Improvements in the growth, yield, and quality of horticultural crops require the development of simply integrated, cost-efficient, and eco-friendly solutions. Hydrogen gas (H2) has been observed to have fertilization effects on soils by influencing rhizospheric microorganisms, resulting in improvements in crop yield and quality. Ample studies have shown that H2 has positive effects on horticultural crops, such as promoting root development, enhancing tolerance against abiotic and biotic stress, prolonging storage life, and improving postharvest quality of fruits, vegetables and cut flowers.
- hydrogen gas
- hydrogen-rich water
- hydrogen nanobubbles
- solid H-storage material
- horticultural crops
- metabolism
1. Introduction
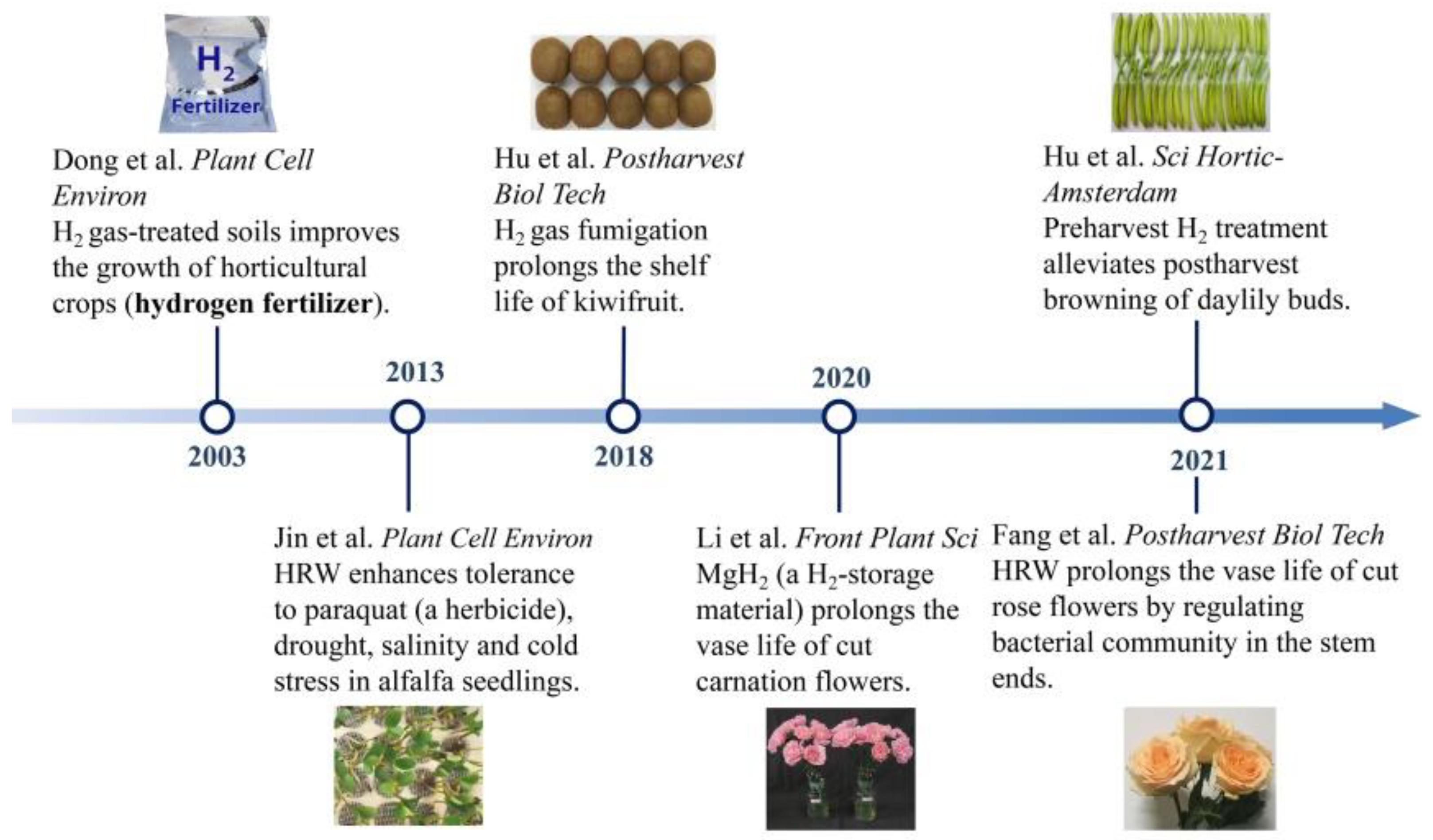
Figure 1. The developing profiles of the application of H2 in horticulture.
