Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Beatrix Zheng and Version 3 by Beatrix Zheng.
Cyanobacteria, also called blue-green algae, are a group of prokaryotic microorganisms largely distributed in both terrestrial and aquatic environments. They produce a wide range of bioactive compounds that are mostly used in cosmetics, animal feed and human food, nutraceutical and pharmaceutical industries, and the production of biofuels. Nowadays, the research concerning the use of cyanobacteria in agriculture has pointed out their potential as biofertilizers and as a source of bioactive compounds, such as phycobiliproteins, for plant pathogen control and as inducers of plant systemic resistance. The use of alternative products in place of synthetic ones for plant disease control is also encouraged by European Directive 2009/128/EC.
- cyanobacteria
- plant pathogens
- fungi
- oomycetes
- antifungal activity
- biocontrol
- plant-induced resistance
- plant biostimulants
- cyanobacteria cultivation
1. Introduction
Cyanobacteria are a group of ancient microorganisms that appeared about 2.6–3.5 billion years ago [1]. They were responsible for the Great Oxygenation Event [2] during the Paleoproterozoic era, which radically changed the composition of the Earth’s life forms and enabled the subsequent development of multicellular life forms [3]. It is assumed that during the evolution, plant plastids, cellular organelles in charge of photosynthesis, originated from cyanobacteria acquired through a process called endosymbiosis [4]. Cyanobacteria are also improperly called blue-green algae on the basis of their color, even though scientists have included them in the Cyanobacteria phylum of the bacteria domain. Cyanobacteria are the only prokaryota that can photosynthesize by using sunlight, water, and carbon dioxide to produce oxygen and energy. They form a monophyletic group with a high diversity in terms of morphology, physiology, and metabolic and molecular properties as a result of extensive time to colonize and adapt to evolving niches that emerge on Earth [1][5][6]. They are found worldwide in almost all environments (e.g., oceans, brackish water, freshwater, rocks, soil, plants, animals), many of which are extreme habitats where they play a prominent role [7][8][9][10].
Cyanobacteria have a high rate of adaptation to fluctuating environmental conditions, and pressure from competing organisms and grazers implies several survival mechanisms, including the production of secondary metabolites that are interesting for biotechnology applications [11][12][13]. Recent advances in genetic and metabolic engineering technologies have shown substantial progress in research aimed at realizing the full potential of cyanobacteria. Toxins naturally present in cyanobacterial cells may reach concentrations that pose a risk to human and animal health [14] due to the cyanobacteria’s ability to produce a wide range of bioactive compounds, some of which are considered toxic (cyanotoxins). By contrast, the property of producing substances with antiviral, antimicrobial, anticancer, antioxidant, and anti-inflammatory activity with potential beneficial effects on human health has long been known [14][15][16]. Additionally, cyanobacteria also produce a variety of enzymes (chitosanase, protease, xylanase, and cellobiase) with antifungal activity and twenty-four families of protease inhibitors involved in several human, animal, and plant metabolic pathways [14][17].
The interest in cyanobacteria is increasing, as shown by numerous studies in many fields, with more than 49,000 scientific documents that are cited in the Scopus database (Figure 1a). More than 1500 documents are cited in each of the following area: Agricultural and Biological Sciences, Biochemistry, Genetics and Molecular Biology, Environmental Science, Immunology and Microbiology, Earth and Planetary Sciences, Medicine, Chemistry, Pharmacology, Toxicology and Pharmaceutics, Multidisciplinary, Chemical Engineering, and Engineering (Figure 1b).
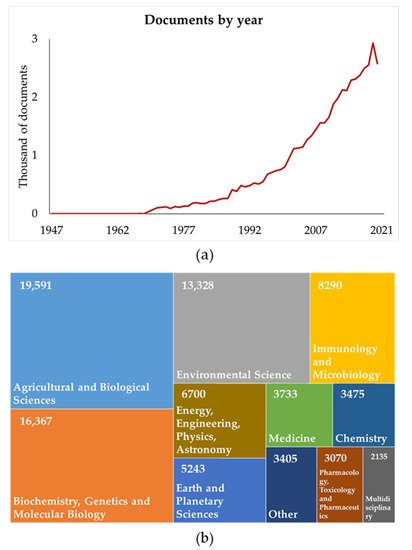
Figure 1. Scientific documents on cyanobacteria by year (a) and by subject area (b) reviewed in the Scopus® database (updated to October 2021). Other: Materials Science, Computer Science, Social Sciences, Mathematics, Neuroscience, Health Professions, Veterinary, Arts and Humanities, Decision Sciences, Economics, Econometrics and Finance, Business, Management and Accounting, Nursing, Psychology, Dentistry, Undefined.
As reported by the Scopus® database (Figure 1b), scientific reports on cyanobacteria primarily cover the agricultural and biological sciences topic area, although some topic areas may overlap. More specifically, most articles refer to the biostimulant effect, as these microorganisms are a source of macro- and micronutrients, bioactive molecules, and plant hormones that may positively influence the plant [18]. More recently, only a few dozen articles referring to the effect of cyanobacteria on plant pathogens were cited in the Scopus database (search for “cyanobacteria + versus + plant pathogens”). Cyanobacteria have been shown to reduce the infection of several fungal pathogens by acting directly against pathogen growth and indirectly stimulating plant defense responses [19][20][21][22]. Recent studies have shown that Anabaena minutissima treatment elicits tomato root defense barriers through the production of phenolic compounds [20], protects cutin and pectin structures in tomato fruits from the etiological agent of gray mold disease (Botrytis cinerea) [23], and elicits the expression of pathogenesis-related genes in cucumber plants [24]. Consequently, cyanobacteria may potentially be an effective alternative to synthetic agrochemicals, which are currently the principal products for plant protection against pathogens, notably fungi and oomycetes. This is consistent with national and international regulations limiting the use of synthetic agrochemicals due to their harmful consequences on human and animal health and the environment.
2. Plant Pathogen Control
The present study describes current knowledge on the activity described for cyanobacteria against a wide range of phytopathogenic fungi and oomycetes that attack agricultural plants.
2.1. Plant Pathogens
Plant pathogens are disease-causing agents belonging to several species of fungi, oomycetes (fungal-like organisms), bacteria, phytoplasmas, viruses, and nematodes. They are largely distributed in the environment and can affect plant root system, stem, leaf, and fruit of several crops in all cultivation systems causing considerable economic losses. The FAO estimation (2019) of plant diseases cost for the global economy is around USD 220 billion per year, with 20–40% of crop production lost to pests. Among the causal agents of infectious diseases, fungi are responsible of most of the diseases occurring in all agricultural crops [25]. Dean et al. [26] reviewed top 10 fungal plant pathogens on the base of scientific and economic relevance, such as Botrytis cinerea, Fusarium oxysporum, and Colletotrichum spp. Fungal diseases also constitute a concern for food security, because some fungi, as reported below, produce mycotoxins that are harmful for both human and livestock [27][28][29]. Oomycetes also include some of the most destructive plant pathogens, such as Phytophthora infestans, that is one of the major pathogens in potato production and Pythium spp. agents of damping-off in nursery [30]. Taking into account the above statements this review deals with cyanobacteria as biocontrol agents of plant pathogenic fungi and oomycetes.
