IgG antibodies can cross and do cross the placenta during pregnancy from the mother to the fetus. The passage starts by the end of the first trimester and continues throughout pregnancy. This is a manner by which the fetus gets passive humoral immunity from the mother. In the early neonatal period, these transferred maternal antibodies will act as a defense and help the developing immunity of the baby. While they are short-lived and cleared from the neonatal circulation within several months, they may be very helpful.
A down-side is that in certain maternal autoimmune conditions auto-antibodies seem to have the capacity to hijack the transfer mechanism. It is the case for anti-Ro antibodies. These antibodies are frequent in women with systemic lupus erythematosus and/or Sjogren syndrome. They can lead to fetal atrioventricular block.
- anti-Ro
- fetal heart block
- covid-19
- passive immunity
1. Introduction
2. Physiological Transfer of IgG
2.1. Role of FcRn
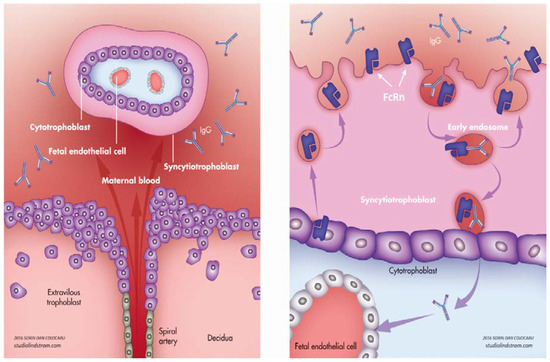
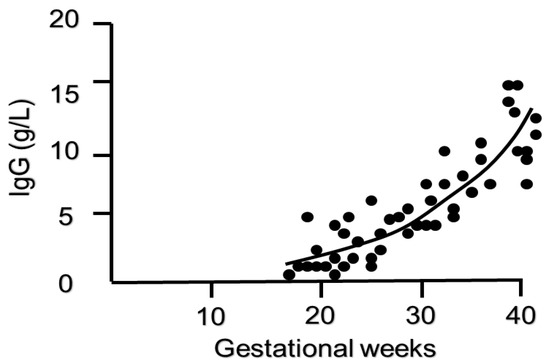
2.2. Timing of IgG Transfer and Other Influencing Factors
2.3. Maternal Immunization
2.3.1. Vaccination
2.3.2. Maternal Infection with COVID-19
3. Pathological Transfer of IgG—Autoimmune Disorders
Transplacental immunoglobulin transfer plays an important role in protecting the newborn in the first few months of life, when the immune system is immature and active immunization through vaccination is not yet recommended. Despite this clear benefit, there are situations in which transplacental transfer may be damaging to the fetus.4. Pathological Transfer of IgG—Alloimmune Disorders
Around 50% of fetal antigens have a paternal origin, and maternal immunization to different fetal antigens inherited from the father, unrecognized as ″self″, determines an immune response with production of antibodies and placental transfer into fetal circulation leading to destruction of different fetal cells, mainly blood cells.5. Pathological Transfer of IgG—Biological Therapy in Pregnancy
6. Conclusions
References
- Chaouat, G.; Petitbarat, M.; Dubanchet, S.; Rahmati, M.; Ledee, N. Tolerance to the Foetal Allograft? Am. J. Reprod. Immunol. 2010, 63, 624–636.
- Lannaman, K.; Romero, R.; Chaiworapongsa, T.; Kim, Y.M.; Korzeniewski, S.J.; Maymon, E.; Gomez-Lopez, N.; Panaitescu, B.; Hassan, S.S.; Yeo, L.; et al. Fetal death: An extreme manifestation of maternal anti-fetal rejection. J. Perinat. Med. 2017, 45, 851–868.
- Wang, Y.; Zhao, S. Vascular biology of the placenta. In Integrated Systems Physiology: From Molecules to Function to Disease; Morgan & Claypool Life Sciences Copyright (c) 2010; Morgan & Claypool Life Sciences: San Rafael, CA, USA, 2010.
