, 13, 4576–4594.
- Hust, M.; Meyer, T.; Voedisch, B.; Rulker, T.; Thie, H.; El-Ghezal, A.; Kirsch, M.I.; Schutte, M.; Helmsing, S.; Meier, D.; et al. A human scFv antibody generation pipeline for proteome research. J. Biotechnol. 2011, 152, 159–170.
- Groves, M.A.; Nickson, A.A. Affinity maturation of phage display antibody populations using ribosome display. Methods Mol. Biol. 2012, 805, 163–1690.
- Kobayashi, N.; Oyama, H.; Kato, Y.; Goto, J.; Soderlind, E.; Borrebaeck, C.A. Two-step in vitro antibody affinity maturation enables estradiol-17beta assays with more than 10-fold higher sensitivity. Anal. Chem. 2010, 82, 1027–1038.
- Finlay, W.J.; Cunningham, O.; Lambert, M.A.; Darmanin-Sheehan, A.; Liu, X.; Fennell, B.J.; Mahon, C.M.; Cummins, E.; Wade, J.M.; O’Sullivan, C.M.; et al. Affinity maturation of a humanized rat antibody for anti-RAGE therapy: Comprehensive mutagenesis reveals a high level of mutational plasticity both inside and outside the complementarity-determining regions. J. Mol. Biol. 2009, 388, 541–558.
- Lipovsek, D.; Pluckthun, A. In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 2004, 290, 51–67.
- Hanes, J.; Schaffitzel, C.; Knappik, A.; Pluckthun, A. Picomolar affinity antibodies from a fully synthetic naive library selected and evolved by ribosome display. Nat. Biotechnol. 2000, 18, 1287–1292.
- Schaffitzel, C.; Berger, I.; Postberg, J.; Hanes, J.; Lipps, H.J.; Pluckthun, A. In vitro generated antibodies specific for telomeric guanine-quadruplex DNA react with Stylonychia lemnae macronuclei. Proc. Natl. Acad. Sci. USA 2001, 98, 8572–8577.
- Jermutus, L.; Honegger, A.; Schwesinger, F.; Hanes, J.; Pluckthun, A. Tailoring in vitro evolution for protein affinity or stability. Proc. Natl. Acad. Sci. USA 2001, 98, 75–80.
- Levin, A.M.; Weiss, G.A. Optimizing the affinity and specificity of proteins with molecular display. Mol. Biosyst. 2006, 2, 49–57.
- Dreier, B.; Pluckthun, A. Rapid selection of high-affinity binders using ribosome display. Methods Mol. Biol. 2012, 805, 261–286.
- Lewis, L.; Lloyd, C. Optimisation of antibody affinity by ribosome display using error-prone or site-directed mutagenesis. Methods Mol. Biol. 2012, 805, 139–161.
- Zahnd, C.; Spinelli, S.; Luginbuhl, B.; Amstutz, P.; Cambillau, C.; Pluckthun, A. Directed in vitro evolution and crystallographic analysis of a peptide-binding single chain antibody fragment (scFv) with low picomolar affinity. J. Biol. Chem. 2004, 279, 18870–18877.
- Chin, S.E.; Ferraro, F.; Groves, M.; Liang, M.; Vaughan, T.J.; Dobson, C.L. Isolation of high affinity, neutralizing anti-idiotype antibodies by phage and ribosome display for application in immunogenicity and pharmacokinetic analyses. J. Immunol. Methods 2015, 416, 49–58.
- Groves, M.A.; Amanuel, L.; Campbell, J.I.; Rees, D.G.; Sridharan, S.; Finch, D.K.; Lowe, D.C.; Vaughan, T.J. Antibody VH and VL recombination using phage and ribosome display technologies reveals distinct structural routes to affinity improvements with VH-VL interface residues providing important structural diversity. MAbs 2014, 6, 236–245.
- Hu, D.; Tateno, H.; Hirabayashi, J. Directed evolution of lectins by an improved error-prone PCR and ribosome display method. Methods Mol. Biol. 2014, 1200, 527–538.
- Kanamori, T.; Fujino, Y.; Ueda, T. PURE ribosome display and its application in antibody technology. Biochim. Biophys. Acta 2014, 1844, 1925–1932.
- Darmanin-Sheehan, A.; Finlay, W.J.; Cunningham, O.; Fennell, B.J. Molecular scanning: Combining random mutagenesis, ribosome display, and bioinformatic analysis for protein engineering. Methods Mol. Biol. 2012, 907, 487–503.
- Lei, L. Identification of candidate vaccine genes using ribosome display. Methods Mol. Biol. 2012, 805, 299–314.
- Groves, M.A.; Osbourn, J.K. Applications of ribosome display to antibody drug discovery. Expert Opin. Biol. 2005, 5, 125–135.
- Chodorge, M.; Fourage, L.; Ravot, G.; Jermutus, L.; Minter, R. In vitro DNA recombination by L-Shuffling during ribosome display affinity maturation of an anti-Fas antibody increases the population of improved variants. Protein Eng. Des. Sel. Peds 2008, 21, 343–351.
- Heyduk, E.; Heyduk, T. Ribosome display enhanced by next generation sequencing: A tool to identify antibody-specific peptide ligands. Anal. Biochem. 2014, 464, 73–82.
- Li, R.; Kang, G.; Hu, M.; Huang, H. Ribosome Display: A Potent Display Technology used for Selecting and Evolving Specific Binders with Desired Properties. Mol. Biotechnol. 2019, 61, 60–71.