In this section, both fungi and oomycetes studied for the activity of cyanobacteria are presented. Among fungi, Aspergillus and Fusarium species are the most considered. Aspergillus species are worldwide saprotrophic and pathogenic fungi. Many species that infect plants in the field can cause visible symptoms only during the postharvest stages. They attack fresh fruits and vegetables, dry fruit, grains, and cereal products. Many strains of Aspergillus produce mycotoxins, such as ochratoxin A (A. carbonarius, A. niger, A. parasiticus, and A. westerdijkiae), and aflatoxins (A. flavus) which are high toxic to mammals [27][31]. Some species are also opportunistic human and animal pathogens [32]. Fusarium oxysporum is a representative example of Fusarium species for studying cyanobacteria extracts activity. Fusarium oxysporum is a species complex including more than 150 different formae speciales adapted to infect specific host plants [33]. They are ubiquitous soil borne pathogens, responsible for a wide range of plant diseases and usually causing a vascular wilt with host-specific forms [34]. These pathogens penetrate the root epidermis and colonize the xylem vessels resulting in plant wilt and yellowing symptoms causing considerable crop losses [35]. Their control is difficult due to the lack of effective fungicides; hence, the research of new alternative solutions is gaining attention in sustainable agriculture. Infection by F. oxysporum can also be harmful to humans and animal health [36]. Other plant pathogens tested for the activity of cyanobacterial extracts are the fungi Penicillium expansum, P. verrucosum, Penicillium sp., Macrophomina phaseolina, Alternaria solani, A. alternata, Drechslera oryzae, Rhizoctonia solani, Botrytis cinerea, Armillaria sp., Rosellinia sp., Sclerotinia sclerotiorum, Verticillium albo-atrum, Colletotrichum gleosporioides, and C. musae. It is noteworthy that P. expansum has a wide host range, causing mainly post-harvest rot symptoms on fruits and produces the neurotoxin patulin, which can be taken by humans with apples and apple products [28]. Concerning oomycetes, Pythium aphanidermatum, P. debaryanum, P. ultimum, Phytophthora cambivora, P. capsici, and P. cinnamomic are studied for cyanobacteria activity. These pathogens survive in soil and can cause damping off in seedlings, root and crown rot, stunted growth, and wilting in many older agricultural plants. Moreover, P. capsici can infect leaf and fruit of major crop hosts (tomato, eggplant, all cucurbits, snap, and lima beans) [37] and P. cinnamomi together with P. cambivora can be pathogens of several forest trees [35].
Important fungal pathogens globally distributed are the soil borne Rhizoctonia solani and the foliar pathogens Botrytis cinerea and Podosphaea xanthii. Rhizoctonia solani causes damping-off of seedlings, root rot, and leaf blight [38] and under unfavorable environmental conditions produces resistant structures that can survive in the soil for several years and on debris from various cultivated plants [35]. Botrytis cinerea is one of the most widely studied fungal pathogens and causes serious disease, namely gray mold disease, on various economically important crops including strawberry, tomato, and grape [26]. This fungus is very aggressive and can infect stems, leaves, flowers, and fruits in pre- and post-harvest [39]. Botrytis cinerea has been recognized as one of the most important post-harvest pathogens in both fresh fruits and vegetables [26]. Podosphaera xanthii is the main causal agent of cucurbits powdery mildew spread worldwide [40]. Typical disease symptoms are whitish powdery mass mainly composed by mycelium and spores that mainly appear on leaf and stems. Under favorable environmental conditions, fungal colonies cover the entire upper leaf surface resulting in chlorosis and early senescence [41]. A worldwide pathogen causing severe damage on one of the most important crops is Magnaporthe oryzae, the agent of rice blast disease. This fungus causes necrotic symptoms on all parts of the plant and is responsible of 10–30% rice global yield losses [42].
2.2. Cyanobacteria as Biocontrol Agents
2.2.1. In Vitro Studies and Mechanisms of Fungal and Oomycete Growth Inhibition
Many research papers demonstrate the activity of cyanobacterial extracts against fungal and oomycetes pathogens in agar medium assay (Table 1). Nostocales and Oscillatoriales orders are the most studied since the early 2000. Several extracts from Microcystis aeruginosa, Anabaena spp., Fischerella sp., Nostoc spp., Scytonema spp., Lyngbya lutea, Oscillatoria spp., Phormidium tenue, Trichodesmium hildebrantii, Synechococcus elongates, and Synechocystis sp. Inhibited Aspergillus spp. Mycelial growth in agar disk diffusion assay [43][44][45][46]. Fungal inhibition has consistently been shown by the methanol extract of these cyanobacteria [43][44][46] and in some cases, depending on cyanobacterium species vs Aspergillus species, by extracts with acetone, diethyl ether, ethyl acetate, ethanol, methyl chloride, n-propanol, petroleum, and petroleum ether [43][46]. Activity against Aspergillus spp. Was also obtained with aqueous extract from Lyngbya lutea, Oscillatoria spp., Phormidium tenue, and Synechocystis sp. [46], and with Nostoc muscorum culture filtrate [45].
Regarding the growth reduction in Fusarium species, it was obtained by Microcystis aeruginosa, Anabaena spp., Calothrix brevissima, Nodularia sp., Nostoc spp., Lyngbya lutea, Oscillatoria spp., Phormidium spp., Trichodesmium hildebrantii, Synechococcus elongates, and Synechocystis sp. [17][21][22][43][45][46][47][48][49][50]. More specifically, the methanol extract from most cyanobacteria species (Nostoc spp., Lyngbya lutea; Oscillatoria spp., Phormidium spp., Trichodesmium hildebrantii, Synechococcus elongates, and Synechocystis sp.) showed efficacy in reducing Fusarium growth [46][47][48][49][50]. Extracts obtained with other organic solvents, such as n-propanol, petroleum ether, aceton, methyl chloride, diethyl ether, and ethyl acetate were also effective against Fusarium spp. [43][46]. Moreover, culture filtrates from several species of Anabaena and an aqueous extract from Phormidium tenue reduced Fusarium spp. growth [21][22][46]. In addition to inhibiting the growth of Aspergillus and Fusarium species, cyanobacteria extracts have shown efficacy against other plant pathogens (Table 1).
Table 1. Activity of extracts and culture filtrates from cyanobacteria against phytopathogenic fungi and oomycetes in agar medium assays.