- Vidarsson, G.; Dekkers, G.; Rispens, T. IgG Subclasses and Allotypes: From Structure to Effector Functions. Front. Immunol. 2014, 5, 520.
- Hashira, S.; Okitsu-Negishi, S.; Yoshino, K. Placental transfer of IgG subclasses in a Japanese population. Pediatr. Int. 2000, 42, 337–342.
- Challa, D.K.; Velmurugan, R.; Ober, R.J.; Ward, E.S. FcRn: From Molecular Interactions to Regulation of IgG Pharmacokinetics and Functions. Curr. Top. Microbiol. Immunol. 2014, 382, 249–272.
- Palmeira, P.; Quinello, C.; Silveira-Lessa, A.L.; Zago, C.A.; Carneiro-Sampaio, M. IgG Placental Transfer in Healthy and Pathological Pregnancies. Clin. Dev. Immunol. 2012, 2012, 985646.
- Simister, N.E.; Mostov, K.E. An Fc receptor structurally related to MHC class I antigens. Nature 1989, 337, 184–187.
- Vidarsson, G.; Stemerding, A.M.; Stapleton, N.M.; Spliethoff, S.E.; Janssen, H.; Rebers, F.E.; De Haas, M.; Van De Winkel, J.G. FcRn: An IgG receptor on phagocytes with a novel role in phagocytosis. Blood 2006, 108, 3573–3579.
- Akilesh, S.; Christianson, G.J.; Roopenian, D.C.; Shaw, A.S. Neonatal FcR expression in bone marrow-derived cells functions to protect serum IgG from catabolism. J. Immunol. 2007, 179, 4580–4588.
- Pyzik, M.; Sand, K.M.K. Hubbard JJ The Neonatal Fc Receptor (FcRn): A Misnomer? Front. Immunol. 2019, 10, 1540.
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725.
- Leach, J.L.; Sedmak, D.D.; Osborne, J.M.; Rahill, B.; Lairmore, M.D.; Anderson, C.L. Isolation from human placenta of the IgG transporter, FcRn, and localization to the syncytiotrophoblast: Implications for maternal-fetal antibody transport. J. Immunol. 1996, 157, 3317–3322.
- Kristoffersen, E.K.; Matre, R. Co-localization of the neonatal Fc gamma receptor and IgG in human placental term syncytiotrophoblasts. Eur. J. Immunol. 1996, 26, 1668–1671.
- Kiskova, T.; Mytsko, Y.; Schepelmann, M.W.; Helmer, H.; Fuchs, R.; Miedl, H.; Wadsack, C.; Ellinger, I.; Heidi, M. Expression of the neonatal Fc-receptor in placental-fetal endothelium and in cells of the placental immune system. Placenta 2019, 78, 36–43.
- Simister, N.E. Human placental Fc receptors and the trapping of immune complexes. Vaccine 1998, 16, 1451–1455.
- Simister, N.E. Placental transport of immunoglobulin G. Vaccine 2003, 21, 3365–3369.
- Chaudhury, C.; Mehnaz, S.; Robinson, J.M.; Hayton, W.L.; Pearl, D.K.; Roopenian, D.C.; Anderson, C.L. The Major Histocompatibility Complex–related Fc Receptor for IgG (FcRn) Binds Albumin and Prolongs Its Lifespan. J. Exp. Med. 2003, 197, 315–322.
- Junghans, R.P.; Anderson, C.L. The protection receptor for IgG catabolism is the β2-microglobulin containing neonatal intestinal transport receptor. Proc. Natl. Acad. Sci. USA 1996, 93, 5512–5516.
- Shields, R.L.; Namenuk, A.K.; Hong, K.; Meng, Y.G.; Rae, J.; Briggs, J.; Xie, D.; Lai, J.; Stadlen, A.; Li, B.; et al. High Resolution Mapping of the Binding Site on Human IgG1 for FcγRI, FcγRII, FcγRIII, and FcRn and Design of IgG1 Variants with Improved Binding to the FcγR. J. Biol. Chem. 2000, 276, 6591–6604.