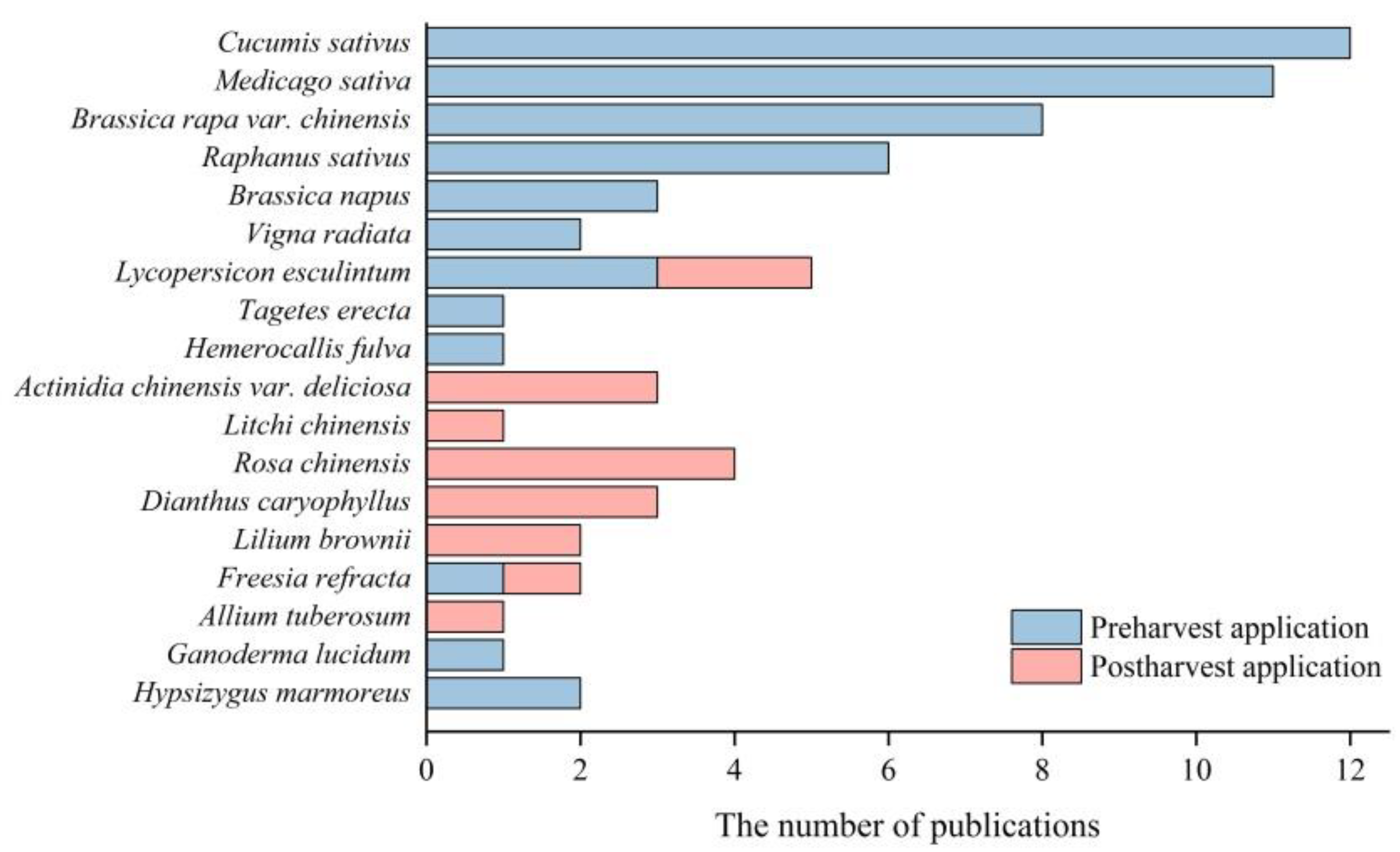
Figure 2. The species of the publications studied on the application of H2 in horticulture.
