Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Beatrix Zheng and Version 1 by Lu Shi.
Immunotherapy has made great progress in recent years, yet the efficacy of solid tumors remains far less than expected. One of the main hurdles is to overcome the immune-suppressive tumor microenvironment (TME). Among all cells in TME, tumor-associated macrophages (TAMs) play pivotal roles because of their abundance, multifaceted interactions to adaptive and host immune systems, as well as their context-dependent plasticity. Underlying the highly plastic characteristic, lots of research interests are focused on repolarizing TAMs from M2-like pro-tumor phenotype towards M1-like antitumoral ones. Nanotechnology offers great opportunities for targeting and modulating TAM polarization to mount the therapeutic efficacy in cancer immunotherapy.
- macrophage polarization
- tumor-associated macrophages
- nanoparticles
- drug delivery
- tumor microenvironment
- cancer immunotherapy
1. Polymeric Nanoparticles and Macrophage Repolarization
Studies are divided according to the polymer matrix and the agents the nanoparticles carry. Some nanoparticles have the capability to polarize macrophages themselves without reprogramming agents, and the effects are varied for different modifications and residues. Ann-Kathrin Fuchs et al. found that both carboxyl-modified polystyrene nanoparticles and amino-modified ones succeeded in suppressing macrophages from polarizing towards M2 by down regulating the expression of CD200R, CD163, as well as IL-10, without affecting the M1 markers [45][1]. Yen-Jang Huang’s group discovered that hydrophilic polyurethane nanoparticles themselves had surface-dependent immunosuppressive properties, preventing macrophages from M1 polarization by decreasing the production of TNF-α and IL-1β [46][2]. Carboxyl-based nanoparticles were more suppressive than the amino-modified ones [46][2]. The two studies suggested that the effect of nanoparticles on macrophage polarization does not only depend on functional groups but also on other properties, and the nanomedicines should be estimated as a whole. Recently, membrane-coating technology has been used widely in the biomedical field [47[3][4],48], and some cellular membranes could be special agents to influence macrophage polarization. For example, cellular membranes of natural killer cells (NKs) and THP1 macrophages were coated with poly(lactic-co-glycolic acid) (PLGA) nanoparticles, resulting in ten times higher IL-6 than that in the control group [49][5]. Likewise, macrophage membranes were also used to coat PLGA, which delivers iron oxide and TLR agonist R837 to potentiate immunotherapy [50][6]. C.G. Da Silvaa’s group used PLGA as a biodegradable matrix core to simultaneously deliver R848, poly (I:C) and MIP3α, leading to the inhibition of TC-1 growth in pre-clinical experiments [51][7].
TLR agonists are significant in macrophage repolarization. Christopher B. Rodell et al. showed that the TLR7/8 agonist, R848, was one of the most powerful molecules for polarizing macrophages in the M1 direction in vitro among 38 immunomodulatory agents reported in the literature [24][8]. β-cyclodextrin nanoparticles encapsulated with R848 succeeded in regulating TAMs and increased the growth inhibition efficacy of cancer cells in various models together with anti-PD-1 therapy [24][8]. A lignin nanoparticle was also a candidate to carry R848 and targeted CD206-expressing M2 with specific peptides modified on the surface [52][9]. In vitro, the nanomedicine successfully promoted M1 marker TNF-α almost twenty times more than the control and reduced the tumor burden in mice [52][9]. In another group, acetylated chondroitin sulfate protoporphyrin polymer was developed to deliver R837 [53][10]. Together with the other polymeric micelle loading with Dox, it suppressed 4T1 growth in mice [53][10]. The TLR9 agonist CpG has also been carried to modulate macrophages. Jutaek Nam’s lab [54][11] used cationic polyethyleneimine (PEI) to absorb CpG and neoantigen peptides to form a polyplex nano-vaccine. In draining lymph nodes (dLNs) of vaccinated mice, the amount of CD86+ M1 increased, and CD206+ M2 decreased [54][11], showing the capacity of nanovaccines to regulate macrophages. Moreover, TLR agonists and other agents also show great potential for TAM polarization. Plasmid DNA and messenger RNA (mRNA) are two therapeutic agent forms of genetic materials and are also leveraged to edit M1 regulators. For example, a novel lipid-coating polymer termed PQDEA was developed to form polyplexes with IL-12 plasmid and inhibited three tumor models in mice with only four doses [55][12]. Therefore, to further prompt macrophage repolarization, F. Zhang et al. discovered that polymers coated with M1-polarization-associated mRNA and modified with di-mannose could target the CD206 receptor of macrophages, increase M1 macrophages and suppress three tumor models (Figure 1) [56][13]. The M1-associated mRNA coded transcription factors IRF5 and IKKβ, which are downstream proteins in the IFN I pathway, thus promoting M1 skew [56][13]. Yudong Song et al. also modified their polymer with mannose to carry two short interfering RNAs (siRNAs), which block VEGF and PIGF. As these two factors mediate M2 polarization and monocyte recruitments, blocking them resulted in promoting IL-12 and IFN-γ in TME [57][14]. HA and miR125b are effective in promoting M1 polarization. HA-PEI nanoparticles loaded with miR125b were fabricated and succeeded in promoting TAM polarization towards the M1 type after being intraperitoneally injected [58][15]. Another article pointed out that the N-(2-hydroxypropyl) methacrylamide (HPMA)-copolymer nanocarrier could also target CD11b+ TAMs and regulate the TME in situ by inducing M1 polarization [59][16]. Some biophosphonates, such as zoledronic acid (ZOL), were also used to mediate TAM repolarization, and a pH-sensitive dendritic poly-lysine nanoparticle loaded with ZOL could release it once inside the TME [60][17]. For the interference of key chemokines, a shrinkable polymer carrying BLZ-945, a CSF1R inhibitor, to regulate TAM succeeded in reducing CD206+ M2 from 40% to 15%, while IL-12 and IFN-γ in TME increased to three times that of the control [61][18].