| Cyanobacterium | Extract/Culture Filtrate 1 | Plant Pathogen | Reference |
|---|---|---|---|
| Chroococcales | |||
| Microcystis aeruginosa | ME | Aspergillus carbonarius, A. niger | [43] |
| ETH | A. flavus, A. niger, A. parasiticus | ||
| AC | A. flavus, A. niger, Fusarium proliferatum | ||
| MC | A. flavus, A. parasiticus, F. proliferatum | ||
| DE | A. carbonarius, A. flavus, A. niger, A. ochraceus, A. westerdijkiae, F. proliferatum, F. verticillioides, Penicillium verrucosum | ||
| EA | A. carbonarius, A. flavus, A. niger, A. westerdijkiae, F. verticillioides | ||
| Nostocales | |||
| Anabaena spp. | CFILT | Alternaria solani, Drechslera oryzae, Fusarium moniliforme, F. solani, Macrophomina phaseolina, Pythium aphanidermatum | [17] |
| Anabaena sp. | PE | Alternaria alternata | [47] |
| Anabaena sp. | ME | Aspergillus flavus | [44] |
| A. cylindrica | ME | A. flavus | [44] |
| A. oscillarioides | CFILT, L | F. moniliforme, F. oxysporum f. sp. lycopersici, Pythium debaryanum, Rhizoctonia solani | [21] |
| A. solitaria | ME | Alternaria alternata | [47] |
| A. variabilis | CFILT, L | F. moniliforme, F. oxysporum f. sp. lycopersici, P. debaryanum, R. solani | [21] |
| A. variabilis | CFILT | F. oxysporum f. sp. lycopersici | [22] |
| A. laxa clones | CFILT | Pythium aphanidermatum | [51] |
| Calothrix brevissima | PE | Alternaria alternata | [47] |
| ME | A. alternata, Botrytis cinerea, F. oxysporum | [47] | |
| Fischerella sp. | ME | Aspergillus flavus | [44] |
| Nodularia sp. | ME | F. oxysporum | [47] |
| Nostoc sp. | ME | A. flavus | [44] |
| Nostoc strain ATCC 53789 | ME | Armillaria sp., Fusarium solani, F. oxysporum f. sp. melonis, Penicillium expansum, Phytophthora cambivora, P. cinnamomi, Rhizoctonia solani, Rosellinia sp., Sclerotinia sclerotiorum, Verticillium albo-atrum | [48] |
| Nostoc strain UTEX 2493 | ME | Rosellinia sp. | [48] |
| N. calcicula | ME | Aspergillus flavus | [44] |
| N. commune | ME | F. oxysporum f. sp. lycopersici | [50] |
| N. commune | PE | Phytophthora capsici, Pythium ultimum | [47] |
| ME | F. oxysporum, P. capsici | [47] | |
| N.commune | PE | Alternaria alternata | [47] |
| N.commune | ME | A. niger | [46] |
| PE | A. flavus, A. niger | [46] | |
| N.entophytum | AC, CHL, ME | R. solani | [52] |
| N. linckia | ME | F. oxysporum f. sp. lycopersici | [49] |
| N.muscorum | ME | A. alternata, B. cinerea, Colletotrichum gleosporioides | [47] |
| N.muscorum | CFILT | Aspergillus flavus, A. niger, Fusarium microsporium, Penicillium sp. | [45] |
| N.muscorum | AC, CHL, ME | R. solani | [52] |
| Scytonema sp., S. hofmanni |
ME | A. flavus | [44] |
| Oscillatoriales | |||
| Arthrospira platensis | PBPs | B. cinerea | [53] |
| Lyngbya lutea | W | A. niger | [46] |
| ME | A. niger, Colletotrichum musae, F. oxysporum | ||
| nPROP | A. flavus, F. oxysporum | ||
| PEE | A. niger, C. musae, F. oxysporum | ||
| Oscillatoria amphibia | W | A. flavus, C. musae | [46] |
| ME | F. oxysporum | ||
| nPROP | A. flavus, C. musae, F. oxysporum | ||
| O. angustissima | PE | C. gleosporioides, F. oxysporum | [47] |
| O. limosa | W | A. niger, C. musae | [46] |
| ME | A. flavus, A. niger, C. musae, F. oxysporum | ||
| nPROP | A. flavus, A. niger, C. musae, F. oxysporum | ||
| O. ornata | ME | A. flavus, A. niger, C. musae, F. oxysporum | |
| nPROP | C. musae, F. oxysporum | ||
| PEE | A. niger | ||
| O. tenuis | PE | A. alternata, P. capsici | [47] |
| ME | P. capsici | ||
| Phormidium autumnale | ME | F. oxysporum f. sp. lycopersici | [49] |
| P. tenue | W | A. niger, C. musae, F. oxysporum | [46] |
| ME | A. niger, F. oxysporum | ||
| nPROP | C. musae, F. oxysporum | ||
| PEE | A. niger, P. lilacimus | ||
| Trichodesmium hildebrantii | W | C. musae | [46] |
| ME | A. niger, C. musae, F. oxysporum | ||
| nPROP | A. flavus, A. niger, C. musae, P. lilacimus | ||
| PEE | C. musae, F. oxysporum | ||
| Synechococcales | |||
| Synechococcus elongates | W | C. musae | [46] |
| ME | A. niger, C. musae, F. oxysporum | ||
| nPROP | A. flavus, A. niger, P. lilacimus | ||
| PEE | C. musae, F. oxysporum | ||
| Synechocystis sp. | W | A. flavus, A. niger, C. musae, P. lilacimus | [46] |
| ME, nPROP | A. flavus, A. niger, C. musae, P. lilacimus, F. oxysporum | ||
| PEE | C. musae |
1 Solvent used for extraction: AC, acetone; CHL, chloroform; DE, diethyl ether; EA, ethyl acetate; ETH, ethanol; ME, methanol; MC, methyl chloride; PE, petroleum; PEE, petroleum ether; W, water; nPROP, n-propanol. CFILT, culture filtrate; L, leachates from compost amended with viable cells; PBPs, phycobiliproteins.
Among Nostocales, methanol extracts from Nostoc spp. were efficient in reducing the colony growth of the fungi Armillaria sp., Rosellinia sp., Sclerotinia sclerotiorum, Penicillium expansum, Verticillium albo-atrum, and R. solani and of the oomycetes Phytophthora cambivora and P. cinnamomi [48][52]. Rhizoctonia solani and the oomycete Pythium debaryanum were also inhibited by Anabaena variabilis and A. oscillarioides culture filtrates and leachates from compost amended with each cyanobacterium viable cells [21]. Prasanna et al. [17] demonstrated that Anabaena spp. culture filtrates were effective against the fungi Macrophomina phaseolina, Drechslera oryzae, and Alternaria solani and against the oomycete Pythium aphanidermatum growth, while methanol extracts of Anabaena solitaria, Calothrix brevissima, and Nostoc muscorum reduced A. alternata [47]. The last two extracts showed efficacy also against B. cinerea, while the extract of N. commune was effective against Phytophthora capsici [47]. Culture filtrates of Anabaena laxa and N. muscorum reduced the oomycete P. aphanidermatum and the fungus Penicillium sp. growth, respectively [45][51]. The growth of a different species of Penicillium, P. verrucosum, was also reduced by a diethyl ether extract from Microcystis aeruginosa that belongs to Chroococcales order [43]. Among the extract of Oscillatoriales order, colony growth reduction in both fungi and oomycetes was obtained. A recent study showed that phycobiliproteins extracted from A. platensis hampered the germination of B. cinerea spores by inhibiting spore germination and mycelial development [53]. This pathogen produces a vast quantity of airborne spores which are the main fungus dispersal mean, therefore, the inhibition of spore germination can play a role in disease control by interrupting secondary infections. The antifungal activity of other kinds of proteins has been also demonstrated on B. cinerea spore germination, as well as on mycelial growth [54]. Arthrospira platensis proteins are characterized by a secondary α-helix structure. This protein structure, being prominently represented in transmembrane proteins, could interact with the fungal cell wall, leading to the perturbation of cell activity [53]. A petrol extract from Oscillatoria angustissima inhibited Colletotrichum gleosporioides [47], while several kinds of extract from Oscillatoria spp., Phormidium tenue and Trichodesmium hildebrantii reduced the growth of Colletotrichum musae [46]. Moreover, Oscillatoria tenuis petroleum and methanol extracts reduced the growth of the oomycete Phytophthora capsici [47]. Colletotrichum musae was also inhibited by aqueous, methanol or petroleum ether extracts from the Synechococcales Synechococcus elongates and Synechocystis sp. [46].