2. Possible Mechanisms Underlying H2 Responses in Horticultural Crops
2.1. Involved in Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) Metabolism
| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. |
|---|---|---|---|---|---|---|---|---|---|---|
| Brassica rapa var. chinensis | ‘Dongfang 2′ | Preharvest | 1/4 Hoagland’s nutrient solution with H | 2 | (830 µM); the seedlings were pretreated for 48 h | ~415 µM | Alleviates cadmium toxicity | Regulates NR-dependent NO signaling and enhances antioxidant capacity | [53] | [26] |
| 1/4 Hoagland solution with H | 2 | (865 µM); the seedlings were pretreated for 2/3 d (replaced every 12 h) | 865 µM | Reduces cadmium uptake in plant roots | Control of NADPH oxidase encoded by | RbohD | , which operates upstream of IRT1, and regulates root Cd uptake at both the transcriptional and functional levels | [54] | [33] | |
| Medicago sativa | ‘Biaogan’ | Preharvest | HRW (220 µM); the seedlings were pretreated for 12 h | ~110 µM | Enhances tolerance to paraquat | Modulates HO-1 signaling | [11] | |||
| Alleviates aluminum toxicity | Decreases NO production | [41] | [25] | |||||||
| HRW (780 µM); the seedlings were pretreated for 12 h | ~390 µM | Induces osmotic stress tolerance | Regulates H | 2 | O | 2 | and HO-1 signaling | [38] | [23] | |
| NO-mediated proline accumulation and reestablishment of redox balance | [70] | [24] | ||||||||
| Cucumis sativus | ‘Xinchun 4′ | Preharvest | HRW (450 µM); the seedlings were incubated for 2/5 d (changed daily) |
~225 µM | Promotes adventitious rooting | Regulates CO signaling and activates antioxidant system | [34] | [32] | ||
| Regulates NO signaling | [73,74] | [34][35] | ||||||||
| Induces adventitious rooting under cadmium stress | Decreases oxidative damage, increases osmotic adjustment substance content, and regulates rooting-related enzyme activity | [71] | [27] | |||||||
| Cucumis sativus | ‘Jinyou 35′ | Preharvest | HRW (450 µM); the seeds were soaked for 8 h | 450 µM | Enhances cold tolerance | Enhances antioxidant capacity and slows dehydration rate by improving osmotic adjustment ability | [33] | [22] | ||
| Solanum lycopersicum | ‘Baiguoqiangfeng’ | Preharvest | AB@hMSN (10 mg/L); the seedlings were incubated for 2/5 d | ~400 µM | Induces lateral root formation | Modulates NR-dependent NO synthesis, cell cycle regulatory genes, and miRNAs expression | [30] | [36] | ||
| Hypsizygus marmoreus | Preharvest | HRW (1000 µM); the mycelia were cultivated until harvesting | ~250 µM | Increases postharvest quality | Enhances antioxidant defense | [46] | [37] | |||
| Hemerocallis fulva | ‘Dawuzui’ | Preharvest | HRW (1.6 µM); irrigation at the stages of bolting, growing and the day prior to the period of harvest |
~0.8 µM | Promotes daylily bud yield and alleviation of bud browning | Decreases ROS level, increases the unsaturated:saturated fatty acid ratio, endogenous H | 2 | and total phenol content, and reduces PAL and PPO activity | [16] | |
| Actinidia chinesis | ‘Huayou’ | Postharvest | HRW (660 µM); the fruits were soaked for 5 min | ~528 µM | Delays postharvest ripening and senescence | Enhances antioxidant defense | [17] | |||
| Litchi chinensis | ‘Huaizhi’ | Postharvest | HRW (500 µM); the fruits were soaked for 3 min | ~350 µM | Delays the pericarp browning | Induces antioxidant system-related characters | [18] | |||
| Rosa chinensis | ‘Kardinal’; | Lilium brownii | ‘Manissa’ | Postharvest | HRW (450 µM); cut flowers were incubated for vase period (changed daily) | ~225 µM (Rose); ~45 µM (Lily) |
Improves the vase life and quality | Maintains water balance and membrane stability by reducing stomatal size and oxidative damage | [19] | |
| Allium tuberosum | Postharvest | Gas; the leaves were fumigated for storage period (renewed daily) | ~1.2×10 | 3 | µM | Prolongs the shelf life and maintain storage quality | Increases antioxidant capacity | [21] | [28] | |
| Dianthus caryophyllus | ‘Pink Diamond’ | Postharvest | HNW (~500 µM); cut flowers were incubated for 3 d (changed daily) | ~50 µM | Prolongs the vase life | Reduces ROS accumulation and senescence-associated enzyme activities | [26] | [38] | ||
| Rosa chinensis | ‘Carola’ | Postharvest | MgH | 2 | (0.001 g/L); cut flowers were incubated for vase periods (changed daily) | Not shown | Prolongs the vase life | Maintains ROS balance by modulating NO synthesis | [28] | [39] |
| Lilium brownii | ‘Manissa’ | Postharvest | HRW; cut flowers were incubated for vase period (changed daily) | Not shown (1% saturation HRW) | Prolongs the vase life | Regulates NO signaling and regulates the expression of the photosynthesis-related AtpA | [50] | [40] | ||
| Freesia refracta | ‘Red passion’ | Postharvest | HRW (75 µM); cut flowers were pretreated for 12 h | ~0.75 µM | Prolongs the vase life | Improves antioxidant capacity | [51] | [41] | ||
| Eustoma grandiflorum | Postharvest | HRW (780 µM); cut flowers were incubated for vase period (changed daily) | ~78 µM | Prolongs the vase life | Maintains redox homeostasis | [52] | [29] |
2.2. Modulation in Sulfur Compounds’ Metabolism
| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brassica rapa | var. | chinensis | ‘Dongfang 2′ | Preharvest | 1/4 Hoagland’s nutrient solution with H | 2 | ; the seedlings were incubated for 48 h (replaced every 12 h) after removing cadmium stress | Not shown (50% saturation HRW) | Enhances cadmium tolerance | Reestablishes reduced GSH homeostasis | [39] | [45] | |||||
| Medicago sativa | ‘Victoria’ | Preharvest | HRW (220 µM); the seedlings were pretreated for 12 h | ~22 µM | Alleviates cadmium toxicity | Reduces cadmium accumulation and reestablishes GSH homeostasis | [15] | ||||||||||
| Expression regulation of genes relevant to sulfur and glutathione metabolism, resulting in enhanced glutathione metabolism and activating antioxidant defense and cadmium chelation | [76] | [43] | |||||||||||||||
| Decreases oxidative damage, enhances sulfur compound metabolic process, and reestablishes nutrient element homeostasis | [77] | [44] | |||||||||||||||
| Alleviates mercury toxicity | Reduces mercury accumulation and reestablishes redox homeostasis (GSH, AsA, and antioxidant enzymes) | [40] | [46] | ||||||||||||||
| Solanum lycopersicum | ‘Baiguoqiangfeng’ | Preharvest | HRW (780 µM); the seedlings were incubated for 4 d (changed daily) | ~390 µM | Influences lateral root branching | Promotes γ-ECS-dependent GSH production | [35] | [47] | |||||||||
| Ganoderma lucidum | strain HG | Preharvest | HRW (220 µM); added to the medium after 4 days of mycelium culture. | ~11 µM | Regulates morphology, growth, and secondary metabolism | Increases glutathione peroxidase activity under HAc stress | [42] | [48] | |||||||||
| Dianthus caryophyllus | ‘Pink Diamond’ | Postharvest | MgH | 2 | (0.1 g/L MgH | 2 | and 0.1 M PBS (pH 3.4); cut flowers were incubated for vase period (changed daily) | ~400 µM | Prolongs the vase life | H | 2 | S-mediated reestablishment of redox homeostasis and increased transcript levels of | DcbGal | and | DcGST1 | [25] | [49] |
2.3. Involvement in Flavonoids Metabolism
| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raphanus sativus | ‘Qingtou’; | R. sativus | ‘Yanghua’ | Preharvest | HRW (220 µM); 1/4 Hoagland’s nutrient solution with H | 2 | (220 µM H | 2 | ); the seeds were soaked in HRW for 12 h; sprouts were incubated in nutrient solution with H | 2 | for 3 d (replaced every 12 h) under UV-A | ~220 µM |
| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. |
|---|---|---|---|---|---|---|---|---|---|---|
| Regulates anthocyanin synthesis under UV-A | ||||||||||
| Cucumis sativus | ‘XinJinchun No. 4′ | Preharvest | Hoagland’s nutrient solution with H | 2 | (220 µM H | 2 | ); the seedlings were pretreated for 7 d (replaced daily)Reestablishes ROS homeostasis and regulates anthocyanin biosynthesis-related gene expression | [37] | [52] | |
| ~110 µM | Improves heat tolerance | Improves photosynthetic and antioxidant and increases HSP70 content | [ | 32 | ] | [31] | ||||
| ~585 µM | ||||||||||
| Reduces nitrite accumulation during storage | Inhibits/increases the activity and transcript level of NR/NiR | [ | 47 | ] | [ | 59] |
2.5. Modulation of Ion Homeostasis
Ion homeostasis plays an important role in plant tolerance to drought, salinity, and heavy metal stress [93][60]. It has been observed that NH3∙BH3 can decrease Na content and increase K content, resulting in a decreased Na/K ratio in rapeseed seedling roots subjected to NaCl stress [29][61] (Table 5).| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Brassica rapa | var. | chinensis | ‘Dongfang 2’ | Preharvest | 1/4 Hoagland’s nutrient solution with H | 2 | ; the seedlings were pretreated for 1 d (replaced every 12 h) | Not shown (50% saturation HRW) | Reduces cadmium accumulation | Inhibits the expression of | BcIRT1 | and | BcZIP2 | , and reduces cadmium absorption | [94,95] | [62][63] |
| Raphanus. sativus | Brassica rapa | var. | chinensis | ‘Dongfang 2′ | Preharvest | |||||||||||
| Brassica napus | ‘Zhongshuang 11′ | HRW; 1/4 Hoagland’s nutrient solution with H2 (835.1 μM H | 2 | Preharvest); regarding soil cultivation, sprays with HRW (50 mL) at every 12 h for 17 d; for hydroponic solutions, the seedlings were incubated in 1/4 Hoagland solution with H | 2 | for 4 d (replaced every 12 h) with Ca(NO | 3 | ) | 2 | Ammonia borane (NH | 3 | ∙BH~417 µM |
2.6. H2 Is Involved in Phytohormones Signaling
Abscisic acid (ABA), ethylene (ETH), and jasmonate acid (JA) can induce H2, but the specific biosynthesis pathway has yet to be elucidated [31,97][65][66]. For alfalfa drought response, H2 acted as a positive regulator in the ABA signaling cascade to regulate stomatal movement [97][66] (Table 6). H2-modified apoplastic pH by H+-ATPase might be involved in this signaling process. Moreover, H2 differentially increased the transcriptional factor genes involved in ABA signaling, including MYB102, MYC2, and ABF/AREB2 [98][67].| Materials | Treatment Stage | H | 2 | Delivery Methods and Treatment | Effective Concentration of H | 2 | Functions of H | 2 | Mechanism | Ref. No. | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Medicagosativa | ‘Victoria’ | Preharvest | HRW; the seedlings were irrigated for 7 d before 15-d drought treatment | Not shown (50% saturation HRW) | Induces drought tolerance | Modulates stomatal sensitivity to ABA and Apoplastic pH | [97] | [66] | ||||||||||||||||||||