- Aloulou, M.; Mkaddem, S.B.; Biarnes-Pelicot, M.; Boussetta, T.; Souchet, H.; Rossato, E.; Benhamou, M.; Crestani, B.; Zhu, Z.; Blank, U.; et al. IgG1 and IVIg induce inhibitory ITAM signaling through FcγRIII controlling inflammatory responses. Blood 2012, 119, 3084–3096.
- Gable, K.L.; Guptill, J.T. Antagonism of the Neonatal Fc Receptor as an Emerging Treatment for Myasthenia Gravis. Front. Immunol. 2020, 10, 3052.
- Jauniaux, E.; Jurkovic, D.; Liesnard, C.; Lees, C.; Campbell, S.; Gulbis, B. Materno-fetal immunoglobulin transfer and passive immunity during the first trimester of human pregnancy. Hum. Reprod. 1995, 10, 3297–3300.
- Malek, A.; Sager, R.; Kuhn, P.; Nicolaides, K.H.; Schneider, H. Evolution of Maternofetal Transport of Immunoglobulins During Human Pregnancy. Am. J. Reprod. Immunol. 1996, 36, 248–255.
- Lessa, A.L.S.; Brasil, T.B.; Pontes, G.N.; Carneiro-Sampaio, M.; Palmeira, P.; Krebs, V.L.J. Preterm and term neonates transplacentally acquire IgG antibodies specific to LPS from Klebsiella pneumoniae, Escherichia coli and Pseudomonas aeruginosa. FEMS Immunol. Med. Microbiol. 2011, 62, 236–243.
- Berg, J.P.V.D.; Westerbeek, E.A.M.; Berbers, G.A.M.; Van Gageldonk, P.; Van Der Klis, F.R.M.; Van Elburg, R.M. Transplacental Transport of IgG Antibodies Specific for Pertussis, Diphtheria, Tetanus, Haemophilus influenzae Type b, and Neisseria meningitidis Serogroup C Is Lower in Preterm Compared with Term Infants. Pediatr. Infect. Dis. J. 2010, 29, 801–805.
- Lozano, N.A.; Lozano, A.; Marini, V.; Saranz, R.J.; Blumberg, R.S.; Baker, K.; Agresta, M.F.; Ponzio, M. Expression of FcRn receptor in placental tissue and its relationship with IgG levels in term and preterm newborns. Am. J. Reprod. Immunol. 2018, 80, e12972.
- Berg, J.V.D.; Westerbeek, E.; Van Der Klis, F.; Berbers, G.; Van Elburg, R. Transplacental transport of IgG antibodies to preterm infants: A review of the literature. Early Hum. Dev. 2011, 87, 67–72.
- Ohlsson, A.; Lacy, J.B. Intravenous immunoglobulin for preventing infection in preterm and/or low birth weight infants. Cochrane Database Syst. Rev. 2020, 1.
- Malek, A.; Sager, R.; Schneider, H. Maternal-Fetal Transport of Immunoglobulin G and Its Subclasses During the Third Trimester of Human Pregnancy. Am. J. Reprod. Immunol. 1994, 32, 8–14.
- Jennewein, M.F.; Goldfarb, I.; Dolatshahi, S.; Cosgrove, C.; Noelette, F.J.; Krykbaeva, M.; Das, J.; Sarkar, A.; Gorman, M.J.; Fischinger, S.; et al. Fc Glycan-Mediated Regulation of Placental Antibody Transfer. Cell 2019, 178, 202–215.
- Hartter, H.K.; Oyedele, O.I.; Dietz, K.; Kreis, S.; Hoffman, J.P.; Muller, C.P. Placental transfer and decay of maternally acquired antimeasles antibodies in Nigerian children. Pediatr. Infect. Dis. J. 2000, 19, 635–641.