The antifungal activity shown in Table 1 is mainly referred to cyanobacteria extracellular culture filtrates [17][21][22][51] and to extracts obtained from cyanobacteria with different organic solvents, such as acetone, chloroform, diethyl ether, ethyl acetate, ethanol, methanol, methyl chloride, n-propanol, petroleum and petroleum ether [17][21][22][43][44][45][46][47][48][49][50][51][52], as well as water [46]. The culture filtrates are rich of many substances that can display interesting antifungal activity. In particular, up to 85 families of metabolites with high antimicrobial activity were isolated from various strains of cyanobacteria [14]. Culture filtrates of several Anabaena species with fungicidal activity produced one or more hydrolytic enzymes, such as proteases, chitosanases, exo-β-1,4-glucanases, and carboxy-methyl cellulase [17][51]. For Anabaena variabilis ATCC 29413, the gene putatively responsible for chitosanase and antifungal activities was attributed to the glycoside hydrolase 3-like family [51]. All these enzymes are known to be involved in the digestion of fungal or oomycetes cells. For example, chitosan and chitosan-glucan complexes were found in the mycelia of Aspergillus niger and Fusarium moniliforme [51]. The use of organic solvents in the extraction process, affects the antifungal activity of cyanobacterial extracts [46]. For example, in the extract of Microcystis aeruginosa obtained with diethyl ether were identified the butylated hydroxytoluene and methyl ester of hexadecanoic acid, which have antifungal activity against Aspergillus spp., Fusarium spp. and Penicillium sp. [43]. Another important antifungal activity against A. flavus was observed in the methanol extract of Anabaena spp., Nostoc sp. and Scytonema sp. The identification of the macrolide scytophycin and the presence of the glycolipopeptide hassallidin extracted from Anabaena strains elucidated the antifungal activity [44]. Osman et al. [52] suggested that phenols and polysaccharides contained in extracts from Nostoc spp. are involved in the antifungal activity against R. solani. From a previous study [45], a phenolic compound was isolated and purified from the chloroform extract of Nostoc muscorum with strong activity against Aspergillus niger, A. flavus, Pencillium sp., and Fusarium microsporium. Aqueous extracts have not been so widely investigated as the extracts obtained with organic solvents, even though they are safer for both human and environment. A recent study showed that soluble polysaccharides extracted from Anabaena minutissima aqueous extract reduced both colony growth and colony-forming units of B. cinerea [55]. In the same extract, proteins, phycobiliproteins, chlorophylls, carotenoids, and antioxidant activities were also determined and correlated with the antifungal effect against the pathogenic fungus Podosphaera xanthii on cucumber detached cotyledons in in vitro assay [24].
2.2.2. In Vivo Studies and Mechanisms of Biocontrol Ability
Most of the examples in Table 2 shows that extracts and compounds from cyanobacteria belonging to order of Nostocales are active against fungal pathogens mainly, whereas those from Oscillatoriales showed activity against fungi only.
Among Nostocales, the Anabaena species are widely studied. Both A. minutissima and A. variabilis were active in controlling foliar, soil-borne, or fruit pathogens [20][21][22][23][24][55][56]. An aqueous extract from A. minutissima sprayed on cucurbit plants reduced powdery mildew symptoms caused by P. xanthii under greenhouse conditions. On cucumber, the treatment reduced the diseased leaf area by 31% and the sporulation (spores/mm2) by 47% [24], while on zucchini the disease reduction was 25% [56]. These activities were attributed to carotenoids, polysaccharides, chlorophylls, and proteins that were determined in the extract. The same extract applied on both cucurbits was able to elicit plant defense systemic responses in which enzyme activities and different expression of various pathogenesis related (PR) protein genes are involved (Figure 2). Plant responses to A. minutissima treatment are jasmonic acid (JA) and salicylic acid (SA) dependent. Indeed, activity and expression of total chitinases, β-1,3-glucanases and peroxidases, and AePR3 and PR1 gene expression were enhanced [24][56]. The elicitation of seedling defense responses might be correlated with the substances contained in the aqueous extract. It is also noteworthy that the same A. minutissima extract was a suitable mean for tomato seed treatment in controlling the soil-borne pathogen Rhizoctonia solani [20]. Additionally, on tomato, the extract elicited plant defense responses that were revealed by the increasing of chitinase activity and by lignin deposition in root seedlings. The cyanobacterium A. minutissima is also a source of bioactive compounds, such as polysaccharides and phycobiliproteins (PBPs). Polysaccharides applied by pre-harvest treatment to strawberry fruits reduced Botrytis cinerea symptoms and sporulation by 50% and 67%, respectively [55]. As regard PBPs, they were able to reduce disease incidence and severity when applied on tomato fruits in a small-scale experiment [23]. The characterization of the tomato fruits by FT-IR and FT-Raman spectroscopic techniques revealed that A. minutissima PBPs preserved cutin and pectin structures by B. cinerea invasion. Concerning the effect of A. variabilis against pathogens, Chaudhary et al. [21] obtained the control of several fungi and of the oomycete P. debaryanum on tomato seedling by soil application of paddy straw compost amended with the cyanobacterium viable cells (Table 2). The same strain of A. variabilis applied as viable cells in a paddy straw compost-vermiculite was tested for suppressing wilt disease by F. oxysporum f. sp. lycopersici on tomato seedling [22]. A low disease severity was observed, along with an increase in β-1,3 glucanases, polyphenol oxidases (PPO) and phenylalanine ammonia lyase (PAL) (Figure 2). Plant defense responses, such as peroxidases, and β-1,3 glucanases were also triggered in pathogen-unchallenged coriander, cumin, and fennel plants by another Anabaena species, A. laxa, and by Calothrix elenkinii, which were applied as biomass culture in a potting mix under controlled conditions [57]. Moreover, extracts from the same plants showed fungicidal activity against F. oxysporum in in vitro assay, suggesting a role of the plant enzymatic activities in the alteration of pathogen cell wall and membrane structures and functions.
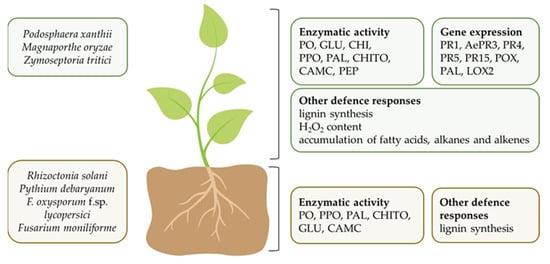
Figure 2. Foliar and soil-borne pathogens controlled by cyanobacteria application and plant defense responses in shoot and root induced by the cyanobacteria reported in this review. CAMC, carboxymethyl cellulose; CHI, chitinases; CHITO, chitosanase; GLU, β-1,3-glucanases; LOX, 13-lipoxygenase 2; PAL, phenylalanine ammonia lyase; PEP, phosphoenolpyruvate carboxylase; PO, peroxidases; PPO, polyphenol oxidases; PR, pathogenesis related proteins; WRKY, family of transcription factors.
Again on Nostocales (Table 2), Nostoc linkia added to soil controlled the soil-borne pathogen F. oxysporum f. sp. lycopersici in pot experiments [49]. In a more in-depth analysis regarding the mechanisms of Nostocales, the liquid medium culture of N. punctiforme showed to protect Arabidopsis thaliana cells from programmed cell death (PCD). The reduction in PCD by N. punctiforme was preceded by the induction of several WRKY family transcription factors [58]. This gene family is widely distributed in plants and play a key role in many metabolic pathways, including biotic and abiotic stress responses [59]. Soil drench treatment with viable cells of a consortium of Nostoc and Anabaena species, as well as C. elenkinii alone isolated from the rice phyllosphere was found effective against Magnaporthe oryzae on rice [60]. The same authors also demonstrated that these biocontrol agents were able to colonize plants, in particular the phyllosphere as resulted by the abundance of 16S genes rRNA copies of the cyanobacterial communities. An important point to underline is that the isolates of Nostoc spp., Anabaena spp., and C. elenkinii have the same habitat of the pathogen M. oryzae, this suggesting a potential pathogen control on rice leaf. The effective treatments resulted in an increase in the leaf antioxidant enzymes PPO, PAL, peroxidases, phosphoenol pyruvate carboxylase activities involved in plant defense induction, for C. elenkinii, and an increase in only PPO for the consortium [60]. Again, on rice, the C. elenkinii inoculation in the seedling growth medium enhanced shoot and root defense enzyme activities, PPO, PAL, peroxidases, chitosanase, β-1,3 glucanases, and carboxymethyl cellulose [61] (Figure 2).