| ‘Yanghua’ | Preharvest | HRW (781 µM); the seedlings were incubated for 48/60 h (replaced every 12 h) under UV-A | 3 | ~781 µM | ; 2 mg/L); the seedlings were incubated for 3 d (changed daily) under NaCl, PEG, or Cd stress | Promotes the biosynthesis of anthocyanin under UV-A | Reduces Ca(NO | 3 | Regulates InsP | 3 | -dependent calcium signaling | ) | [86] | |||||||||||||||
| Medicagosativa | ‘Victoria’ | ~300 µM | 2 | [ | 53 | ] | ||||||||||||||||||||||
| toxicity and improves the growth of seedlings | Enhances antioxidant capacities and reestablishes nitrate homeostasis | [ | Enhances the tolerance against salinity, drought, or cadmium | 44 | Preharvest | 1/4 Hoagland’s nutrient solution with H | 2 | (780 µM H | 2 | ); the seedlings were pretreated for 12 hDecreases cell death rebuilds redox and ion homeostasis, increases proline content, thus reducing cadmium absorption and accumulation | ] | [56] | ||||||||||||||||
| [ | 29 | ] | [ | 61 | ] | ~390 µM | Induces tolerance against osmotic stress | Involved in phytohormone signaling | [98] | [67] | Involved in phytohormones, MAPKs and Ca | 2+ | signaling | Cucumis sativus | ’Jinyou 35′ | Preharvest | HRW (450 µM); the seeds were soaked for 8 h | |||||||||||
| Cucumis sativus | ~450 µM | ‘Xinchun 4′ | [87] | [54] | ||||||||||||||||||||||||
| Enhances lower temperature tolerance | Increases the activities of key photosynthetic enzymes and maintains a high level of carbon and nitrogen metabolism | Preharvest | [ | 90 | ] | [57] | ||||||||||||||||||||||
| Cucumis sativus | ‘Xinchun 4′ | Preharvest | HRW (450 µM); the seedlings incubated for 2/5 d (changed daily) | ~450 µM | HRW (680 µM); the seedlings were incubated for 7 d (changed daily) | Induces adventitious rooting | ~350 µM | Regulates the protein and gene expressions of PM H | + | -ATPase and 14-3-3 mediated by NO. | Induces adventitious rooting | [96 | Ethylene may be the downstream signaling molecule during H | 2 | ] | [64] | -induced adventitious rooting, and proteins RuBisCo, SBPase, OEE1, TDH, CAPX, and PDI may play important roles | [100] | [68] | HRW (220 µM); the seedlings were incubated for 72 h (replaced every 12 h) under short wavelength light | Hypsizygus marmoreus | ~220 µM | PreharvestPromotes anthocyanin accumulation under short wavelength light | HRW (800 µM); mycelia were incubated for 5 d (replaced every 12 h) after removal of cadmium stressPromotes activities and transcription of anthocyanin biosynthesis-related enzyme (including CHS and UFGT) | ~800 µM | Alleviates salinity and heavy metal toxicity | Activates pyruvate kinase, along with its induced gene expression | [43] |
| Cucumis sativus | ‘Lufeng’ | [ | 88 | ] | [55] | |||||||||||||||||||||||
| [ | Preharvest | HRW (220 µM); incubated for 4 d | ~110 µM | Regulates adventitious root development | Regulates HO-1 signaling | 58] | ||||||||||||||||||||||
| [ | 12 | ] | Medicagosativa | ‘Victoria’ | Preharvest | Solanum lycopersicum | ‘Jiafen No. 2′ | |||||||||||||||||||||
| Vigna radiata | ; | Cucumis sativus | ‘Jinchun 4′; | Raphanus sativus | HRW (781 µM); the seedlings were pretreated for 12 h | Postharvest | ‘Yanghua’ | Preharvest | HRW (780 µM); the fruits were soaked for 20 min~390 µM | |||||||||||||||||||
| 1/8 strength Hoagland nutrition solution with H | ||||||||||||||||||||||||||||
| 2 | ||||||||||||||||||||||||||||
| (800 µM); the seedlings were incubated for 5 d (replaced every 12 h) | ||||||||||||||||||||||||||||
| Alleviates UV-B-triggered oxidative damage | Regulates (iso)flavonoids metabolism and antioxidant defense | [ | 83 | ] | [ | 51] |
2.4. H2 Is Involved in Carbon and Nitrogen Metabolism
A previous study has observed that endogenous H2 production can be inhibited by an inhibitor of photosynthetic electron flow, indicating that, in plants, endogenous H2 production may be associated with photosynthesis [| ~480 µM | ||||||||
| Promotes elongation of hypocotyls and roots | ||||||||
| Increases GA and IAA contents in the hypocotyl and the root | [ | 99 | ] | [ | 69 | ] | ||
| Vigna radiata | Preharvest | HRW; seeds were soaked for 3 d | 100/250 µM | Promotes the growth of shoots and roots | Involved in phytohormone signaling | [31] | [65] | |
| Freesia refracta | Preharvest | HRW (75 µM); the bulbs were soaked for 6 h; irrigated HRW at every 7–10 d and total 3 times after scape sticking out | ~37.5 µM | Promotes early flowering; increases the number and diameters of florets | Regulates phytohormone and soluble sugar content | [45] | [70] | |
| Actinidia deliciosa | ‘Xuxiang’ | Postharvest | Gas; the fruits were fumigated for 24 h/12 h + 12 h | ~0.2 µM | Prolongs the shelf life | Decreases ethylene biosynthesis | [22] | [71] |
| Rosa chinensis | ‘Movie star’ | Postharvest | HRW (235 µM); cut flowers were incubated for vase periods (changed daily) | ~2.35 µM | Alleviates postharvest senescence | Inhibits ethylene production and alleviates ethylene signal transduction | [49] | [72] |
3. Conclusions and Prospects
References
- López-García, P.; Moreira, D. The syntrophy hypothesis for the origin of eukaryotes revisited. Nat. Microbiol. 2020, 5, 655–667.