- Ryabova, L.A.; Desplancq, D.; Spirin, A.S.; Plückthun, A. Functional antibody production using cell-free translation: Effects of protein disulfide isomerase and chaperones. Nat. Biotechnol. 1997, 15, 79–84.
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554.
- Glockshuber, R.; Malia, M.; Pfitzinger, I.; Plueckthun, A. A comparison of strategies to stabilize immunoglobulin Fv-fragments. Biochemistry 1990, 29, 1362–1367.
- Groves, M.; Lane, S.; Douthwaite, J.; Lowne, D.; Rees, D.G.; Edwards, B.; Jackson, R.H. Affinity maturation of phage display antibody populations using ribosome display. J. Immunol. Methods 2006, 313, 129–139.
- Sun, Y.; Ning, B.; Liu, M.; Gao, X.; Fan, X.; Liu, J.; Gao, Z. Selection of diethylstilbestrol-specific single-chain antibodies from a non-immunized mouse ribosome display library. PLoS ONE 2012, 7, e33186.
- Whiteaker, J.R.; Zhao, L.; Frisch, C.; Ylera, F.; Harth, S.; Knappik, A.; Paulovich, A.G. High-affinity recombinant antibody fragments (Fabs) can be applied in peptide enrichment immuno-MRM assays. J. Proteome Res. 2014, 13, 2187–2196.
- Ylera, F.; Harth, S.; Waldherr, D.; Frisch, C.; Knappik, A. Off-rate screening for selection of high-affinity anti-drug antibodies. Anal. Biochem. 2013, 441, 208–213.
- Binz, H.K.; Stumpp, M.T.; Forrer, P.; Amstutz, P.; Pluckthun, A. Designing repeat proteins: Well-expressed, soluble and stable proteins from combinatorial libraries of consensus ankyrin repeat proteins. J. Mol. Biol. 2003, 332, 489–503.
- Binz, H.K.; Amstutz, P.; Kohl, A.; Stumpp, M.T.; Briand, C.; Forrer, P.; Grutter, M.G.; Pluckthun, A. High-affinity binders selected from designed ankyrin repeat protein libraries. Nat. Biotechnol. 2004, 22, 575–582.
- Dreier, B.; Pluckthun, A. Ribosome display: A technology for selecting and evolving proteins from large libraries. Methods Mol. Biol. 2011, 687, 283–306.
- Schilling, J.; Schoppe, J.; Pluckthun, A. From DARPins to LoopDARPins: Novel LoopDARPin design allows the selection of low picomolar binders in a single round of ribosome display. J. Mol. Biol. 2014, 426, 691–721.
- Pluckthun, A. Ribosome display: A perspective. Methods Mol. Biol. 2012, 805, 3–28.
- Stefan, N.; Martin-Killias, P.; Wyss-Stoeckle, S.; Honegger, A.; Zangemeister-Wittke, U.; Pluckthun, A. DARPins recognizing the tumor-associated antigen EpCAM selected by phage and ribosome display and engineered for multivalency. J. Mol. Biol. 2011, 413, 826–843.
- Schilling, J.; Schoppe, J.; Sauer, E.; Pluckthun, A. Co-crystallization with conformation-specific designed ankyrin repeat proteins explains the conformational flexibility of BCL-W. J. Mol. Biol. 2014, 426, 2346–2362.
- Scholz, O.; Hansen, S.; Pluckthun, A. G-quadruplexes are specifically recognized and distinguished by selected designed ankyrin repeat proteins. Nucleic Acids Res. 2014, 42, 9182–9194.
- Brauchle, M.; Hansen, S.; Caussinus, E.; Lenard, A.; Ochoa-Espinosa, A.; Scholz, O.; Sprecher, S.G.; Pluckthun, A.; Affolter, M. Protein interference applications in cellular and developmental biology using DARPins that recognize GFP and mCherry. Biol. Open 2014, 3, 1252–1261.
- Tamaskovic, R.; Simon, M.; Stefan, N.; Schwill, M.; Pluckthun, A. Designed ankyrin repeat proteins (DARPins) from research to therapy. Methods Enzym. 2012, 503, 101–134.
- Wetzel, S.K.; Ewald, C.; Settanni, G.; Jurt, S.; Pluckthun, A.; Zerbe, O. Residue-resolved stability of full-consensus ankyrin repeat proteins probed by NMR. J. Mol. Biol. 2010, 402, 241–258.
- Wetzel, S.K.; Settanni, G.; Kenig, M.; Binz, H.K.; Pluckthun, A. Folding and unfolding mechanism of highly stable full-consensus ankyrin repeat proteins. J. Mol. Biol. 2008, 376, 241–257.
- Schweizer, A.; Roschitzki-Voser, H.; Amstutz, P.; Briand, C.; Gulotti-Georgieva, M.; Prenosil, E.; Binz, H.K.; Capitani, G.; Baici, A.; Pluckthun, A.; et al. Inhibition of caspase-2 by a designed ankyrin repeat protein: Specificity, structure, and inhibition mechanism. Structure 2007, 15, 625–636.
- Zahnd, C.; Wyler, E.; Schwenk, J.M.; Steiner, D.; Lawrence, M.C.; McKern, N.M.; Pecorari, F.; Ward, C.W.; Joos, T.O.; Pluckthun, A. A designed ankyrin repeat protein evolved to picomolar affinity to Her2. J. Mol. Biol. 2007, 369, 1015–1028.