Table 2. Pathogen control and plant defence induction by cyanobacterial extracts and their single compounds applied on several plant species with different kind of treatments.
| Cyanobacterium | Extract/ Biomass/ Compound 1 |
Plant/ Treatment 3 |
Pathogen Control 4/ Plant Defense Responses 5 |
Reference |
|---|---|---|---|---|
| Nostocales | ||||
| Anabaena laxa | Biomass culture |
Coriander GS |
Shoot and root: PO activity; shoot: GLU activity | [57] |
| Cumin GS |
Shoot and root: PO activity | |||
| Fennel GS |
Shoot: PO activity | |||
| A. minutissima | W | Zucchini L |
Podosphaera xanthii (25%) CHI, GLU, PO activity, isoforms of CHI, GLU, PO |
[56] |
| W | Cucumber L |
P. xanthii (31%) PR1, AePR3 genes |
[24] | |
| W | Tomato S |
Rhizoctonia solani Seedling: CHI activity, lignin content |
[20] | |
| PBPs | Tomato F |
Botrytis cinerea cutin and pectin preservation |
[23] | |
| POL | Strawberry F |
B. cinerea | [55] | |
| A. variabilis | Biomass 2 | Tomato GS |
Pythium debaryanum, R. solani, Fusarium moniliforme, F. oxysporum f. sp. lycopersici | [21] |
| A. variabilis | Biomass 2 | Tomato seedling |
F. oxysporum f. sp. lycopersici (100%) GLU, PPO, PAL activity |
[22] |
| Calothrix elenkinii | Biomass culture |
Coriander GS |
Shoot and root: PO activity | [57] |
| Cumin GS |
Shoot and root: PO activity | |||
| Fennel GS |
Shoot: PO, GLU activity | |||
| C. elenkinii | Biomass | Rice GS |
Shoot and root: PPO, PAL, PO, CHITO, GLU, CAMC activity | [61] |
| C. elenkinii | Biomass | Rice GS |
Magnaporthe oryzae (50%) Leaf: PO, PPO, PAL, PEP activity |
[60] |
| Nostoc linkia | Biomass | Tomato GS |
F. oxyporum f. sp. lycopersici | [49] |
| N. punctiforme | Medium culture | Arabidopsis thaliana | WRKY | [58] |
| Nostoc-Anabaena consortium | Biomass | Rice GS |
Magnaporthe oryzae (69%) PPO activity |
[60] |
| Oscillatoriales | ||||
| Arthrospira platensis | DB | Wheat L |
Zymoseptoria tritici (~70%) PR4, PR5, PR15, PO, PAL, LOX genes |
[62] |
| A. platensis | PBPs | Tomato F |
B. cinerea | [53] |
| A. platensis | POL | Tomato L |
PAL, CHI, GLU, PO activity; H2O2 content; accumulation of fatty acids, azelaic acid, alkanes, alkenes, other metabolites | [63] |
1 W, water extract; DB, dry biomass; PBPs, phycobiliproteins; POL, polysaccharides. 2 Biomass applied as viable cells in paddy straw compost. 3 L, leaf; S, seed; F, fruit, GS, plant growth substrate. 4 Disease reduction. 5 CAMC, carboxymethyl cellulose; CHI, chitinases; CHITO, chitosanase; GLU, β-1,3-glucanases; LOX, 13-lipoxygenase 2; PAL, phenylalanine ammonia lyase; PEP, phosphoenolpyruvate carboxylase; PO, peroxidases; PPO, polyphenol oxidases; PR, pathogenesis related proteins; WRKY, family of transcription factors.
Concerning Oscillatoriales order, Arthrospira platensis (synonymous Spirulina platensis) sprayed on plants as dry biomass suspended in water, protected wheat plants from Zymoseptoria tritici in a similar way of the commercial product Bion® 50 WG, which is based on the elicitor acibenzolar-s-methyl [62]. In addition to the efficacy against this pathogen (about 70% with respect to the control), the treatment elicited wheat defenses by upregulating PR4, PR5, PR15 (oxalate oxidase), PAL, peroxidases, and mainly 13-lipoxygenase 2 genes. More recent research showed that A. platensis PBPs applied before Botrytis cinerea inoculation, controlled the pathogen on tomato fruit in a small-scale experiment. The PBPs characterization revealed an α-helix secondary structure that is a class including transmembrane proteins, probably involved in the perturbation of fungal cell activity [53]. In tomato leaves, A. platensis polysaccharides induced many biochemical changes correlated to plant defense (PAL, chitinases, glucanases and peroxidases activities and H2O2 accumulation) and involved in the construction of wax and cutin, such as fatty acids (C16:3, C18:2 and C18:3, C18:0) and azelaic acid, alkanes (eicosane, tetracosane, octacosane, nonacosaene, triacontane, dotriacontane; the alkane derivate 1-chloroeicosane) and alkenes (1-octadecene, 1-pentadecene) [63]. Other new metabolites (2(4H)-benzofuranone, 5,6,7,7a-tetrahydro-4,4,7a-trim, neophytadiene, alpha-tocospiro-B, tris(2,4-di-tert-butylphenyl) phosphate) were also detected in the same leaves.
3. Biostimulant Effects
Recently, cyanobacteria have gained considerable attention as beneficial bioagents based on their ability to produce biomass for biofertilizers to be used in sustainable farming [18][64][65]. These organisms have been considered beneficial to soil fertility and crops through their ability to fix atmospheric nitrogen solubilize phosphate [66], produce bioactive substances, such as hormones (auxins, gibberellins, cytokinins) [67], polypeptides, amino acids [68], polysaccharides [69], and siderophores [70], having stimulatory effects on plants. Bioactive substances influence the expression of many genes in host plants, thereby boosting plant growth and helping plants to counteract both biotic and abiotic stresses [18][64][65]. In general, there is an improvement in plant physiological processes (Table S1), which facilitates nutrient uptake, yield, or fruit quality, while also improving the healthy environment [64]. Thus, the application of cyanobacteria in agriculture and environmental management includes: (i) economic benefits (lower production costs), (ii) improved nutrient cycling, (iii) water storage, and (iv) prevention of pollution and soil degradation, primarily through reduction in agrochemical use, nutrient recycling, and restoration of soil fertility through remediation [71]. The plant response is considerably influenced by the amount and concentration of bioactive molecules present in cyanobacteria which usually depends on the species and their growth conditions [64]. Several studies have found a dose-dependent response between cyanobacteria concentration and plant growth. As a consequence, the effects on plants are greater at low concentrations in terms of a higher fresh weight and chlorophyll content in radish seedlings after application of A. platensis [72]. Given the importance of cyanobacteria as biostimulants, they can play a key role in addressing sustainability challenges, as they can reduce mineral fertilizer inputs by encouraging the development of environmentally sustainable technologies.
The global market of biostimulants was valued at USD 2.6 billion in 2019 and is forecast to grow by 11.24% until 2025 [73]. In addition, the price of biostimulants ranges between EUR 10 and 80, and it varies by crop type, application timing, and dosage; the price of the application varies between EUR 100 and 600 per hectare [74]. The production of cyanobacterial biostimulants is limited, as these microorganisms are not currently listed in EU Regulation 2019/1009. Nerveless, Spain is the European Union leader in the cultivation and marketing of Arthrospira spp. as biostimulants. Thus, cyanobacteria-based biostimulant market remains a very small niche compared to the very high number of algae-based products that account for over 33% of the global market worldwide [75]. The updated list of cyanobacterial biostimulants currently on the market is reviewed by Santini et al. [18].