- Gaffron, H. Reduction of carbon dioxide with molecular hydrogen in green algæ. Nature 1939, 143, 204–205.
- Renwick, G.M.; Giumarro, C.; Siegel, S.M. Hydrogen metabolism in higher plants. Plant Physiol. 1964, 39, 303–306.
- Czerkawski, J.W. Fate of metabolic hydrogen in the rumen. Proc. Nutr. Soc. 1972, 31, 141–146.
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694.
- Xie, Y.; Mao, Y.; Lai, D.; Zhang, W.; Shen, W. H2 enhances arabidopsis salt tolerance by manipulating ZAT10/12-mediated antioxidant defence and controlling sodium exclusion. PLoS ONE 2012, 7, e49800.
- Russell, G.; Zulfiqar, F.; Hancock, J.T. Hydrogenases and the role of molecular hydrogen in plants. Plants 2020, 9, 1136.
- Constant, P.; Poissant, L.; Villemur, R. Tropospheric H2 budget and the response of its soil uptake under the changing environment. Sci. Total Environ. 2009, 407, 1809–1823.
- Piche-Choquette, S.; Constant, P. Molecular hydrogen, a neglected key driver of soil biogeochemical processes. Appl. Environ. Microbiol. 2019, 85, e02418-18.
- Dong, Z.; Wu, L.; Kettlewell, B.; Caldwell, C.D.; Layzell, D.B. Hydrogen fertilization of soils—is this a benefit of legumes in rotation? Plant. Cell Environ. 2003, 26, 1875–1879.
- Jin, Q.; Zhu, K.; Cui, W.; Xie, Y.; Han, B.; Shen, W. Hydrogen gas acts as a novel bioactive molecule in enhancing plant tolerance to paraquat-induced oxidative stress via the modulation of heme oxygenase-1 signalling system. Plant Cell Environ. 2013, 36, 956–969.
- Lin, Y.; Zhang, W.; Qi, F.; Cui, W.; Xie, Y.; Shen, W. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8.
- Cao, Z.; Duan, X.; Yao, P.; Cui, W.; Cheng, D.; Zhang, J.; Jin, Q.; Chen, J.; Dai, T.; Shen, W. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084.
- Wu, Q.; Su, N.; Cai, J.; Shen, Z.; Cui, J. Hydrogen-rich water enhances cadmium tolerance in Chinese cabbage by reducing cadmium uptake and increasing antioxidant capacities. J. Plant Physiol. 2015, 175, 174–182.
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724.
- Hu, H.; Li, P.; Shen, W. Preharvest application of hydrogen-rich water not only affects daylily bud yield but also contributes to the alleviation of bud browning. Sci. Hortic. Amst. 2021, 287, 110267.
- Hu, H.; Li, P.; Wang, Y.; Gu, R. Hydrogen-rich water delays postharvest ripening and senescence of kiwifruit. Food Chem. 2014, 156, 100–109.
- Yun, Z.; Gao, H.; Chen, X.; Chen, Z.; Zhang, Z.; Li, T.; Qu, H.; Jiang, Y. Effects of hydrogen water treatment on antioxidant system of litchi fruit during the pericarp browning. Food Chem. 2021, 336, 127618.
- Ren, P.; Jin, X.; Liao, W.; Wang, M.; Niu, L.; Li, X.; Xu, X.; Zhu, Y. Effect of hydrogen-rich water on vase life and quality in cut lily and rose flowers. Hortic. Environ. Biote. 2017, 58, 576–584.
- Cai, M.; Du, H. Effects of hydrogen-rich water pretreatment on vase life of carnation (Dianthus caryophyllus) cut flowers. J. Shanghai Jiao Tong Univ. (Agric. Sci.) 2015, 33, 41–45. (In Chinese)
- Zhou, X.; Joshi, S.; Patil, S.; Khare, T.; Kumar, V. Reactive oxygen, nitrogen, carbonyl and sulfur species and their roles in plant abiotic stress responses and tolerance. J. Plant Growth Regul. 2021.