- Zahnd, C.; Pecorari, F.; Straumann, N.; Wyler, E.; Pluckthun, A. Selection and characterization of Her2 binding-designed ankyrin repeat proteins. J. Biol. Chem. 2006, 281, 35167–35175.
- Amstutz, P.; Koch, H.; Binz, H.K.; Deuber, S.A.; Pluckthun, A. Rapid selection of specific MAP kinase-binders from designed ankyrin repeat protein libraries. Protein Eng. Des. Sel. Peds 2006, 19, 219–229.
- Amstutz, P.; Binz, H.K.; Parizek, P.; Stumpp, M.T.; Kohl, A.; Grutter, M.G.; Forrer, P.; Pluckthun, A. Intracellular kinase inhibitors selected from combinatorial libraries of designed ankyrin repeat proteins. J. Biol. Chem. 2005, 280, 24715–24722.
- Dreier, B.; Mikheeva, G.; Belousova, N.; Parizek, P.; Boczek, E.; Jelesarov, I.; Forrer, P.; Pluckthun, A.; Krasnykh, V. Her2-specific multivalent adapters confer designed tropism to adenovirus for gene targeting. J. Mol. Biol. 2011, 405, 410–426.
- Veesler, D.; Dreier, B.; Blangy, S.; Lichiere, J.; Tremblay, D.; Moineau, S.; Spinelli, S.; Tegoni, M.; Pluckthun, A.; Campanacci, V.; et al. Crystal structure and function of a DARPin neutralizing inhibitor of lactococcal phage TP901–1: Comparison of DARPin and camelid VHH binding mode. J. Bio.l Chem. 2009, 284, 30718–30726.
- Milovnik, P.; Ferrari, D.; Sarkar, C.A.; Pluckthun, A. Selection and characterization of DARPins specific for the neurotensin receptor 1. Protein Eng. Des. Sel. Peds 2009, 22, 357–366.
- Scholz, O.; Henssler, E.M.; Bail, J.; Schubert, P.; Bogdanska-Urbaniak, J.; Sopp, S.; Reich, M.; Wisshak, S.; Kostner, M.; Bertram, R.; et al. Activity reversal of Tet repressor caused by single amino acid exchanges. Mol. Microbiol. 2004, 53, 777–789.
- Yau, K.Y.; Dubuc, G.; Li, S.; Hirama, T.; Mackenzie, C.R.; Jermutus, L.; Hall, J.C.; Tanha, J. Affinity maturation of a V(H)H by mutational hotspot randomization. J. Immunol. Methods 2005, 297, 213–224.
- Perruchini, C.; Pecorari, F.; Bourgeois, J.P.; Duyckaerts, C.; Rougeon, F.; Lafaye, P. Llama VHH antibody fragments against GFAP: Better diffusion in fixed tissues than classical monoclonal antibodies. Acta Neuropathol. 2009, 118, 685–695.
- WHO. Ebola Virus Disease (Fact Sheets). Archived 30th May 2019. Available online: https://www.who.int/en/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 4 February 2020).
- Kunamneni, A.; Clarke, E.C.; Ye, C.; Bradfute, S.B.; Durvasula, R. Generation and Selection of a Panel of Pan-Filovirus Single-Chain Antibodies using Cell-Free Ribosome Display. Am. J. Trop. Med. Hyg. 2019, 101, 198–206.
- Robert W. Malone; Jane Homan; Michael V. Callahan; Jill Glasspool Malone; Lambodhar Damodaran; Adriano de Bernardi Schneider; Rebecca Zimler; James Talton; Ronald R. Cobb; Ivan Ruzic; et al.Julie Smith-GagenDaniel JaniesJames WilsonZika Response Working Group Zika Virus: Medical Countermeasure Development Challenges. PLOS Neglected Tropical Diseases 2016, 10, e0004530, 10.1371/journal.pntd.0004530.Malone, R.W.; Homan, J.; Callahan, M.V.; Glasspool-Malone, J.; Damodaran, L.; Schneider Ade, B.; Zimler, R.; Talton, J.; Cobb, R.R.; Ruzic, I.; et al. Zika Virus: Medical Countermeasure Development Challenges. Plos Negl. Trop. Dis. 2016, 10, e0004530.
- WHO. Zika Virus (Fact Sheets). Archived 20th July 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/zika-virus (accessed on 4 February 2020).
- Adinarayana Kunamneni; Chunyan Ye; Steven B. Bradfute; Ravi Durvasula; Ribosome display for the rapid generation of high-affinity Zika-neutralizing single-chain antibodies. PLOS ONE 2018, 13, e0205743, 10.1371/journal.pone.0205743.Kunamneni, A.; Ye, C.; Bradfute, S.B.; Durvasula, R. Ribosome display for the rapid generation of high-affinity Zika-neutralizing single-chain antibodies. PLoS ONE 2018, 13, e0205743.
- WHO. Tuberculosis. Archived 24th March, 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 12 May 2020).
- Haileyesus Getahun; Mark Harrington; Rick O'Brien; Paul Nunn; Diagnosis of smear-negative pulmonary tuberculosis in people with HIV infection or AIDS in resource-constrained settings: informing urgent policy changes. The Lancet 2007, 369, 2042-2049, 10.1016/s0140-6736(07)60284-0.Getahun, H.; Harrington, M.; O’Brien, R.; Nunn, P. Diagnosis of smear-negative pulmonary tuberculosis in people with HIV infection or AIDS in resource-constrained settings: Informing urgent policy changes. Lancet 2007, 369, 2042–2049.