Nowadays, the cultivation of cyanobacteria for plant biostimulant production is quite expensive depending on the cultivation technology used (natural or artificial light, photobioreactors, or open ponds). Santini et al. [18] quote an extensive marketing analysis on the cost of treatment with cyanobacterial biostimulants, ranging from EUR 20 to 375 per hectare, depending on doses and number of applications. To make these biostimulants more market competitive, the cost of biomass production has to be reduced. To overcome some of the cost constraints of the process technology, an integrated biorefining concept should be applied to cyanobacterial biostimulant production [76]. This concept considers the sustainability of the entire process, from the use of non-potable water and recovery of nutrients from wastewater (C, N, P) to eco-friendly end products, such as high-value compounds or the entire biomass [77] (SABANA H2020 Project). This also involves the reduction in the dehydration process, with the biomass being used directly in the process (SABANA H2020 Project). In particular, molecules with biostimulant activity may be extracted in biorefining process and the residual waste may be utilized as biofertilizer [78]. On the other hand, controlled cultivation and adaptability of cyanobacteria metabolism can provide a wide range of options for quality improvement and standardization of biostimulant production.
The application of cyanobacteria encompasses direct inoculation with living cells or extracts obtained by chemical, enzymatic, and mechanical/physical extractions, such as autoclaving, drying, heating with water after sonication, and cell destruction (grinding with pestle and blender) [18]. However, there is general evidence that the extraction method greatly influences the composition and bioactivity of the extracts. In general, cyanobacterial extraction procedures are more advantageous than using viable biomass as they allow for greater recovery of active ingredients contained in the cell or bound to the cell wall [18].
In order to provide cyanobacteria extracts or viable cells to crops, several application methods were adopted, depending on the cyanobacteria and their formulation. The most common treatments include: (i) soil amendment with dry biomass (e.g., powder, granules, or pellets) or suspended liquid culture, and (ii) foliar application by spraying leachate and supernatant [79]. The foliar spray application appears to be more efficient under high relative humidity conditions as leaf stomata are open, thus the product absorption is maximized.
Particularly interesting is the biostimulant effect of cyanobacteria on root development. Studies on several treated seedlings with cyanobacterial extracts showed significant changes in root architecture (higher total root length, surface, and number of root tips). The changes induced by cyanobacterial extracts reflected an enhancement in nutrients, water uptake and plant growth as observed on lettuce [80], tomato [81], red beet [80], and cucumber [82]. It is known that cyanobacteria excrete bioactive substances that act as signaling molecules, promoting plant growth and the synthesis of phytohormones such as IAA auxin, cytokinins, gibberellins, ethylene, jasmonic or abscisic acids [83][84]. These substances influence diverse physiological processes in plants, including stimulation of root growth and branching, seedling growth, flowering, and fruit ripening [85]. In addition, an influence is expected at the molecular level with an upregulation of several genes involved in the primary and secondary metabolic pathways [86].
Considering the large number of scientific papers on the beneficial effects of cyanobacteria on a variety of vegetables and crops, few studies have been conducted to identify the singular chemical constituents responsible for stimulating plant growth. Given the complex molecular mixtures present in cyanobacteria extracts, the identification of the role of single molecules in biological activity is very hard. However, synergistic action of several compounds in the mixture is known to boost the biostimulant activity [18]. Accordingly, research effort has been directed to identifying the mechanisms of action of cyanobacterial extracts, given their positive impact on plant productivity via enhancement of processes, such as photosynthesis, nutrient and water uptakes, and activation of resistance genes to abiotic stresses.
References
- Schopf, J.W. The fossil record of cyanobacteria. In Ecology of Cyanobacteria II; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 15–36.
- Blankenship, R.E. Origin and early evolution of photosynthesis. Photosynth. Res. 1992, 33, 91–111.
- Buick, R. The antiquity of oxygenic photosynthesis: Evidence from stromatolites in sulphate-deficient archaean lakes. Science 1992, 255, 74–77.
- Barsanti, L.; Gualtieri, P. General Overview. In Algae: Anatomy, Biochemistry, and Biotechnology; Barsanti, L., Gualtieri, P., Eds.; CRC Press Taylor & Francis: Boca Raton, FL, USA, 2005; pp. 1–6.
- Dvořák, P.; Casamatta, D.A.; Hašler, P.; Jahodářová, E.; Norwich, A.R.; Poulíčková, A. Diversity of the Cyanobacteria. In Modern Topics in the Phototrophic Prokaryotes; Hallenbeck, P., Ed.; Springer: Cham, Switzerland, 2017; pp. 3–17.
- Demoulin, C.F.; Lara, Y.J.; Cornet, L.; François, C.; Baurain, D.; Wilmotte, A.; Javaux, E.J. Cyanobacteria evolution: Insight from the fossil record. Free Radic. Biol. Med. 2019, 140, 206–223.
- Stal, L.J. Cyanobacteria: Diversity and versatility, clues to life in extreme environments. In Algae and Cyanobacteria in Extreme Environments; Seckbach, J., Ed.; Springer: Dordrecht, The Netherlands, 2007; Volume 11, pp. 661–682.
- Whitton, B.A.; Potts, M. Ecology of Cyanobacteria II: Their Diversity in Space and Time; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2012; pp. 1–13.
- Kurmayer, R.; Deng, L.; Entfellner, E. Role of toxic and bioactive secondary metabolites in colonization and bloom formation by filamentous cyanobacteria Planktothrix. Harmful Algae 2016, 54, 69–86.
- Mazard, S.; Penesyan, A.; Ostrowski, M.; Paulsen, I.T.; Egan, S. Tiny microbes with a big impact: The role of cyanobacteria and their metabolites in shaping our future. Mar. Drugs 2016, 14, 97.
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the key ecological traits of cyanobacteria as a basis for their management and control in changing lakes. Aquat. Ecol. 2016, 50, 333–350.
- Herrero, A.; Flores, E. The Cyanobacteria: Molecular Biology, Genomics and Evolution; Caister Academic Press: Sevilla, Spain, 2008; p. 484.
- Boden, J.S.; Konhauser, K.O.; Robbins, L.J.; Sánchez-Baracaldo, P. Timing the evolution of antioxidant enzymes in cyanobacteria. Nat. Commun. 2021, 12, 4742.
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320.
- Shah, S.A.A.; Akhter, N.; Auckloo, B.N.; Khan, I.; Lu, Y.; Wang, K.; Wu, B.; Guo, Y.-W. Structural diversity, biological properties and applications of natural products from cyanobacteria. A review. Mar. Drugs 2017, 15, 354.
- Carpine, R.; Sieber, S. Antibacterial and antiviral metabolites from cyanobacteria: Their application and their impact on human health. Curr. Res. Biotechnol. 2021, 3, 65–81.
- Prasanna, R.; Nain, L.; Tripathi, R.; Gupta, V.; Chaudhary, V.; Middha, S.; Joshi, M.; Ancha, R.; Kaushik, B.D. Evaluation of fungicidal activity of extracellular filtrates of cyanobacteria—Possible role of hydrolytic enzymes. J. Basic Microbiol. 2008, 48, 186–194.
- Santini, G.; Biondi, N.; Rodolfi, L.; Tredici, M.R. Plant biostimulants from cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643.
- Berthon, J.-Y.; Michel, T.; Wauquier, A.; Joly, P.; Gerbore, J.; Filaire, E. Seaweed and microalgae as major actors of blue biotechnology to achieve plant stimulation and pest and pathogen—A review of the latest advances and future prospects. J. Agric. Sci. 2021, 1, 12.
- Righini, H.; Francioso, O.; Di Foggia, M.; Prodi, A.; Martel Quintana, A.; Roberti, R. Tomato seed biopriming with water extracts from Anabaena minutissima, Ecklonia maxima and Jania adhaerens as a new agro-ecological option against Rhizoctonia solani. Sci. Hortic. 2021, 281, 109921.