- Liu, F.; Cai, B.; Sun, S.; Bi, H.; Ai, X. Effect of hydrogen-rich water soaked cucumber seeds on cold tolerance and its physiological mechanism in cucumber seedlings. Sci. Agric. Sin. 2017, 50, 881–889. (In Chinese)
- Jin, Q.; Cui, W.; Dai, C.; Zhu, K.; Zhang, J.; Wang, R.; La, H.; Li, X.; Shen, W. Involvement of hydrogen peroxide and heme oxygenase-1 in hydrogen gas-induced osmotic stress tolerance in alfalfa. Plant Growth Regul. 2016, 80, 215–223.
- Su, J.; Zhang, Y.; Nie, Y.; Cheng, D.; Wang, R.; Hu, H.; Chen, J.; Zhang, J.; Du, Y.; Shen, W. Hydrogen-induced osmotic tolerance is associated with nitric oxide-mediated proline accumulation and reestablishment of redox balance in alfalfa seedlings. Environ. Exp. Bot. 2018, 147, 249–260.
- Chen, M.; Cui, W.; Zhu, K.; Xie, Y.; Zhang, C.; Shen, W. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47.
- Su, N.; Wu, Q.; Chen, H.; Huang, Y.; Zhu, Z.; Chen, Y.; Cui, J. Hydrogen gas alleviates toxic effects of cadmium in Brassica campestris seedlings through up-regulation of the antioxidant capacities: Possible involvement of nitric oxide. Environ. Pollut. 2019, 251, 45–55.
- Wang, B.; Bian, B.; Wang, C.; Li, C.; Fang, H.; Zhang, J.; Huang, D.; Huo, J.; Liao, W. Hydrogen gas promotes the adventitious rooting in cucumber under cadmium stress. PLoS ONE 2019, 14, e212639.
- Jiang, K.; Kuang, Y.; Feng, L.; Liu, Y.; Wang, S.; Du, H.; Shen, W. Molecular hydrogen maintains the storage quality of chinese chive through improving antioxidant capacity. Plants 2021, 10, 1095.
- Su, J.; Nie, Y.; Zhao, G.; Cheng, D.; Wang, R.; Chen, J.; Zhang, S.; Shen, W. Endogenous hydrogen gas delays petal senescence and extends the vase life of lisianthus cut flowers. Postharvest Biol. Tec. 2019, 147, 148–155.
- Del Rio, L.A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837.
- Chen, Q.; Zhao, X.; Lei, D.; Hu, S.; Shen, Z.; Shen, W.; Xu, X. Hydrogen-rich water pretreatment alters photosynthetic gas exchange, chlorophyll fluorescence, and antioxidant activities in heat-stressed cucumber leaves. Plant Growth Regul. 2017, 83, 69–82.
- Chen, Y.; Wang, M.; Hu, L.; Liao, W.; Dawuda, M.M.; Li, C. Carbon monoxide is involved in hydrogen gas-induced adventitious root development in cucumber under simulated drought stress. Front. Plant Sci. 2017, 8, 128.
- Wu, Q.; Huang, L.; Su, N.; Shabala, L.; Wang, H.; Huang, X.; Wen, R.; Yu, M.; Cui, J.; Shabala, S. Calcium-dependent hydrogen peroxide mediates hydrogen-rich water-reduced cadmium uptake in plant roots. Plant Physiol. 2020, 183, 1331–1344.
- Zhu, Y.; Liao, W.; Niu, L.; Wang, M.; Ma, Z. Nitric oxide is involved in hydrogen gas-induced cell cycle activation during adventitious root formation in cucumber. BMC Plant Biol. 2016, 16, 146.
- Zhu, Y.; Liao, W.; Wang, M.; Niu, L.; Xu, Q.; Jin, X. Nitric oxide is required for hydrogen gas-induced adventitious root formation in cucumber. J. Plant Physiol. 2016, 195, 50–58.
- Wang, Y.; Lv, P.; Kong, L.; Shen, W.; He, Q. Nanomaterial-mediated sustainable hydrogen supply induces lateral root formation via nitrate reductase-dependent nitric oxide. Chem. Eng. J. 2021, 405, 126905.
- Chen, H.; Zhang, J.; Hao, H.; Feng, Z.; Chen, M.; Wang, H.; Ye, M. Hydrogen-rich water increases postharvest quality by enhancing antioxidant capacity in Hypsizygus marmoreus. Amb. Express. 2017, 7, 221.
- Li, L.; Yin, Q.; Zhang, T.; Cheng, P.; Xu, S.; Shen, W. Hydrogen nanobubble water delays petal senescence and prolongs the vase life of cut carnation (Dianthus caryophyllus L.) Flowers. Plants 2021, 10, 1662.