- Shahrzad Ahangarzadeh; Mojgan Bandehpour; Bahram Kazemi; Selection of single-chain variable fragments specific for Mycobacterium tuberculosis ESAT-6 antigen using ribosome display. Iran J Basic Med Sci 2017, 20, 327-333, 10.22038/IJBMS.2017.8363.Ahangarzadeh, S.; Bandehpour, M.; Kazemi, B. Selection of single-chain variable fragments specific for Mycobacterium tuberculosis ESAT-6 antigen using ribosome display. Iran J. Basic Med. Sci. 2017, 20, 327–333.
- WHO. Q&A on Coronaviruses (COVID-19). Archived 17th April, 2020. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/question-and-answers-hub/q-a-detail/q-a-coronaviruses (accessed on 12 May 2020).
- (WHO), W.H.O. Coronavirus disease (COVID-19) Pandemic. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 12 May 2020).
- Chiranjib Chakraborty; Manojit Bhattacharya; Ashish Ranjan Sharma; Present variants of concern and variants of interest of severe acute respiratory syndrome coronavirus 2: Their significant mutations in S‐glycoprotein, infectivity, re‐infectivity, immune escape and vaccines activity. Reviews in Medical Virology 2021, 0, e2270, 10.1002/rmv.2270.Schaffitzel, C.; Hanes, J.; Jermutus, L.; Pluckthun, A. Ribosome display: An in vitro method for selection and evolution of antibodies from libraries. J. Immunol. Methods 1999, 231, 119–135.
- Aleem, A.; AB, A.S.; Slenker A.K.. Emerging variants of SARS-CoV-2 and novel therapeutics against coronavirus (COVID-19); In: StatPearls, Eds.; StatPearls Publishing : Treasure Island (FL), 2022; pp. PMID: 34033342.Huang, S.; Feng, L.; An, G.; Zhang, X.; Zhao, Z.; Han, R.; Lei, F.; Zhang, Y.; Luo, A.; Jing, X.; et al. Ribosome display and selection of single-chain variable fragments effectively inhibit growth and progression of microspheres in vitro and in vivo. Cancer Sci. 2018, 109, 1503–1512.
- William T. Harvey; Alessandro M. Carabelli; Ben Jackson; Ravindra K. Gupta; Emma C. Thomson; Ewan M. Harrison; Catherine Ludden; Richard Reeve; Andrew Rambaut; Sharon J. Peacock; et al.David L. Robertson SARS-CoV-2 variants, spike mutations and immune escape. Nature Reviews Microbiology 2021, 19, 409-424, 10.1038/s41579-021-00573-0.Center for Disease Control and Prevention (CDC), About HIV/AIDS. Archived 2nd December, 2019. Available online: https://www.cdc.gov/hiv/basics/whatishiv.html (accessed on 17 May 2020).
- Kaiming Tao; Philip L. Tzou; Janin Nouhin; Ravindra K. Gupta; Tulio de Oliveira; Sergei L. Kosakovsky Pond; Daniela Fera; Robert W. Shafer; The biological and clinical significance of emerging SARS-CoV-2 variants. Nature Reviews Genetics 2021, 22, 757-773, 10.1038/s41576-021-00408-x.WHO. HIV/AIDS. Archived 17th April, 2020. Available online: https://www.who.int/health-topics/hiv-aids/#tab=tab_1 (accessed on 17 May 2020).
- Chiranjib Chakraborty; Ashish Ranjan Sharma; Manojit Bhattacharya; Govindasamy Agoramoorthy; Sang-Soo Lee; The Drug Repurposing for COVID-19 Clinical Trials Provide Very Effective Therapeutic Combinations: Lessons Learned From Major Clinical Studies. Frontiers in Pharmacology 2021, 12, 704205, 10.3389/fphar.2021.704205.Burton, D.R.; Weiss, R.A. AIDS/HIV. A boost for HIV vaccine design. Science 2010, 329, 770–773.
- Kunamneni, A.; Ye, C.; Bradfute, S.B.; Durvasula, R. Ribosome display for the rapid generation of high-affinity Zika-neutralizing single-chain antibodies. PLoS ONE 2018, 13, e0205743. Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.; Pietzsch, J.; Fenyö, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637.
- Shangke Huang; Lu Feng; Gaili An; Xiaojin Zhang; Zixuan Zhao; Rui Han; Fuxi Lei; Yujiao Zhang; Anqi Luo; Xin Jing; et al.Lin ZhaoShanzhi GuXinhan ZhaoLingxiao Zhang Ribosome display and selection of single‐chain variable fragments effectively inhibit growth and progression of microspheres in vitro and in vivo. Cancer Science 2018, 109, 1503-1512, 10.1111/cas.13574.Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470.
- Getahun, H.; Harrington, M.; O’Brien, R.; Nunn, P. Diagnosis of smear-negative pulmonary tuberculosis in people with HIV infection or AIDS in resource-constrained settings: Informing urgent policy changes. Lancet 2007, 369, 2042–2049. Davis, M.J.; Purcell, A.H.; Thomson, S.V. Pierce’s disease of grapevines: Isolation of the causal bacterium. Science 1978, 199, 75–77.