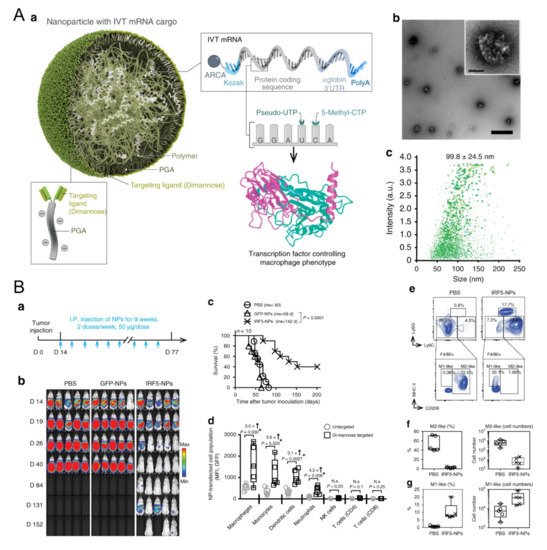
Figure 1. Polymeric nanoparticles loaded with mRNA for macrophage repolarization. (A) Characteristics of the polymeric nanocarriers: (a) The components of the polymeric nanoparticles (termed IRF5-NPs). (b) Transmission electron microscopy of the nanoparticles. (c) Size distribution of the nanocarriers. Reproduced with permission [56][13]. Copyright 2019 Nature Publishing Group. (B) IRF5-NPs prolong the survival time of mice with ovarian cancer and reprogram macrophages in vivo. (a) Treatment schedule. (b) Tumor growth after intraperitoneal administration of IRF5-NPs. (c) The survival curves of tumor-bearing mice. (d) Quantitation of transfection rates in various immune cells using flow cytometry. (e–g) The reprogramming effect of IRF5-NPs on peritoneal macrophages in tumor-bearing mice. Reproduced with permission [56][13]. Copyright 2019 Nature Publishing Group.
2. Lipid-Based Nanomaterials and Macrophage Repolarization
Liposomes are vesicles with a sphere shape and consist of one or more lipid bilayers. They have been in the biomedical research spotlight for several decades because they have special advantages, including: (i) the biomimetic structure where the core of liposomes can encapsulate hydrophilic drugs while the lipid layers can entrap hydrophobic agents. This trait of liposomes might conquer the inconvenient loading of drugs with different dissolubility; (ii) a size of 40–150 nm, exhibiting the same as natural exosomes and thus serving as a biological nanocarrier in therapeutics [65,66][19][20]; (iii) extraordinary biocompatibility to achieve fewer side effects. Of note, liposomes encapsulated with doxorubicin (Doxil®) are the first nanomedicine approved by the FDA. Liposomal cisplatin has also been approved as a drug in pancreatic cancer by the European Agency for the Evaluation of Medicinal Products (EMEA) [67][21]. Moreover, the fast development of microfluidics in recent years facilitates the large-scale manufacture of liposomes and reduces differences from batches to batches [68][22].
Exertions have been made in leveraging liposomal vesicles to carry different agents to polarize TAMs, such as bisphosphonate, siRNA, cytokines, chemokines and TLR agonists. A report proved that ZOL-loaded liposomes succeeded in repolarizing cancer-educated macrophages towards a pro-inflammatory subtype with increasing expression of iNOS and TNF-α [69][23]. Another study showed that the pegylated liposomal nanoparticles (PLNs) loaded with alendronate could suppress tumor growth and increase progression-free survival in tumor-bearing mice. Interestingly, the PLNs themselves, on the contrary, empowered immunosuppression and impaired T-cell immunity [70][24]. As aforementioned, cues in TME polarize TAMs towards M2-like subtypes, and among them, hypoxia-induced factor 1-α (HIF-1α) are notorious. Therefore, a lipid nanocarrier with HIF-1α-blocking siRNA was developed and succeeded in promoting the secretion of TNF-α and IFN-γ in the TME [71][25]. The amount of CD169+ TAMs was also enhanced to 1.5-fold over the control group [71][25], demonstrating that the nanostrategy had regulated TAMs towards an immune-supportive direction. The polarization effect of TLR agonists on TAMs has greatly benefited from nanotechnology. A research group found that R848-encapsulated liposomes together with anti-EGFR antibody treatment could significantly suppress tumor growth in mice. Intriguingly, they found that M1 macrophages induced antibody-dependent cellular phagocytosis more strongly than M2 [72][26]. In addition, cytokine and chemokine therapies, which also benefited from nanocarriers, are candidates for reprogramming TAMs because of their powerful roles in tuning cellular signal pathways and phenotypes. In a recent report, one kind of clinical-approved liposome was used to deliver mRNA, which edited a bispecific antibody to neutralize CCL2 and CCL5 (Figure 2) [73][27]. The nanomedicine down regulated IL-10, ARG1, MRC1 and CD206 and suppressed liver cancer growth in mice together with PD-1 antibody treatment [73][27]. Anujan Ramesh’s lab constructed a liposome to carry BLZ945 and SHP099 (inhibitors of CSF1R and CD47-SIRPα signal pathway, respectively,) and the nanoparticle significantly down regulated the expression of CD206 in a macrophage cell line Raw264.7 from ~75% to ~10%, increasing the M1/M2 ratio by six times [74][28]. Recently, the same group constructed another liposome to deliver BLZ945 and another inhibitor Selumetinib to promote M1 polarization [75][29]. Some drugs available in the clinic might also have positive effects on programming macrophages. For instance, liposomes loaded with simvastatin and vorinostat also had positive impacts in down regulating CD206 and promoting CD86 expression [76][30]. To overcome the off-target effects of regulating compounds, another research group modified sialic acid to their liposomes and loaded with zoledronic acid because of the hig -expression of sialic acid receptors in TAMs [77][31].