- Jones, C.; Naidoo, S.; De Beer, C.; Esser, M.; Kampmann, B.; Hesseling, A.C. Maternal HIV Infection and Antibody Responses Against Vaccine-Preventable Diseases in Uninfected Infants. JAMA 2011, 305, 576–584.
- De Moraes-Pinto, M.I.; Verhoeff, F.; Chimsuku, L.; Milligan, P.; Wesumperuma, L.; Broadhead, R.L.; Brabin, B.J.; Johnson, P.M.; Hart, C.A. Placental antibody transfer: Influence of maternal HIV infection and placental malaria. Arch. Dis. Child. Fetal Neonatal Ed. 1998, 79, F202–F205.
- Palmeira, P.; Costa-Carvalho, B.T.; Arslanian, C.; Pontes, G.N.; Nagao, A.T.; Carneiro-Sampaio, M.M.S. Transfer of antibodies across the placenta and in breast milk from mothers on intravenous immunoglobulin. Pediatr. Allergy Immunol. 2009, 20, 528–535.
- Omer, S.B.; Jamieson, D.J. Maternal Immunization. Plotkin’s Vaccines 2018, 133, 567–578.
- Sperling, R.S.; Riley, L.E. Immunization and Emerging Infections Expert Work Group. Influenza vaccination, pregnancy safety, and risk of early pregnancy loss. Obstet. Gynecol. 2018, 131, 799–802.
- Omer, S.B.; Clark, D.R.; Aqil, A.R.; Tapia, M.D.; Nunes, M.C.; Kozuki, N.; Steinhoff, M.C.; Madhi, S.A.; Wairagkar, N. BMGF Supported Maternal Influenza Immunization Trials Investigators Group Maternal Influenza Immunization and Prevention of Severe Clinical Pneumonia in Young Infants. Pediatr. Infect. Dis. J. 2018, 37, 436–440.
- Jones, C.; Pollock, L.; Barnett, S.M.; Battersby, A.; Kampmann, B. Specific antibodies against vaccine-preventable infections: A mother–infant cohort study. BMJ Open 2013, 3, e002473.
- Centers for Disease Control and Prevention. Updated recommendations for use of tetanus toxoid, reduced diphtheria toxoid, and acellular pertussis vaccine (Tdap) in pregnant women—Advisory committee on immunization practices (ACIP). Morb. Mortal. Wkly. Rep. 2013, 62, 131–135.
- Raya, B.A.; Srugo, I.; Kessel, A.; Peterman, M.; Bader, D.; Gonen, R.; Bamberger, E. The effect of timing of maternal tetanus, diphtheria, and acellular pertussis (Tdap) immunization during pregnancy on newborn pertussis antibody levels—A prospective study. Vaccine 2014, 32, 5787–5793.
- Hillier, S.L.; Ferrieri, P.; Edwards, M.S.; Ewell, M.; Ferris, D.; Fine, P.; Carey, V.; Meyn, L.; Hoagland, D.; Kasper, D.L.; et al. A Phase 2, Randomized, Control Trial of Group B Streptococcus (GBS) Type III Capsular Polysaccharide-tetanus Toxoid (GBS III-TT) Vaccine to Prevent Vaginal Colonization with GBS III. Clin. Infect. Dis. 2018, 68, 2079–2086.
- Muñoz, F.M.; Bond, N.H.; Maccato, M.; Pinell, P.; Hammill, H.A.; Swamy, G.K.; Walter, E.B.; Jackson, L.A.; Englund, J.A.; Edwards, M.S.; et al. Safety and Immunogenicity of Tetanus Diphtheria and Acellular Pertussis (Tdap) Immunization During Pregnancy in Mothers and Infants. JAMA 2014, 311, 1760–1769.
- Siegrist, C.A. Mechanisms by which maternal antibodies influence infant vaccine responses: Review of hypotheses and definition of main determinants. Vaccine 2003, 21, 3406–3412.