- Ahangarzadeh, S.; Bandehpour, M.; Kazemi, B. Selection of single-chain variable fragments specific for Mycobacterium tuberculosis ESAT-6 antigen using ribosome display. Iran J. Basic Med. Sci. 2017, 20, 327–333. Morano, L.D.; Bextine, B.R.; Garcia, D.A.; Maddox, S.V.; Gunawan, S.; Vitovsky, N.J.; Black, M.C. Initial genetic analysis of Xylella fastidiosa in Texas. Curr. Microbiol. 2008, 56, 346–351.
- Dennis R. Burton; Robin A. Weiss; A Boost for HIV Vaccine Design. Science 2010, 329, 770-773, 10.1126/science.1194693.Ramirez, J.L.; Lacava, P.T.; Miller, T.A. Detection of the bacterium, Xylella fastidiosa, in saliva of glassy-winged sharpshooter, Homalodisca vitripennis. J. Insect Sci. 2008, 8, 1–7.
- Johannes F. Scheid; Hugo Mouquet; Beatrix Ueberheide; Ron Diskin; Florian Klein; Thiago Y. K. Oliveira; John Pietzsch; David Fenyo; Alexander Abadir; Klara Velinzon; et al.Arlene HurleySunnie MyungFarid BouladPascal PoignardDennis R. BurtonFlorencia PereyraDavid D. HoBruce D. WalkerMichael S. SeamanPamela J. BjorkmanBrian T. ChaitMichel C. Nussenzweig Sequence and Structural Convergence of Broad and Potent HIV Antibodies That Mimic CD4 Binding. Science 2011, 333, 1633-1637, 10.1126/science.1207227.Myers, A.L.; Sutton, T.B.; Abad, J.A.; Kennedy, G.G. Pierce’s Disease of Grapevines: Identification of the Primary Vectors in North Carolina. Phytopathology 2007, 97, 1440–1450.
- Laura M. Walker; Michael Huber; Katie Doores; Emilia Falkowska; Robert Pejchal; Jean-Philippe Julien; Sheng-Kai Wang; Alejandra Ramos; Po-Ying Chan-Hui; Matthew Moyle; et al.Jennifer L. MitchamPhillip W. HammondOle A. OlsenPham PhungSteven FlingChi-Huey WongSanjay PhogatTerri WrinMelissa D. SimekProtocol G. Principal InvestigatorsWayne C. KoffIan A. WilsonDennis R. BurtonPascal Poignard Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466-470, 10.1038/nature10373.Jackson, B.C.; Blua, M.J.; Bextine, B. Impact of duration versus frequency of probing by Homalodisca vitripennis (Hemiptera: Cicadellidae) on inoculation of Xylella fastidiosa. J. Econ. Entomol. 2008, 101, 1122–1126.
- Huang, S.; Feng, L.; An, G.; Zhang, X.; Zhao, Z.; Han, R.; Lei, F.; Zhang, Y.; Luo, A.; Jing, X.; et al. Ribosome display and selection of single-chain variable fragments effectively inhibit growth and progression of microspheres in vitro and in vivo. Cancer Sci. 2018, 109, 1503–1512. Hendson, M.; Purcell, A.H.; Chen, D.; Smart, C.; Guilhabert, M.; Kirkpatrick, B. Genetic diversity of Pierce’s disease strains and other pathotypes of Xylella fastidiosa. Appl. Environ. Microbiol. 2001, 67, 895–903.
- Center for Disease Control and Prevention (CDC), About HIV/AIDS. Archived 2nd December, 2019. Available online: https://www.cdc.gov/hiv/basics/whatishiv.html (accessed on 17 May 2020).Simpson, A.J.; Reinach, F.C.; Arruda, P.; Abreu, F.A.; Acencio, M.; Alvarenga, R.; Alves, L.M.; Araya, J.E.; Baia, G.S.; Baptista, C.S.; et al. The genome sequence of the plant pathogen Xylella fastidiosa. The Xylella fastidiosa Consortium of the Organization for Nucleotide Sequencing and Analysis. Nature 2000, 406, 151–159.
- WHO. HIV/AIDS. Archived 17th April, 2020. Available online: https://www.who.int/health-topics/hiv-aids/#tab=tab_1 (accessed on 17 May 2020).Retchless, A.; Labroussaa, F.; Shapiro, L.; Stenger, D.; Lindow, S.; Almeida, R. Genomic Insights into Xylella Fastidiosa Interactions with Plant and Insect Hosts; Springer: Berlin, Germany, 2014; pp. 177–202.
- Burton, D.R.; Weiss, R.A. AIDS/HIV. A boost for HIV vaccine design. Science 2010, 329, 770–773. Caserta, R.; Takita, M.A.; Targon, M.L.; Rosselli-Murai, L.K.; de Souza, A.P.; Peroni, L.; Stach-Machado, D.R.; Andrade, A.; Labate, C.A.; Kitajima, E.W.; et al. Expression of Xylella fastidiosa fimbrial and afimbrial proteins during biofilm formation. Appl. Environ. Microbiol. 2010, 76, 4250–4259.
- Scheid, J.F.; Mouquet, H.; Ueberheide, B.; Diskin, R.; Klein, F.; Oliveira, T.; Pietzsch, J.; Fenyö, D.; Abadir, A.; Velinzon, K.; et al. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 2011, 333, 1633–1637. Fjellbirkeland, A.; Bemanian, V.; McDonald, I.R.; Murrell, J.C.; Jensen, H.B. Molecular analysis of an outer membrane protein, MopB, of Methylococcus capsulatus (Bath) and structural comparisons with proteins of the OmpA family. Arch. Microbiol. 2000, 173, 346–351.