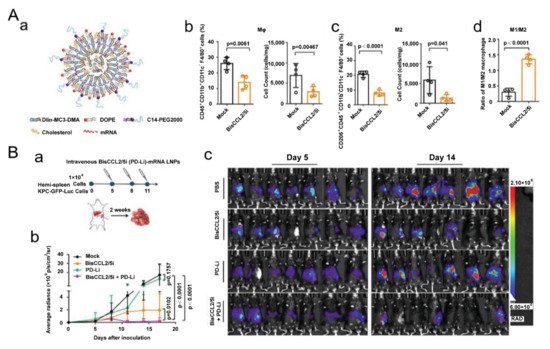
Figure 2. Lipid-based nanoparticles used for mRNA delivery of a bispecific single-domain antibody for repolarizing TAMs. (A) Lipid-based nanoparticles loaded with mRNA to realize CCL2 and CCL5 dual blockade reprogram TAMs. (a) Designs of the lipid-based nanocarriers. (b) The percentage of macrophages and (c) M2 subtypes in tumor tissue 48 h after systemic administration of the nanoparticles. (d) The ratio of M1/M2 in the TME. Reproduced with permission [73][27]. Copyright 2021 Wiley-VCH. (B) Dual blockade nano-strategy combined with PD-1/PD-L1 inhibition suppresses KPC liver cancer growth. (a) Time lines of the experiment. (b) Tumor growth after various treatments. (c) In vivo bioluminescence of tumor-bearing mice on days 5 and 14. Reproduced with permission [73][27]. Copyright 2021 Wiley-VCH.
Lipid nanoemulsions can also be fabricated with microfluidic or with ultrasonic devices. Similar to liposomes, nanoemulsions are sphere-shaped particles consisting of several kinds of lipids in a droplet and usually exhibit a low polydispersity. For the oil-in-water system, lipid emulsion can entrap hydrophobic drug molecules inside the oil droplet. Different TLR agonists might contribute to macrophage polarization to a different degree. A nanoemulsion carried with TLR7/8 agonists, R848, skewed more macrophages towards M1 directions than that loaded with R837 [78][32]. Together with vaccine treatment, this nanomedicine prolonged the survival time of animal models with melanoma and cervical cancer for it might revert “cold” tumors into the “hot” ones [78][32]. Special therapeutic agents have also been discovered to have the ability to convert macrophages. Ye Hui et al. [79][33] constructed a lipid nanoemulsion to carry an isoflavone gained from Psoralea corylifolia L., termed Neobavaisoflavone. This anti-cancer material was able to switch M2 macrophages to pro-inflammatory M1 in vivo [79][33].
3. Inorganic Nanoparticles and Macrophage Repolarization
Inorganic nanomaterials are defined as nanoparticles composed of inanimate maters and usually include a metal matrix, such as calcium, iron and gold, and nonmetal materials, such as silicon. The original properties confer inorganic nanoparticles lots of merits, including: (i) relatively stable for long time period conservation and for strict sterilization conditions compared to organic materials [80][34]; (ii) fine controllability for the structure and various shapes with a low polydispersity index [81][35]; (iii) the intrinsic physical properties for multipurpose applications, such as superparamagnetism, up-conversion luminescence and surface plasmon resonance [81,82][35][36]. In fact, some inorganic nanoparticles have been clinically approved. For instance, iron oxide-based nanomaterials, ferumoxytol (i.e., Feraheme®) and ferucarbotran (i.e., Resovist®) have been used to treat iron deficiency and complete magnetic resonance imaging (MRI), respectively [35,83][37][38]. In order to demonstrate inorganic nanomedicines in macrophage repolarization, we outline the approaches according to their intrinsic properties and the agents as well.