- Gans, H. Measles and mumps vaccination as a model to investigate the developing immune system: Passive and active immunity during the first year of life. Vaccine 2003, 21, 3398–3405.
- Lochlainn, L.M.N.; De Gier, B.; Van Der Maas, N.; Strebel, P.M.; Goodman, T.; Van Binnendijk, R.S.; De Melker, H.E.; Hahné, S.J.M. Immunogenicity, effectiveness, and safety of measles vaccination in infants younger than 9 months: A systematic review and meta-analysis. Lancet Infect. Dis. 2019, 19, 1235–1245.
- Sauerbrei, A.; Wutzler, P. Placental boost to varicella-zoster antibodies in the newborn. J. Perinat. Med. 2002, 30, 345–348.
- Leineweber, B.; Grote, V.; Schaad, B.; Heininger, U. Transplacentally acquired immunoglobulin G antibodies against measles, mumps, rubella and varicella-zoster virus in preterm and full term newborns. Pediatr. Infect. Dis. J. 2004, 23, 361–363.
- Plans-Rubió, P.; De Ory, F.; Campins, M.; Álvarez, E.; Payà, T.; Guisasola, E.; Compte, C.; Vellbé, K.; Sanchez, C.; Lozano, M.J.; et al. Prevalence of anti-rubella, anti-measles and anti-mumps IgG antibodies in neonates and pregnant women in Catalonia (Spain) in 2013: Susceptibility to measles increased from 2003 to 2013. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1161–1171.
- McLean, H.Q.; Fiebelkorn, A.P.; Temte, J.L.; Wallace, G.S. Prevention of measles, rubella, congenital rubella syndrome, and mumps, 2013: Summary recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2013, 62, 1–34.
- Van Savage, J.; Decker, M.D.; Edwards, K.M.; Sell, S.H.; Karzon, D.T. Natural History of Pertussis Antibody in the Infant and Effect on Vaccine Response. J. Infect. Dis. 1990, 161, 487–492.
- Novavax. Novavax Announces Topline Results from Phase 3 PrepareTM Trial of ResVaxTM for Prevention of RSV Disease in Infants via Maternal Immunization; Novavax: Gaithersburg, MD, USA, 2019.
- Madhi, S.A.; Cutland, C.; Jose, L.; Koen, A.; Govender, N.; Wittke, F.; Olugbosi, M.; Meulen, A.S.-T.; Baker, S.; Dull, P.M.; et al. Safety and immunogenicity of an investigational maternal trivalent group B streptococcus vaccine in healthy women and their infants: A randomised phase 1b/2 trial. Lancet Infect. Dis. 2016, 16, 923–934.
- Heyderman, R.S.; Madhi, S.A.; French, N.; Cutland, C.L.; Ngwira, B.; Kayambo, D.; Mboizi, R.; Koen, A.; Jose, L.; Olugbosi, M.; et al. Group B streptococcus vaccination in pregnant women with or without HIV in Africa: A non-randomised phase 2, open-label, multicentre trial. Lancet Infect. Dis. 2016, 16, 546–555.
- Barrett, A.D. Current status of Zika vaccine development: Zika vaccines advance into clinical evaluation. NPJ Vaccines 2018, 3, 24.
- Schwartz, D.A. Being Pregnant during the Kivu Ebola Virus Outbreak in DR Congo: The rVSV-ZEBOV Vaccine and Its Accessibility by Mothers and Infants during Humanitarian Crises and in Conflict Areas. Vaccines 2020, 8, 38.
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020.
- Kimberlin, D.W.; Stagno, S. Can SARS-CoV-2 Infection Be Acquired in Utero? More Definitive Evidence Is Needed. JAMA 2020, 18, 1788–1789.
- Zeng, H.; Xu, C.; Fan, J.; Tang, Y.; Deng, Q.; Zhang, W.; Long, X. Antibodies in Infants Born to Mothers with COVID-19 Pneumonia. JAMA 2020, 323.