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. Voegel, T.M.; Warren, J.G.; Matsumoto, A.; Igo, M.M.; Kirkpatrick, B.C. Localization and characterization of Xylella fastidiosa haemagglutinin adhesins. Microbiology 2010, 156 Pt 7, 2172–2179.
- Davis, M.J.; Purcell, A.H.; Thomson, S.V. Pierce’s disease of grapevines: Isolation of the causal bacterium. Science 1978, 199, 75–77. Pierce, B.K.; Voegel, T.; Kirkpatrick, B.C. The Xylella fastidiosa PD1063 protein is secreted in association with outer membrane vesicles. PLoS ONE 2014, 9, e113504.
- Morano, L.D.; Bextine, B.R.; Garcia, D.A.; Maddox, S.V.; Gunawan, S.; Vitovsky, N.J.; Black, M.C. Initial genetic analysis of Xylella fastidiosa in Texas. Curr. Microbiol. 2008, 56, 346–351. Bishop, R.E. Structural biology of membrane-intrinsic beta-barrel enzymes: Sentinels of the bacterial outer membrane. Biochim. Et Biophys. Acta 2008, 1778, 1881–1896.
- Ramirez, J.L.; Lacava, P.T.; Miller, T.A. Detection of the bacterium, Xylella fastidiosa, in saliva of glassy-winged sharpshooter, Homalodisca vitripennis. J. Insect Sci. 2008, 8, 1–7. Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253.
- Myers, A.L.; Sutton, T.B.; Abad, J.A.; Kennedy, G.G. Pierce’s Disease of Grapevines: Identification of the Primary Vectors in North Carolina. Phytopathology 2007, 97, 1440–1450. Kostakioti, M.; Newman, C.L.; Thanassi, D.G.; Stathopoulos, C. Mechanisms of protein export across the bacterial outer membrane. J. Bacteriol. 2005, 187, 4306–4314.
- Jackson, B.C.; Blua, M.J.; Bextine, B. Impact of duration versus frequency of probing by Homalodisca vitripennis (Hemiptera: Cicadellidae) on inoculation of Xylella fastidiosa. J. Econ. Entomol. 2008, 101, 1122–1126. Chen, Y.Y.; Wu, C.H.; Lin, J.W.; Weng, S.F.; Tseng, Y.H. Mutation of the gene encoding a major outer-membrane protein in Xanthomonas campestris pv. campestris causes pleiotropic effects, including loss of pathogenicity. Microbiology 2010, 156 Pt 9, 2842–2854.
- Hendson, M.; Purcell, A.H.; Chen, D.; Smart, C.; Guilhabert, M.; Kirkpatrick, B. Genetic diversity of Pierce’s disease strains and other pathotypes of Xylella fastidiosa. Appl. Environ. Microbiol. 2001, 67, 895–903. Gotoh, N.; Wakebe, H.; Yoshihara, E.; Nakae, T.; Nishino, T. Role of protein F in maintaining structural integrity of the Pseudomonas aeruginosa outer membrane. J. Bacteriol. 1989, 171, 983–990.
- Simpson, A.J.; Reinach, F.C.; Arruda, P.; Abreu, F.A.; Acencio, M.; Alvarenga, R.; Alves, L.M.; Araya, J.E.; Baia, G.S.; Baptista, C.S.; et al. The genome sequence of the plant pathogen Xylella fastidiosa. The Xylella fastidiosa Consortium of the Organization for Nucleotide Sequencing and Analysis. Nature 2000, 406, 151–159. Woodruff, W.A.; Hancock, R.E. Pseudomonas aeruginosa outer membrane protein F: Structural role and relationship to the Escherichia coli OmpA protein. J. Bacteriol. 1989, 171, 3304–3309.
- Retchless, A.; Labroussaa, F.; Shapiro, L.; Stenger, D.; Lindow, S.; Almeida, R. Genomic Insights into Xylella Fastidiosa Interactions with Plant and Insect Hosts; Springer: Berlin, Germany, 2014; pp. 177–202. Wang, Y. The function of OmpA in Escherichia coli. Biochem. Biophys. Res. Commun. 2002, 292, 396–401.
- Caserta, R.; Takita, M.A.; Targon, M.L.; Rosselli-Murai, L.K.; de Souza, A.P.; Peroni, L.; Stach-Machado, D.R.; Andrade, A.; Labate, C.A.; Kitajima, E.W.; et al. Expression of Xylella fastidiosa fimbrial and afimbrial proteins during biofilm formation. Appl. Environ. Microbiol. 2010, 76, 4250–4259. Khan, N.A.; Shin, S.; Chung, J.W.; Kim, K.J.; Elliott, S.; Wang, Y.; Kim, K.S. Outer membrane protein A and cytotoxic necrotizing factor-1 use diverse signaling mechanisms for Escherichia coli K1 invasion of human brain microvascular endothelial cells. Microb. Pathog. 2003, 35, 35–42.
- Fjellbirkeland, A.; Bemanian, V.; McDonald, I.R.; Murrell, J.C.; Jensen, H.B. Molecular analysis of an outer membrane protein, MopB, of Methylococcus capsulatus (Bath) and structural comparisons with proteins of the OmpA family. Arch. Microbiol. 2000, 173, 346–351. Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.H.; Kim, K.S. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 1996, 64, 146–153.