As aforementioned, some nanoparticles have the ability to reprogram macrophages themselves, and iron oxide nanomaterials are one of them. In 2016, Saeid Zanganeh et al. [84][39] found that the clinical-approved iron oxide nanoparticle ferumoxytol could inhibit tumor growth by polarizing macrophages in the M1 direction. Interestingly, it was later discovered that both clinical-approved iron oxide nanoparticles, feracarbotran and ferumoxytol, were able to induce macrophage autophagy and arouse inflammatory response through TLR4-mediated signaling and oxidative stress [85][40]. With the extracts help from targeting molecules and other reprogramming agents to TAMs, these self-service nanomedicines might further promote the efficacy and retardation of tumor development. Jiaojiao Zhao et al. found that ferumoxytol surface-functionalized with poly(I:C) could achieve macrophage activation and suppress malignant melanoma in mice [86][41]. Similarly, other iron oxide nanoparticles have also been reported to be effective in turning macrophages into anti-tumor subtypes [83,87,88,89,90,91][38][42][43][44][45][46]. Coating iron oxide with a cellular membrane expressing SIRPα from genetic-edited cells was proven to be an effective strategy. The iron-containing nanoparticles were magnetically oriented to aggregate in the TME, and with the blockade of the “do not eat me” signal, TAMs were repolarized towards M1, eliciting potent immune responses and suppressing both B16F10 and 4T1 cancer growth with T lymphocytes [92][47]. Super-paramagnetic iron oxide nanoparticles (SPIONs) with different charges were compared in a recent study [93][48]. SPIONs with positive or negative charges could skew macrophages towards M1-like phenotypes with a great promotion in TNF-α production [93][48]. Hollow iron oxide with PI3Kγ inhibitor payload and modified with mannose efficiently promoted NF-kB p65 expression and reprogrammed TAMs to M1 [94][49]. In an animal model with human breast cancer MDA-MB-231, the hollow nanomedicine inhibited tumor development [94][49]. Hyaluronic acid (HA) was carried by various iron oxide nanoparticles to regulate macrophages to promote the polarization effect [95,96,97][50][51][52]. Of note, the clinical observation is in accordance with the story above, where the number of iron-containing TAMs in patients with non-small cell lung cancer is usually associated with tumor regression [98][53]. Researchers further confirmed that the phenomenon resulted from TAM repolarizing into pro-inflammatory type after consuming iron-containing sub heme [98][53].
Furthermore, iron-relative nanoparticles, calcium ions and some catalytic nanoparticles have also been proven to regulate macrophage phenotypes. Calcium iron itself could boost pro-inflammatory cytokines production, such as IL-1β [99][54]. Therefore, nanoparticle medicine using calcium might have positive effects on M1 repolarization. Xiao-Yan He et al. delivered HA and IL-12-coded plasmid DNA with peptide-modified calcium carbonate and succeeded in multiple M1 markers increasing in J774A.1 cells [100][55]. Additionally, a nano-catalytic medicine was developed to target mitochondrial DNA (mtDNA) of cancer cells [101][56]. With the mtDNA oxidative damagecaused by nanoparticles, the remaining parts of nucleic acid escaped from tumor cells and then entered TAMs as damage-associated molecular patterns (DAMPs), which turned macrophages into M1 types [101][56]. IL-1β, TNF-α, IFN-β and IL-18 production reached as high as the positive control (i.e., LPS-treated group) after treatment. In addition, this novel strategy was verified to be effective on a PANC-1 tumor xenograft model [101][56].
Like other materials, inorganic nanoparticles can also be loaded with different reprogramming cargos. Photo-responsive up-conversion nanoparticles loaded with TAM-reprogramming agents achieved photo-related therapy and macrophage polarization simultaneously [102,103][57][58]. Ru nanoparticles loaded with BLZ945 also worked to decrease iNOS and CD206 expressions [104][59]. Non-mental materials, such as mesoporous silicon nanoparticles (MSN), are famous for their porous structure, which contributes to the delivery of more polarization-related agents in vivo and in vitro [101,105][56][60]. Leonard et al. also leveraged MSN to load albumin-paclitaxel, promoting M1 polarization [106][61]. Mesoporous silica loaded with siRNA to block monocarboxylate transporter-4 (MCT-4) also succeeded in preventing TAM from M2 polarization for MCT play a significant role in maintaining the acidic TME (Figure 3) [105][60]. Liming Bian and his colleagues coated MSN with up-conversion materials and loaded it with calcium regulators, which are released under near-infrared rays. Eventually, calcium levels in the cells increase or decrease, resulting in macrophage polarizing towards M1 or M2, respectively [103][58]. Gold nanoparticles linked with mucin-1 peptides succeeded in promoting M1 polarization with increasing cytokines, such as TNF-α, IL-6, IL-10, as well as IL-12 [107][62]. Li et al. rapped albumin and paclitaxel into gold nanorods, which suppressed M2 polarization and modulated the TME in tumor-bearing mice [108][63].