- Chaudhary, V.; Prasanna, R.; Nain, L.; Dubey, S.C.; Gupta, V.; Singh, R.; Jaggi, S.; Bhatnagar, A.K. Bioefficacy of novel cyanobacteria-amended formulations in suppressing damping off disease in tomato seedlings. World J. Microbiol. Biotechnol. 2012, 28, 3301–3310.
- Prasanna, R.; Chaudhary, V.; Gupta, V.; Babu, S.; Kumar, A.; Singh, R.; Shivay, Y.S.; Nain, L. Cyanobacteria mediated plant growth promotion and bioprotection against Fusarium wilt in tomato. Eur. J. Plant Pathol. 2013, 136, 337–353.
- Righini, H.; Francioso, O.; Di Foggia, M.; Martel Quintana, A.; Roberti, R. Assessing the potential of the terrestrial cyanobacterium Anabaena minutissima for controlling Botrytis cinerea on tomato fruits. Horticulturae 2021, 7, 210.
- Righini, H.; Somma, A.; Cetrullo, S.; D’Adamo, S.; Flamigni, F.; Martel Quintana, A.; Roberti, R. Inhibitory activity of aqueous extracts from Anabaena minutissima, Ecklonia maxima and Jania adhaerens on the cucumber powdery mildew pathogen in vitro and in vivo. J. Appl. Phycol. 2020, 32, 3363–3375.
- Knogge, W. Fungal infection of plants. Plant Cell 1996, 8, 1711–1722.
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430.
- Gil-Serna, J.; Vázquez, C.; Patiño, B. Genetic regulation of aflatoxin, ochratoxin A, trichothecene, and fumonisin biosynthesis: A review. Int. Microbiol. 2020, 23, 89–96.
- Morales, H.; Marín, S.; Rovira, A.; Ramos, A.J.; Sanchis, V. Patulin accumulation in apples by Penicillium expansum during postharvest stages. Lett. Appl. Microbiol. 2007, 44, 30–35.
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium molds and mycotoxins: Potential species-specific effects. Toxins 2018, 10, 244.
- Carris, L.M.; Little, C.R.; Stiles, C.M. Introduction to Fungi. 2012. Available online: https://www.apsnet.org/edcenter/disandpath/fungalasco/intro/Pages/IntroFungi.aspx (accessed on 15 October 2021).
- Barberis, C.L.; Dalcero, A.M.; Magnoli, C.E. Evaluation of aflatoxin B1 and ochratoxin A in interacting mixed cultures of Aspergillus sections Flavi and Nigri on peanut grains. Mycotoxin Res. 2012, 28, 149–156.
- Gnat, S.; Łagowski, D.; Nowakiewicz, A.; Dyląg, M. A global view on fungal infections in humans and animals: Opportunistic infections and microsporidioses. J. Appl. Microbiol. 2021, 131, 2095–2113.
- Jenkins, S.; Taylor, A.; Jackson, A.C.; Armitage, A.D.; Bates, H.J.; Mead, A.; Harrison, R.J.; Clarkson, J.P. Identification and expression of secreted in xylem pathogenicity genes in Fusarium oxysporum f. sp. pisi. Front. Microbiol. 2021, 12, 593140.
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324.
- Agrios, G. Plant Pathology, 5th ed.; Elsevier Academic Press: San Diego, CA, USA, 2005; p. 952.
- Batista, B.G.; de Chaves, M.A.; Reginatto, P.; Saraiva, O.J.; Fuentefria, A.M. Human fusariosis: An emerging infection that is difficult to treat. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200013.
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant. Pathol. 2012, 13, 329–337.
- Baker, K.F. Types of Rhizoctonia diseases and their occurrence. In Rhizoctonia solani: Biology and Pathology; Parmeter, J.R., Jr., Ed.; University of California Press: Berkeley, CA, USA, 1970; pp. 125–148.
- Fillinger, S.; Elad, Y. Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Springer International Publishing: Cham, Switzerland, 2016; p. 486.
- Pérez-García, A.; Romero, D.; Fernández-Ortuño, D.; López-Ruiz, F.; De Vicente, A.; Tores, J.A. The powdery mildew fungus Podosphaera fusca (synonym Podosphaera xanthii), a constant threat to cucurbits. Mol. Plant Pathol. 2009, 10, 153–160.
- Keinath, A.P.; DuBose, V.B. Evaluation of fungicides for prevention and management of powdery mildew on watermelon. Crop Prot. 2004, 23, 35–42.
- Boddy, L. Pathogens of autotrophs. In The Fungi, 3rd ed.; Watkinson, S.C., Boddy, L., Money, N.P., Eds.; Academic Press: London, UK, 2016; pp. 245–292.
- Marrez, D.A.; Sultan, Y.Y. Antifungal activity of the cyanobacterium Microcystis aeruginosa against mycotoxigenic fungi. J. Appl. Pharm. Sci. 2016, 6, 191–198.
- Shishido, T.K.; Humisto, A.; Jokela, J.; Liu, L.; Wahlsten, M.; Tamrakar, A.; Fewer, D.P.; Permi, P.; Andreote, A.P.D.; Fiore, M.F.; et al. Antifungal compounds from cyanobacteria. Mar. Drugs 2015, 13, 2124–2140.
- El-Sheekh, M.M.; Osman, M.E.H.; Dyab, M.A.; Amer, M.S. Production and characterization of antimicrobial active substance from the cyanobacterium Nostoc muscorum. Environ. Toxicol. Phar. 2006, 21, 42–50.
- Pawar, S.T.; Puranik, P.R. Screening of terrestrial and freshwater halotolerant cyanobacteria for antifungal activities. World J. Microbiol. Biotechnol. 2008, 24, 1019–1025.
- Kim, J.D. Screening of cyanobacteria (blue-green algae) from rice paddy soil for antifungal activity against plant pathogenic fungi. Microbiology 2006, 34, 138–142.
- Biondi, N.; Piccardi, R.; Margheri, M.C.; Rodolfi, L.; Smith, G.D.; Tredici, M.R. Evaluation of Nostoc strain ATCC 53789 as a potential source of natural pesticides. Appl. Environ. Microbiol. 2004, 70, 3313–3320.
- Alwathnani, H.A.; Perveen, K. Biological control of Fusarium wilt of tomato by antagonist fungi and cyanobacteria. Afr. J. Biotechnol. 2012, 11, 1100–1105.
- Kim, J.; Kim, J.-D. Inhibitory effect of algal extracts on mycelial growth of the tomato-wilt pathogen, Fusarium oxysporum f. sp. lycopersici. Mycobiology 2008, 36, 242–248.
- Gupta, V.; Prasanna, R.; Natarajan, C.; Srivastava, A.K.; Sharma, J. Identification, characterization, and regulation of a novel antifungal chitosanase gene (cho) in Anabaena spp. Appl. Environ. Microbiol. 2010, 76, 2769–2777.
- Osman, M.E.A.H.; El-Sheekh, M.M.; Metwally, M.A.; Ismail, A.E.W.A.; Ismail, M.M. Antagonistic activity of some fungi and cyanobacteria species against Rhizoctonia solani. Int. J. Plant Pathol. 2011, 2, 101–114.
- Righini, H.; Francioso, O.; Di Foggia, M.; Martel Quintana, A.; Roberti, R. Preliminary study on the activity of phycobiliproteins against Botrytis cinerea. Mar. Drugs 2020, 18, 600.
- Moreno, A.B.; Martínez del Pozo, Á.; Borja, M.; San Segundo, B. Activity of the antifungal protein from Aspergillus giganteus against Botrytis cinerea. Phytopathology 2003, 93, 1344–1353.
- Righini, H.; Baraldi, E.; García Fernández, Y.; Martel Quintana, A.; Roberti, R. Different Antifungal Activity of Anabaena sp., Ecklonia sp., and Jania sp. against Botrytis cinerea. Mar. Drugs 2019, 17, 299.