- Dong, Y.; Chi, X.; Hai, H.; Sun, L.; Zhang, M.; Xie, W.F.; Chen, W.; Huang, H. Antibodies in the breast milk of a maternal woman with COVID-19. Emerg. Microbes Infect. 2020, 9, 1467–1469.
- Davanzo, R.; Moro, G.; Sandri, F.; Agosti, M.; Moretti, C.; Mosca, F. Breastfeeding and coronavirus disease-2019: Ad interim indications of the Italian Society of Neonatology endorsed by the Union of European Neonatal & Perinatal Societies. Matern. Child Nutr. 2020, 16, e13010.
- Donders, F.; Lonnée-Hoffmann, R.; Tsiakalos, A.; Mendling, W.; De Oliveira, J.M.; Judlin, P.; Xue, F.; Donders, G.; Workgroup, G.; Covid, I. ISIDOG Recommendations Concerning COVID-19 and Pregnancy. Diagnostics 2020, 10, 243.
- Borba, V.V.; Zandman-Goddard, G.; Shoenfeld, Y. Exacerbations of autoimmune diseases during pregnancy and postpartum. Best Prac. Res. Clin. Endocrinol. Metab. 2019, 33, 101321.
- Djelmis, J.; Sostarko, M.; Mayer, D.; Ivanisević, M. Myasthenia gravis in pregnancy: Report on 69 cases. Eur. J. Obstet. Gynecol. Reprod. Biol. 2002, 104, 21–25.
- Langer-Gould, A.; Beaber, B.E. Effects of pregnancy and breastfeeding on the multiple sclerosis disease course. Clin. Immunol. 2013, 149, 244–250.
- Brandt, S.V.D.; Zbinden, A.; Baeten, D.; Villiger, P.M.; Østensen, M.; Förger, F. Risk factors for flare and treatment of disease flares during pregnancy in rheumatoid arthritis and axial spondyloarthritis patients. Arthritis Res. Ther. 2017, 19, 64.
- Kwan, L.Y.; Mahadevan, U. Inflammatory bowel disease and pregnancy: An update. Expert Rev. Clin. Immunol. 2010, 6, 643–657.
- Julsgaard, M.; Christensen, L.A.; Gibson, P.R.; Gearry, R.B.; Fallingborg, J.; Hvas, C.L.; Bibby, B.M.; Uldbjerg, N.; Connell, W.; Rosella, O.; et al. Concentrations of Adalimumab and Infliximab in Mothers and Newborns, and Effects on Infection. Gastroenterology 2016, 151, 110–119.
- Chaparro, M.; Verreth, A.; Lobatón, T.; Gravito-Soares, E.; Julsgaard, M.; Savarino, E.V.; Magro, F.; Biron, A.; Lopez-Serrano, P.; Casanova, M.J.; et al. Long-Term Safety of In Utero Exposure to Anti-TNFα Drugs for the Treatment of Inflammatory Bowel Disease: Results from the Multicenter European TEDDY Study. Am. J. Gastroenterol. 2018, 113, 396–403.
- Komaki, F.; Komaki, Y.; Micic, D.; Ido, A.; Sakuraba, A. Outcome of pregnancy and neonatal complications with anti-tumor necrosis factor-α use in females with immune mediated diseases; a systematic review and meta-analysis. J. Autoimmun. 2017, 76, 38–52.
- Bröms, G.; Kieler, H.; Ekbom, A.; Gissler, M.; Hellgren, K.; Lahesmaa-Korpinen, A.-M.; Pedersen, L.; Schmitt-Egenolf, M.; Sørensen, H.; Granath, F. Anti-TNF treatment during pregnancy and birth outcomes: A population-based study from Denmark, Finland, and Sweden. Pharmacoepidemiol. Drug Saf. 2020, 29, 316–327.
- Bröms, G.; Kieler, H.; Ekbom, A.; Gissler, M.; Hellgren, K.; Leinonen, M.K.; Pedersen, L.; Schmitt-Egenolf, M.; Sørensen, H.T.; Granath, F. Paediatric infections in the first 3 years of life after maternal anti-TNF treatment during pregnancy. Aliment. Pharmacol. Ther. 2020.