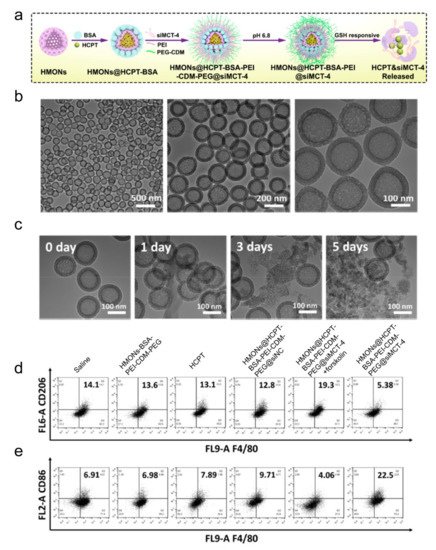
Figure 3. Inorganic nanoparticles with siRNA to block MCT-4 for repolarizing TAMs. (a) Schematic illustration for hollow mesoporous organosilica loaded with hydroxycamptothecin and siRNA-blocking MCT-4. (b) Transmission electron microscopy of the nanomedicines. (c) Transmission electron microscopy of the nanoparticles after incubating with a buffer containing redox glutathione for the number of indicated days. (d,e) Flow cytometric analysis of TAM phenotypes using the (d) M2 marker CD206 and (e) M1 marker CD86. Reproduced with permission [60]. Copyright 2020 American Chemical Society.
References
- Fuchs, A.K.; Syrovets, T.; Haas, K.A.; Loos, C.; Musyanovych, A.; Mailander, V.; Landfester, K.; Simmet, T. Carboxyl- and amino-functionalized polystyrene nanoparticles differentially affect the polarization profile of M1 and M2 macrophage subsets. Biomaterials 2016, 85, 78–87.
- Huang, Y.J.; Hung, K.C.; Hung, H.S.; Hsu, S.H. Modulation of Macrophage Phenotype by Biodegradable Polyurethane Nanoparticles: Possible Relation between Macrophage Polarization and Immune Response of Nanoparticles. Appl. Mater Interfaces 2018, 10, 19436–19448.
- Zhou, J.; Kroll, A.V.; Holay, M.; Fang, R.H.; Zhang, L. Biomimetic Nanotechnology toward Personalized Vaccines. Adv Mater 2019, e1901255.
- Fang, R.H.; Jiang, Y.; Fang, J.C.; Zhang, L. Cell memb.brane-derived nanomaterials for biomedical applications. Biomaterials 2017, 128, 69–83.
- Deng, G.; Sun, Z.; Li, S.; Peng, X.; Li, W.; Zhou, L.; Ma, Y.; Gong, P.; Cai, L. Cell-Membrane Immunotherapy Based on Natural Killer Cell Membrane Coated Nanoparticles for the Effective Inhibition of Primary and Abscopal Tumor Growth. ACS Nano 2018, 12, 12096–12108.
- Liu, L.; Wang, Y.; Guo, X.; Zhao, J.; Zhou, S. A Biomimetic Polymer Magnetic Nanocarrier Polarizing Tumor-Associated Macrophages for Potentiating Immunotherapy. Small 2020, e2003543.
- Da Silva, C.G.; Camps, M.G.M.; Li, T.; Chan, A.B.; Ossendorp, F.; Cruz, L.J. Co-delivery of immunomodulators in biodegradable nanoparticles improves therapeutic efficacy of cancer vaccines. Biomaterials 2019, 220, 119417.
- Rodell, C.B.; Arlauckas, S.P.; Cuccarese, M.F.; Garris, C.S.; Li, R.; Ahmed, M.S.; Kohler, R.H.; Pittet, M.J.; Weissleder, R. TLR7/8-agonist-loaded nanoparticles promote the polarization of tumour-associated macrophages to enhance cancer immunotherapy. Nat. Biomed. Eng. 2018, 2, 578–588.
- Figueiredo, P.; Lepland, A.; Scodeller, P.; Fontana, F.; Torrieri, G.; Tiboni, M.; Shahbazi, M.A.; Casettari, L.; Kostiainen, M.A.; Hirvonen, J.; et al. Peptide-guided resiquimod-loaded lignin nanoparticles convert tumor-associated macrophages from M2 to M1 phenotype for enhanced chemotherapy. Acta Biomater. 2020.
- Wei, X.; Liu, L.; Li, X.; Wang, Y.; Guo, X.; Zhao, J.; Zhou, S. Selectively targeting tumor-associated macrophages and tumor cells with polymeric micelles for enhanced cancer chemo-immunotherapy. J. Control. Release 2019, 313, 42–53.
- Nam, J.; Son, S.; Park, K.S.; Moon, J.J. Modularly Programmable Nanoparticle Vaccine Based on Polyethyleneimine for Personalized Cancer Immunotherapy. Adv. Sci. 2021, 8, 2002577.
- Qiu, N.; Wang, G.; Wang, J.; Zhou, Q.; Guo, M.; Wang, Y.; Hu, X.; Zhou, H.; Bai, R.; You, M.; et al. Tumor-Associated Macrophage and Tumor-Cell Dually Transfecting Polyplexes for Efficient Interleukin-12 Cancer Gene Therapy. Adv. Mater. 2021, 33, e2006189.
- Zhang, F.; Parayath, N.N.; Ene, C.I.; Stephan, S.B.; Koehne, A.L.; Coon, M.E.; Holland, E.C.; Stephan, M.T. Genetic programming of macrophages to perform anti-tumor functions using targeted mRNA nanocarriers. Nat. Commun. 2019, 10, 3974.
- Song, Y.; Tang, C.; Yin, C. Combination antitumor immunotherapy with VEGF and PIGF siRNA via systemic delivery of multi-functionalized nanoparticles to tumor-associated macrophages and breast cancer cells. Biomaterials 2018, 185, 117–132.