- Roberti, R.; Galletti, S.; Burzi, P.L.; Righini, H.; Cetrullo, S.; Perez, C. Induction of defence responses in zucchini (Cucurbita pepo) by Anabaena sp. water extract. Biol. Control 2015, 82, 61–68.
- Kumar, M.; Prasanna, R.; Bidyarani, N.; Babu, S.; Mishra, B.K.; Kumar, A.; Adak, A.; Jauhari, S.; Yadav, K.; Singh, R.; et al. Evaluating the plant growth promoting ability of thermotolerant bacteria and cyanobacteria and their interactions with seed spice crops. Sci. Hortic. 2013, 164, 94–101.
- Belton, S.; McCabe, P.F.; Ng, C.K.Y. The cyanobacterium, Nostoc punctiforme can protect against programmed cell death and induce defence genes in Arabidopsis thaliana. J. Plant Interact. 2021, 16, 64–74.
- Zheng, J.; Zhang, Z.; Tong, T.; Fang, Y.; Zhang, X.; Niu, C.; Li, J.; Wu, Y.; Xue, D.; Zhang, X. Genome-wide identification of wrky gene family and expression analysis under abiotic stress in barley. Agronomy 2021, 11, 521.
- Thapa, S.; Prasanna, R.; Ramakrishnan, B.; Mahawar, H.; Bharti, A.; Kumar, A.; Velmourougane, K.; Shivay, Y.S.; Kumar, A. Microbial inoculation elicited changes in phyllosphere microbial communities and host immunity suppress Magnaporthe oryzae in a susceptible rice cultivar. Physiol. Mol. Plant Pathol. 2021, 114, 101625.
- Priya, H.; Prasanna, R.; Ramakrishnan, B.; Bidyarani, N.; Babu, S.; Thapa, S.; Renuka, N. Influence of cyanobacterial inoculation on the culturable microbiome and growth of rice. Microbiol. Res. 2015, 171, 78–89.
- Le Mire, G.; Siah, A.; Marolleau, B.; Gaucher, M.; Maumené, C.; Brostaux, Y.; Massart, S.; Brisset, M.N.; Jijakli, M.H. Evaluation of λ-carrageenan, CpG-ODN, glycine betaine, Spirulina platensis, and ergosterol as elicitors for control of Zymoseptoria tritici in wheat. Phytopathology 2019, 109, 409–417.
- Rachidi, F.; Benhima, R.; Kasmi, Y.; Sbabou, L.; El Arroussi, H. Evaluation of microalgae polysaccharides as biostimulants of tomato plant defense using metabolomics and biochemical approaches. Sci. Rep. 2021, 11, 930.
- Kapoore, R.V.; Wood, E.E.; Llewellyn, C.A. Algae biostimulants: A critical look at microalgal biostimulants for sustainable agricultural practices. Biotechnol. Adv. 2021, 49, 107754.
- Chiaiese, P.; Corrado, G.; Colla, G.; Kyriacou, M.C.; Rouphael, Y. Renewable sources of plant biostimulation: Microalgae as a sustainable means to improve crop performance. Front. Plant Sci. 2018, 9, 1782.
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587.
- Li, J.; Li, C.; Smith, S.M. Hormone Metabolism and Signaling in Plants; Academic Press Elsevier: London, UK, 2017; p. 597.
- Karthikeyan, N.; Prasanna, R.; Sood, A.; Jaiswal, P.; Nayak, S.; Kaushik, B.D. Physiological characterization and electron microscopic investigation of cyanobacteria associated with wheat rhizosphere. Folia Microbiol. 2009, 54, 43–51.
- Maqubela, M.P.; Mnkeni, P.N.S.; Malam Issa, O.; Pardo, M.T.; D’Acqui, L.P. Nostoc cyanobacterial inoculation in South African agricultural soils enhances soil structure, fertility, and maize growth. Plant Soil 2009, 315, 79–92.
- Řezanka, T.; Palyzová, A.; Sigler, K. Isolation and identification of siderophores produced by cyanobacteria. Folia Microbiol. 2018, 63, 569–579.
- Shukia, S.P.; Singh, J.S.; Kashyap, S.; Giri, D.D.; Kashyap, A.K. Antarctic cyanobacteria as a source of phycocyanin: An assessment. Indian J. Mar. Sci. 2008, 37, 446–449.
- Godlewska, K.; Michalak, I.; Pacyga, P.; Baśladyńska, S.; Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis filtrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 80.
- Markets and Markets. Available online: https://www.marketsandmarkets.com/Market-Reports/biostimulant-market-1081.html (accessed on 9 October 2021).
- Arnau, L. Techno-Economic Feasibility Study for the Production of Microalgae Based Plant Biostimulant. Master’s Thesis, KTH, Royal Institute of Technology School of Chemical Science and Engineering, Stockholm, Sweden, 2016.
- El Boukhari, M.E.M.E.; Barakate, M.; Bouhia, Y.; Lyamlouli, K. Trends in seaweed extract based biostimulants: Manufacturing process and beneficial effect on soil-plant systems. Plants 2020, 9, 359.
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, S.P.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Yunus Khan, T.M.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae biomass as a sustainable source for biofuel, biochemical and biobased value-added products: An integrated biorefinery concept. Fuel 2022, 307, 121782.
- Romero Villegas, G.I.; Fiamengo, M.; Acién Fernández, F.G.; Molina Grima, E. Outdoor production of microalgae biomass at pilot-scale in seawater using centrate as the nutrient source. Algal Res. 2017, 25, 538–548.
- Aghofack-Nguemezi, J.; Schinzoumka, P.A.; Tatchago, V. Effects of extracts or powder of Jatropha curcas and Spirulina platensis on the growth and development of tomato plant. J. Appl. Biosci. 2015, 90, 8413–8420.
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as multi-functional options in modern agriculture: Current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273.
- Mógor, Á.F.; Ördög, V.; Pereira Lima, G.P.; Molnár, Z.; Mógor, G. Biostimulant properties of cyanobacterial hydrolysate related to polyamines. J. Appl. Phycol. 2018, 30, 453–460.
- Supraja, K.V.; Behera, B.; Balasubramanian, P. Efficacy of microalgal extracts as biostimulants through seed treatment and foliar spray for tomato cultivation. Ind. Crops Prod. 2020, 151, 112453.
- Toribio, A.J.; Suárez-Estrella, F.; Jurado, M.M.; López, M.J.; López-González, J.A.; Moreno, J. Prospection of cyanobacteria producing bioactive substances and their application as potential phytostimulating agents. Biotechnol. Rep. 2020, 26, e00449.
- Singh, S. A review on possible elicitor molecules of cyanobacteria: Their role in improving plant growth and providing tolerance against biotic or abiotic stress. J. Appl. Microbiol. 2014, 117, 1221–1244.
- Žižková, E.; Kubeš, M.; Dobrev, P.I.; Přibyl, P.; Šimura, J.; Zahajská, L.; Drábková, L.Z.; Novák, O.; Motyka, V. Control of cytokinin and auxin homeostasis in cyanobacteria and algae. Ann. Bot. 2017, 119, 151–166.
- Bareke, T. Biology of seed development and germination physiology. Adv. Plants Agric. Res. 2018, 8, 336–346.
- Barone, V.; Baglieri, A.; Stevanato, P.; Broccanello, C.; Bertoldo, G.; Bertaggia, M.; Cagnin, M.; Pizzeghello, D.; Moliterni, V.M.C.; Mandolino, G.; et al. Root morphological and molecular responses induced by microalgae extracts in sugar beet (Beta vulgaris L.). J. Appl. Phycol. 2017, 30, 1061–1071.
More