- Voegel, T.M.; Warren, J.G.; Matsumoto, A.; Igo, M.M.; Kirkpatrick, B.C. Localization and characterization of Xylella fastidiosa haemagglutinin adhesins. Microbiology 2010, 156 Pt 7, 2172–2179. Killiny, N.; Rashed, A.; Almeida, R.P. Disrupting the transmission of a vector-borne plant pathogen. Appl. Environ. Microbiol. 2012, 78, 638–643.
- Pierce, B.K.; Voegel, T.; Kirkpatrick, B.C. The Xylella fastidiosa PD1063 protein is secreted in association with outer membrane vesicles. PLoS ONE 2014, 9, e113504. Lampe, D.J.; Lauzon, C.R.; Miller, T. Development of symbiotic control of Pierce’s Disease. 2016. Available online: https://biopesticide.ucr.edu/abstracts/assets/Lampe_abstract.pdf (accessed on 12 April 2020).
- Bishop, R.E. Structural biology of membrane-intrinsic beta-barrel enzymes: Sentinels of the bacterial outer membrane. Biochim. Et Biophys. Acta 2008, 1778, 1881–1896. Miller, T. Symbiotic Control in agriculture and medicine. Comp. Biochem. Physiol. A Mol. Integr. Physiol. Comp. Biochem. Physiol. Pt A 2007, 146.
- Koebnik, R.; Locher, K.P.; Van Gelder, P. Structure and function of bacterial outer membrane proteins: Barrels in a nutshell. Mol. Microbiol. 2000, 37, 239–253. Azizi, A.; Arora, A.; Markiv, A.; Lampe, D.J.; Miller, T.A.; Kang, A.S. Ribosome display of combinatorial antibody libraries derived from mice immunized with heat-killed Xylella fastidiosa and the selection of MopB-specific single-chain antibodies. Appl. Environ. Microbiol. 2012, 78, 2638–2647.
- Kostakioti, M.; Newman, C.L.; Thanassi, D.G.; Stathopoulos, C. Mechanisms of protein export across the bacterial outer membrane. J. Bacteriol. 2005, 187, 4306–4314. Feil, H.; Feil, W.S.; Lindow, S.E. Contribution of Fimbrial and Afimbrial Adhesins of Xylella fastidiosa to Attachment to Surfaces and Virulence to Grape. Phytopathology 2007, 97, 318–324.
- Chen, Y.Y.; Wu, C.H.; Lin, J.W.; Weng, S.F.; Tseng, Y.H. Mutation of the gene encoding a major outer-membrane protein in Xanthomonas campestris pv. campestris causes pleiotropic effects, including loss of pathogenicity. Microbiology 2010, 156 Pt 9, 2842–2854. De La Fuente, L.; Burr, T.J.; Hoch, H.C. Autoaggregation of Xylella fastidiosa cells is influenced by type I and type IV pili. Appl. Environ. Microbiol. 2008, 74, 5579–5582.
- Gotoh, N.; Wakebe, H.; Yoshihara, E.; Nakae, T.; Nishino, T. Role of protein F in maintaining structural integrity of the Pseudomonas aeruginosa outer membrane. J. Bacteriol. 1989, 171, 983–990. Hoch, H.C. Continued assessment of Xylella fastidiosa fimbrial adhesins as important virulence factors in Pierce’s disease: Influence of xylem sap, p 87–91. In Proceedings of the 2010 Pierce’s Disease Research Symposium; Esser, T., Ed.; California Department of Food and Agriculture: Sacramento, CA, USA, 2010.
- Woodruff, W.A.; Hancock, R.E. Pseudomonas aeruginosa outer membrane protein F: Structural role and relationship to the Escherichia coli OmpA protein. J. Bacteriol. 1989, 171, 3304–3309. Li, Y.; Hao, G.; Galvani, C.D.; Meng, Y.; De La Fuente, L.; Hoch, H.C.; Burr, T.J. Type I and type IV pili of Xylella fastidiosa affect twitching motility, biofilm formation and cell-cell aggregation. Microbiology 2007, 153 Pt 3, 719–726.
- Wang, Y. The function of OmpA in Escherichia coli. Biochem. Biophys. Res. Commun. 2002, 292, 396–401. Markiv, A.; Anani, B.; Durvasula, R.V.; Kang, A.S. Module based antibody engineering: A novel synthetic REDantibody. J. Immunol. Methods 2011, 364, 40–49.
- Khan, N.A.; Shin, S.; Chung, J.W.; Kim, K.J.; Elliott, S.; Wang, Y.; Kim, K.S. Outer membrane protein A and cytotoxic necrotizing factor-1 use diverse signaling mechanisms for Escherichia coli K1 invasion of human brain microvascular endothelial cells. Microb. Pathog. 2003, 35, 35–42. Markiv, A.; Beatson, R.; Burchell, J.; Durvasula, R.V.; Kang, A.S. Expression of recombinant multi-coloured fluorescent antibodies in gor -/trxB- E. coli cytoplasm. BMC Biotechnol. 2011, 11, 117.
- Prasadarao, N.V.; Wass, C.A.; Weiser, J.N.; Stins, M.F.; Huang, S.H.; Kim, K.S. Outer membrane protein A of Escherichia coli contributes to invasion of brain microvascular endothelial cells. Infect. Immun. 1996, 64, 146–153. Fang, W.; Vega-Rodriguez, J.; Ghosh, A.K.; Jacobs-Lorena, M.; Kang, A.; St Leger, R.J. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science 2011, 331, 1074–1077.