- Parayath, N.N.; Parikh, A.; Amiji, M.M. Repolarization of Tumor-Associated Macrophages in a Genetically Engineered Nonsmall Cell Lung Cancer Model by Intraperitoneal Administration of Hyaluronic Acid-Based Nanoparticles Encapsulating MicroRNA-125b. Nano Lett. 2018, 18, 3571–3579.
- Zimel, M.N.; Horowitz, C.B.; Rajasekhar, V.K.; Christ, A.B.; Wei, X.; Wu, J.; Wojnarowicz, P.M.; Wang, D.; Goldring, S.R.; Purdue, P.E.; et al. HPMA-Copolymer Nanocarrier Targets Tumor-Associated Macrophages in Primary and Metastatic Breast Cancer. Mol. Cancer 2017, 16, 2701–2710.
- Guo, Q.; He, X.; Li, C.; He, Y.; Peng, Y.; Zhang, Y.; Lu, Y.; Chen, X.; Zhang, Y.; Chen, Q.; et al. Dandelion-Like Tailorable Nanoparticles for Tumor Microenvironment Modulation. Adv. Sci. 2019, 6, 1901430.
- Wang, J.; Shen, S.; Li, J.; Cao, Z.; Yang, X. Precise Depletion of Tumor Seed and Growing Soil with Shrinkable Nanocarrier for Potentiated Cancer Chemoimmunotherapy. ACS Nano 2021, 15, 4636–4646.
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics 2018, 10, 218.
- Yuba, E. Liposome-based immunity-inducing systems for cancer immunotherapy. Mol. Immunol. 2018, 98, 8–12.
- Boulikas, T. Clinical overview on Lipoplatin: A successful liposomal formulation of cisplatin. Expert Opin. Investig. Drugs 2009, 18, 1197–1218.
- Damiati, S.; Kompella, U.B.; Damiati, S.A.; Kodzius, R. Microfluidic Devices for Drug Delivery Systems and Drug Screening. Genes 2018, 9, 103.
- Sousa, S.; Auriola, S.; Monkkonen, J.; Maatta, J. Liposome encapsulated zoledronate favours M1-like behaviour in murine macrophages cultured with soluble factors from breast cancer cells. BMC Cancer 2015, 15, 4.
- Rajan, R.; Sabnani, M.K.; Mavinkurve, V.; Shmeeda, H.; Mansouri, H.; Bonkoungou, S.; Le, A.D.; Wood, L.M.; Gabizon, A.A.; La-Beck, N.M. Liposome-induced immunosuppression and tumor growth is mediated by macrophages and mitigated by liposome-encapsulated alendronate. J. Control. Release 2018, 271, 139–148.
- Shobaki, N.; Sato, Y.; Suzuki, Y.; Okabe, N.; Harashima, H. Manipulating the function of tumor-associated macrophages by siRNA-loaded lipid nanoparticles for cancer immunotherapy. J. Control. Release 2020, 325, 235–248.
- Li, H.; Somiya, M.; Kuroda, S.i. Enhancing antibody-dependent cellular phagocytosis by Re-education of tumor-associated macrophages with resiquimod-encapsulated liposomes. Biomaterials 2021, 268, 120601.
- Wang, Y.; Tiruthani, K.; Li, S.; Hu, M.; Zhong, G.; Tang, Y.; Roy, S.; Zhang, L.; Tan, J.; Liao, C.; et al. mRNA Delivery of a Bispecific Single-Domain Antibody to Polarize Tumor-Associated Macrophages and Synergize Immunotherapy against Liver Malignancies. Adv. Mater. 2021.
- Ramesh, A.; Kumar, S.; Nandi, D.; Kulkarni, A. CSF1R- and SHP2-Inhibitor-Loaded Nanoparticles Enhance Cytotoxic Activity and Phagocytosis in Tumor-Associated Macrophages. Adv. Mater. 2019, e1904364.
- Ramesh, A.; Brouillard, A.; Kumar, S.; Nandi, D.; Kulkarni, A. Dual inhibition of CSF1R and MAPK pathways using supramolecular nanoparticles enhances macrophage immunotherapy. Biomaterials 2019, 227, 119559.
- Tu, B.; He, Y.; Chen, B.; Wang, Y.; Gao, Y.; Shi, M.; Liu, T.; Asrorov, A.M.; Huang, Y. Deformable liposomal codelivery of vorinostat and simvastatin promotes antitumor responses through remodeling tumor microenvironment. Biomater. Sci. 2020, 8, 7166–7176.
- Tang, X.; Sui, D.; Liu, M.; Zhang, H.; Liu, M.; Wang, S.; Zhao, D.; Sun, W.; Liu, M.; Luo, X.; et al. Targeted delivery of zoledronic acid through the sialic acid—Siglec axis for killing and reversal of M2 phenotypic tumor-associated macrophages—A promising cancer immunotherapy. Int. J. Pharm. 2020, 590, 119929.
- Kim, S.Y.; Kim, S.; Kim, J.E.; Lee, S.N.; Shin, I.W.; Shin, H.S.; Jin, S.M.; Noh, Y.W.; Kang, Y.J.; Kim, Y.S.; et al. Lyophilizable and Multifaceted Toll-like Receptor 7/8 Agonist-Loaded Nanoemulsion for the Reprogramming of Tumor Microenvironments and Enhanced Cancer Immunotherapy. ACS Nano 2019, 13, 12671–12686.