- Murray, K.E.; Moore, L.; O’Brien, C.; Clohessy, A.; Brophy, C.; Minnock, P.; Fitzgerald, O.; Molloy, E.S.; Mongey, A.-B.; Higgins, S.; et al. Updated pharmacological management of rheumatoid arthritis for women before, during, and after pregnancy, reflecting recent guidelines. Ir. J. Med. Sci. 2018, 188, 169–172.
- Luu, M.; Benzénine, E.; Doret, M.; Michiels, C.; Barkun, A.; Degand, T.; Quantin, C.; Bardou, M. Continuous Anti-TNFα Use Throughout Pregnancy: Possible Complications for the Mother but Not for the Fetus. A Retrospective Cohort on the French National Health Insurance Database (EVASION). Am. J. Gastroenterol. 2018, 113, 1669–1677.
- Soh, M.C.; Moretto, M. The use of biologics for autoimmune rheumatic diseases in fertility and pregnancy. Obstet. Med. 2019, 13, 5–13.
- Kazatchkine, M.D.; Kaveri, S. Immunomodulation of Autoimmune and Inflammatory Diseases with Intravenous Immune Globulin. N. Engl. J. Med. 2001, 345, 747–755.
- Lünemann, J.D.; Quast, I.; Dalakas, M.C. Efficacy of Intravenous Immunoglobulin in Neurological Diseases. Neurotherapeutics 2016, 13, 34–46.
- Li, N.; Zhao, M.; Hilario-Vargas, J.; Prisayanh, P.; Warren, S.; Diaz, L.A.; Roopenian, D.C.; Liu, Z. Complete FcRn dependence for intravenous Ig therapy in autoimmune skin blistering diseases. J. Clin. Investig. 2005, 115, 3440–3450.
- Perez, E.E.; Orange, J.S.; Bonilla, F.; Chinen, J.; Chinn, I.K.; Dorsey, M.; El-Gamal, Y.; Harville, T.O.; Hossny, E.; Mazer, B.; et al. Update on the use of immunoglobulin in human disease: A review of evidence. J. Allergy Clin. Immunol. 2017, 139, S1–S46.
- Lieberman, L.; Greinacher, A.; Murphy, M.F.; Bussel, J.B.; Bakchoul, T.; Corke, S.; Kjaer, M.; Kjeldsen-Kragh, J.; Bertrand, G.; Oepkes, D.; et al. Fetal and neonatal alloimmune thrombocytopenia: Recommendations for evidence-based practice, an international approach. Int. Collab. Transfus. Med. Guidel. (ICTMG) Br. J. Haematol. 2019, 185, 549–562.
- Regan, F.; Lees, C.C.; Jones, B.; Nicolaides, K.H.; Wimalasundera, R.C.; Mijovic, A. Royal College of Obstetricians and Gynaecologists. Prenatal Management of Pregnancies at Risk of Fetal Neonatal Alloimmune Thrombocytopenia (FNAIT): Scientific Impact Paper No. 61. Royal College of Obstetricians and Gynaecologists. BJOG 2019, 126, e173–e185.
- Revello, M.G.; Lazzarotto, T.; Guerra, B.; Spinillo, A.; Ferrazzi, E.; Kustermann, A.; Guaschino, S.; Vergani, P.; Todros, T.; Frusca, T.; et al. A Randomized Trial of Hyperimmune Globulin to Prevent Congenital Cytomegalovirus. N. Engl. J. Med. 2014, 370, 1316–1326.
- Hamprecht, K.; Kagan, K.-O.; Goelz, R.; Van Leeuwen, E.; Rengerink, K.O.; Pajkrt, E.; Spinillo, A.; Gerna, G.; Nigro, G. Hyperimmune Globulin to Prevent Congenital CMV Infection. N. Engl. J. Med. 2014, 370, 2543–2545.