- Killiny, N.; Rashed, A.; Almeida, R.P. Disrupting the transmission of a vector-borne plant pathogen. Appl. Environ. Microbiol. 2012, 78, 638–643. Bukhari, T.; Takken, W.; Koenraadt, C.J. Development of Metarhizium anisopliae and Beauveria bassiana formulations for control of malaria mosquito larvae. Parasites Vectors 2011, 4, 23.
- Lampe, D.J.; Lauzon, C.R.; Miller, T. Development of symbiotic control of Pierce’s Disease. 2016. Available online: https://biopesticide.ucr.edu/abstracts/assets/Lampe_abstract.pdf (accessed on 12 April 2020).Miller, T.A. Paratransgenesis as a potential tool for pest control: Review of applied arthropod symbiosis. J. Appl. Entomol. 2011, 135, 474–478.
- Miller, T. Symbiotic Control in agriculture and medicine. Comp. Biochem. Physiol. A Mol. Integr. Physiol. Comp. Biochem. Physiol. Pt A 2007, 146.
- Azizi, A.; Arora, A.; Markiv, A.; Lampe, D.J.; Miller, T.A.; Kang, A.S. Ribosome display of combinatorial antibody libraries derived from mice immunized with heat-killed Xylella fastidiosa and the selection of MopB-specific single-chain antibodies. Appl. Environ. Microbiol. 2012, 78, 2638–2647.
- Feil, H.; Feil, W.S.; Lindow, S.E. Contribution of Fimbrial and Afimbrial Adhesins of Xylella fastidiosa to Attachment to Surfaces and Virulence to Grape. Phytopathology 2007, 97, 318–324.
- De La Fuente, L.; Burr, T.J.; Hoch, H.C. Autoaggregation of Xylella fastidiosa cells is influenced by type I and type IV pili. Appl. Environ. Microbiol. 2008, 74, 5579–5582.
- Hoch, H.C. Continued assessment of Xylella fastidiosa fimbrial adhesins as important virulence factors in Pierce’s disease: Influence of xylem sap, p 87–91. In Proceedings of the 2010 Pierce’s Disease Research Symposium; Esser, T., Ed.; California Department of Food and Agriculture: Sacramento, CA, USA, 2010.
- Mohammed Akli Ayoub; Pascale Crépieux; Markus Koglin; Marc Parmentier; Jean-Philippe Pin; Anne Poupon; Eric Reiter; Martine Smit; Jan Steyaert; Hervé Watier; et al.Trevor Wilkinson Antibodies targeting G protein-coupled receptors: Recent advances and therapeutic challenges. Platform development for expression and purification of stable isotope labeled monoclonal antibodies in Escherichia coli 2017, 9, 735-741, 10.1080/19420862.2017.1325052.
- David C. Butler; Julie A. McLear; Anne Messer; Engineered antibody therapies to counteract mutant huntingtin and related toxic intracellular proteins. Progress in Neurobiology 2011, 97, 190-204, 10.1016/j.pneurobio.2011.11.004.
- Nives Škrlj; Marko Dolinar; New engineered antibodies against prions.. Bioengineered 2013, 5, 10-4, 10.4161/bioe.26069.
- Alessandro Angelini; Yoshishige Miyabe; Daniel Newsted; Byron H. Kwan; Chie Miyabe; Ryan L. Kelly; Misha N. Jamy; Andrew D. Luster; K. Dane Wittrup; Directed evolution of broadly crossreactive chemokine-blocking antibodies efficacious in arthritis. Nature Communications 2018, 9, 1461, 10.1038/s41467-018-03687-x.
- K.N. Westlund; M.A. Montera; A.E. Goins; S.R.A. Alles; M. Afaghpour-Becklund; R. Bartel; R. Durvasula; A. Kunamneni; Single-chain Fragment variable antibody targeting cholecystokinin-B receptor for pain reduction. Neurobiology of Pain 2021, 10, 100067, 10.1016/j.ynpai.2021.100067.
- Yaron Haviv; Yehuda Zadik; Yair Sharav; Rafael Benoliel; Painful Traumatic Trigeminal Neuropathy: An Open Study on the Pharmacotherapeutic Response to Stepped Treatment. Journal of Oral & Facial Pain and Headache 2013, 28, 52-60, 10.11607/jop.1154.
- Lene Baad-Hansen; Rafael Benoliel; Neuropathic orofacial pain: Facts and fiction. Cephalalgia 2017, 37, 670-679, 10.1177/0333102417706310.
- Karin N. Westlund; Marena A. Montera; Aleyah E. Goins; Sascha R. A. Alles; Nikita Suri; Sabrina L. McIlwrath; Robyn Bartel; Ravi V. Durvasula; Adinarayana Kunamneni; Single-Dose P2 X4R Single-Chain Fragment Variable Antibody Permanently Reverses Chronic Pain in Male Mice. International Journal of Molecular Sciences 2021, 22, 13612, 10.3390/ijms222413612.
- Robert E. Sorge; Josiane Mapplebeck; Sarah Rosen; Simon Beggs; Sarah Taves; Jessica K. Alexander; Loren Martin; Jean-Sebastien Austin; Susana G. Sotocinal; Di Chen; et al.Mu YangXiang Qun ShiHao HuangNicolas PillonPhilip J. BilanYu Shan TuAmira KlipRu-Rong JiJi ZhangMichael W. SalterJeffrey S. Mogil Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nature Neuroscience 2015, 18, 1081-1083, 10.1038/nn.4053.