- Ye, H.; He, X.; Feng, X. Developing neobavaisoflavone nanoemulsion suppresses lung cancer progression by regulating tumor microenvironment. Biomed. Pharm. 2020, 129, 110369.
- Das, A.; Ali, N. Nanovaccine: An emerging strategy. Expert Rev. Vaccines 2021, 20, 1273–1290.
- Jiao, M.; Zhang, P.; Meng, J.; Li, Y.; Liu, C.; Luo, X.; Gao, M. Recent advancements in biocompatible inorganic nanoparticles towards biomedical applications. Biomater. Sci. 2018, 6, 726–745.
- Wang, X.; Zhong, X.; Li, J.; Liu, Z.; Cheng, L. Inorganic nanomaterials with rapid clearance for biomedical applications. Chem. Soc. Rev. 2021, 50, 8669–8742.
- Bobo, D.; Robinson, K.J.; Islam, J.; Thurecht, K.J.; Corrie, S.R. Nanoparticle-Based Medicines: A Review of FDA-Approved Materials and Clinical Trials to Date. Pharm. Res. 2016, 33, 2373–2387.
- Shah, A.; Dobrovolskaia, M.A. Immunological effects of iron oxide nanoparticles and iron-based complex drug formulations: Therapeutic benefits, toxicity, mechanistic insights, and translational considerations. Nanomedicine 2018, 14, 977–990.
- Zanganeh, S.; Hutter, G.; Spitler, R.; Lenkov, O.; Mahmoudi, M.; Shaw, A.; Pajarinen, J.S.; Nejadnik, H.; Goodman, S.; Moseley, M.; et al. Iron oxide nanoparticles inhibit tumour growth by inducing pro-inflammatory macrophage polarization in tumour tissues. Nat. Nanotechnol. 2016, 11, 986–994.
- Jin, R.; Liu, L.; Zhu, W.; Li, D.; Yang, L.; Duan, J.; Cai, Z.; Nie, Y.; Zhang, Y.; Gong, Q.; et al. Iron oxide nanoparticles promote macrophage autophagy and inflammatory response through activation of toll-like Receptor-4 signaling. Biomaterials 2019, 203, 23–30.
- Zhao, J.; Zhang, Z.; Xue, Y.; Wang, G.; Cheng, Y.; Pan, Y.; Zhao, S.; Hou, Y. Anti-tumor macrophages activated by ferumoxytol combined or surface-functionalized with the TLR3 agonist poly (I: C) promote melanoma regression. Theranostics 2018, 8, 6307–6321.
- Rojas, J.M.; Sanz-Ortega, L.; Mulens-Arias, V.; Gutierrez, L.; Perez-Yague, S.; Barber, D.F. Superparamagnetic iron oxide nanoparticle uptake alters M2 macrophage phenotype, iron metabolism, migration and invasion. Nanomedicine 2016, 12, 1127–1138.
- Kang, H.; Kim, S.; Wong, D.S.H.; Jung, H.J.; Lin, S.; Zou, K.; Li, R.; Li, G.; Dravid, V.P.; Bian, L. Remote Manipulation of Ligand Nano-Oscillations Regulates Adhesion and Polarization of Macrophages in vivo. Nano Lett. 2017, 17, 6415–6427.
- Guo, J.-C.; An, Q.; Guo, M.; Xiao, Y.; Li, B.; Gao, F.; Wang, Y.; Li, J.; Wang, Y.; Liu, Y.; et al. Oxygen-independent free radical generation mediated by core-shell magnetic nanocomposites synergizes with immune checkpoint blockade for effective primary and metastatic tumor treatment. Nano Today 2021, 36, 101024.
- Yang, Y.; Tian, Q.; Wu, S.; Li, Y.; Yang, K.; Yan, Y.; Shang, L.; Li, A.; Zhang, L. Blue light-triggered Fe2+-release from monodispersed ferrihydrite nanoparticles for cancer iron therapy. Biomaterials 2021, 271.
- Chen, L.; Ma, X.; Dang, M.; Dong, H.; Hu, H.; Su, X.; Liu, W.; Wang, Q.; Mou, Y.; Teng, Z. Simultaneous T Cell Activation and Macrophage Polarization to Promote Potent Tumor Suppression by Iron Oxide-Embedded Large-Pore Mesoporous Organosilica Core-Shell Nanospheres. Adv. Healthc. Mater. 2019, 8, e1900039.
- Rao, L.; Zhao, S.K.; Wen, C.; Tian, R.; Lin, L.; Cai, B.; Sun, Y.; Kang, F.; Yang, Z.; He, L.; et al. Activating Macrophage-Mediated Cancer Immunotherapy by Genetically Edited Nanoparticles. Adv. Mater. 2020, 32, e2004853.
- Zhang, W.; Cao, S.; Liang, S.; Tan, C.H.; Luo, B.; Xu, X.; Saw, P.E. Differently Charged Super-Paramagnetic Iron Oxide Nanoparticles Preferentially Induced M1-Like Phenotype of Macrophages. Front. Bioeng. Biotechnol. 2020, 8, 537.
- Li, K.; Lu, L.; Xue, C.; Liu, J.; He, Y.; Zhou, J.; Xia, Z.; Dai, L.; Luo, Z.; Mao, Y.; et al. Polarization of tumor-associated macrophage phenotype via porous hollow iron nanoparticles for tumor immunotherapy in vivo. Nanoscale 2020, 12, 130–144.
- Gong, T.; Dong, Z.; Fu, Y.; Gong, T.; Deng, L.; Zhang, Z. Hyaluronic acid modified doxorubicin loaded Fe3O4 nanoparticles effectively inhibit breast cancer metastasis. J. Mater. Chem. B 2019, 7, 5861–5872.
- Li, C.X.; Zhang, Y.; Dong, X.; Zhang, L.; Liu, M.D.; Li, B.; Zhang, M.K.; Feng, J.; Zhang, X.Z. Artificially Reprogrammed Macrophages as Tumor-Tropic Immunosuppression-Resistant Biologics to Realize Therapeutics Production and Immune Activation. Adv. Mater. 2019, 31, e1807211.
- Sun, W.; Yang, J.; Hou, M.; Xie, S.; Xiong, L.; Li, B.; Zhang, C. A Nano “Immune-Guide” Recruiting Lymphocytes and Modulating the Ratio of Macrophages from Different Origins to Enhance Cancer Immunotherapy. Adv. Funct. Mater. 2021.
- Costa da Silva, M.; Breckwoldt, M.O.; Vinchi, F.; Correia, M.P.; Stojanovic, A.; Thielmann, C.M.; Meister, M.; Muley, T.; Warth, A.; Platten, M.; et al. Iron Induces Anti-tumor Activity in Tumor-Associated Macrophages. Front. Immunol. 2017, 8, 01479.
- Pazar, B.; Ea, H.K.; Narayan, S.; Kolly, L.; Bagnoud, N.; Chobaz, V.; Roger, T.; Liote, F.; So, A.; Busso, N. Basic calcium phosphate crystals induce monocyte/macrophage IL-1beta secretion through the NLRP3 inflammasome in vitro. J. Immunol. 2011, 186, 2495–2502.
- He, X.Y.; Liu, B.Y.; Xu, C.; Zhuo, R.X.; Cheng, S.X. A multi-functional macrophage and tumor targeting gene delivery system for the regulation of macrophage polarity and reversal of cancer immunoresistance. Nanoscale 2018, 10, 15578–15587.
- Jiang, H.; Guo, Y.; Wei, C.; Hu, P.; Shi, J. Nanocatalytic Innate Immunity Activation by Mitochondrial DNA Oxidative Damage for Tumor-Specific Therapy. Adv. Mater. 2021, e2008065.
- Ai, X.; Hu, M.; Wang, Z.; Lyu, L.; Zhang, W.; Li, J.; Yang, H.; Lin, J.; Xing, B. Enhanced Cellular Ablation by Attenuating Hypoxia Status and Reprogramming Tumor-Associated Macrophages via NIR Light-Responsive Upconversion Nanocrystals. Bioconjug. Chem. 2018, 29, 928–938.
- Kang, H.; Zhang, K.; Wong, D.S.H.; Han, F.; Li, B.; Bian, L. Near-infrared light-controlled regulation of intracellular calcium to modulate macrophage polarization. Biomaterials 2018, 178, 681–696.
- Liu, Y.; Wen, Y.; Chen, X.; Zhu, X.; Yu, Q.; Gong, Y.; Yuan, G.; Liu, J.; Qin, X. Inflammation-responsive functional Ru nanoparticles combining a tumor-associated macrophage repolarization strategy with phototherapy for colorectal cancer therapy. J. Mater. Chem. B 2019, 7, 6210–6223.
- Li, K.; Lin, C.; He, Y.; Lu, L.; Xu, K.; Tao, B.; Xia, Z.; Zeng, R.; Mao, Y.; Luo, Z.; et al. Engineering of Cascade-Responsive Nanoplatform to Inhibit Lactate Efflux for Enhanced Tumor Chemo-Immunotherapy. ACS Nano 2020, 14, 14164–14180.
- Leonard, F.; Curtis, L.T.; Ware, M.J.; Nosrat, T.; Liu, X.; Yokoi, K.; Frieboes, H.B.; Godin, B. Macrophage Polarization Contributes to the Anti-Tumoral Efficacy of Mesoporous Nanovectors Loaded with Albumin-Bound Paclitaxel. Front. Immunol. 2017, 8, 693.
- Mocan, T.; Matea, C.; Tabaran, F.; Iancu, C.; Orasan, R.; Mocan, L. In Vitro Administration of Gold Nanoparticles Functionalized with MUC-1 Protein Fragment Generates Anticancer Vaccine Response via Macrophage Activation and Polarization Mechanism. J. Cancer 2015, 6, 583–592.
- Li, D.; Zhang, M.; Xu, F.; Chen, Y.; Chen, B.; Chang, Y.; Zhong, H.; Jin, H.; Huang, Y. Biomimetic albumin-modified gold nanorods for photothermo-chemotherapy and macrophage polarization modulation. Acta Pharm. Sin. B 2018, 8, 74–84.
More