- Li, Y.; Li, L.; Wang, S.; Liu, Y.; Zou, J.; Ding, W.; Du, H.; Shen, W. Magnesium hydride acts as a convenient hydrogen supply to prolong the vase life of cut roses by modulating nitric oxide synthesis. Postharvest Biol. Tec. 2021, 177, 111526.
- Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Ma, Z.; Liao, W. Comparative proteomic analysis during the involvement of nitric oxide in hydrogen gas-improved postharvest freshness in cut lilies. Int. J. Mol. Sci. 2018, 19, 3955.
- Song, Y.; Cong, F.; Li, C.; Du, H. Effects of hydrogen-rich water pretreatment on vase life and antioxidant system in cut freesia. J. Shanghai Jiao Tong Univ. (Agric. Sci.) 2018, 36, 1–6. (In Chinese)
- Maruyama-Nakashita, A.; Ohkama-Ohtsu, N. Sulfur assimilation and glutathione metabolism in plants. In Glutathione in Plant Growth, Development, and Stress Tolerance; Hossain, M.A., Mostofa, M.G., Diaz-Vivancos, P., Burritt, D.J., Fujita, M., Tran, L.P., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 287–308.
- Cui, W.; Yao, P.; Pan, J.; Dai, C.; Cao, H.; Chen, Z.; Zhang, S.; Xu, S.; Shen, W. Transcriptome analysis reveals insight into molecular hydrogen-induced cadmium tolerance in alfalfa: The prominent role of sulfur and (homo)glutathione metabolism. BMC Plant Biol. 2020, 20, 58.
- Dai, C.; Cui, W.; Pan, J.; Xie, Y.; Wang, J.; Shen, W. Proteomic analysis provides insights into the molecular bases of hydrogen gas-induced cadmium resistance in Medicago sativa. J. Proteomics 2017, 152, 109–120.
- Wu, Q.; Su, N.; Chen, Q.; Shen, W.; Shen, Z.; Xia, Y.; Cui, J. Cadmium-induced hydrogen accumulation is involved in cadmium tolerance in Brassica campestris by reestablishment of reduced glutathione homeostasis. PLoS ONE 2015, 10, e139956.
- Cui, W.; Fang, P.; Zhu, K.; Mao, Y.; Gao, C.; Xie, Y.; Wang, J.; Shen, W. Hydrogen-rich water confers plant tolerance to mercury toxicity in alfalfa seedlings. Ecotox. Environ. Safe. 2014, 105, 103–111.
- Liu, F.; Lou, W.; Wang, J.; Li, Q.; Shen, W. Glutathione produced by γ-glutamyl cysteine synthetase acts downstream of hydrogen to positively influence lateral root branching. Plant Physiol. Bioch. 2021, 167, 68–76.
- Ren, A.; Liu, R.; Miao, Z.; Zhang, X.; Cao, P.; Chen, T.; Li, C.; Shi, L.; Jiang, A.; Zhao, M. Hydrogen-rich water regulates effects of ROS balance on morphology, growth and secondary metabolism via glutathione peroxidase in Ganoderma lucidum. Environ. Microbiol. 2017, 19, 566–583.
- Li, L.; Liu, Y.; Wang, S.; Zou, J.; Ding, W.; Shen, W. Magnesium hydride-mediated sustainable hydrogen supply prolongs the vase life of cut carnation flowers via hydrogen sulfide. Front. Plant Sci. 2020, 11, 595376.
- Buer, C.S.; Imin, N.; Djordjevic, M.A. Flavonoids: New roles for old molecules. J. Integr Plant Biol 2010, 52, 98–111.
- Xie, Y.; Zhang, W.; Duan, X.; Dai, C.; Zhang, Y.; Cui, W.; Wang, R.; Shen, W. Hydrogen-rich water-alleviated ultraviolet-B-triggered oxidative damage is partially associated with the manipulation of the metabolism of (iso)flavonoids and antioxidant defence in Medicago sativa. Funct. Plant Biol. 2015, 42, 1141.
- Su, N.; Wu, Q.; Liu, Y.; Cai, J.; Shen, W.; Xia, K.; Cui, J. Hydrogen-rich water reestablishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation. J. Agr. Food Chem. 2014, 62, 6454–6462.
- Zhang, X.; Wei, J.; Huang, Y.; Shen, W.; Chen, X.; Lu, C.; Su, N.; Cui, J. Increased cytosolic calcium contributes to hydrogen-rich water-promoted anthocyanin biosynthesis under UV-A irradiation in radish sprouts hypocotyls. Front. Plant Sci 2018, 9, 1020.
- Zhang, X.; Su, N.; Jia, L.; Tian, J.; Li, H.; Huang, L.; Shen, Z.; Cui, J. Transcriptome analysis of radish sprouts hypocotyls reveals the regulatory role of hydrogen-rich water in anthocyanin biosynthesis under UV-A. BMC Plant Biol. 2018, 18, 227.
- Zhang, X.; Wei, J.; Tian, J.; Li, N.; Jia, L.; Shen, W.; Cui, J. Enhanced anthocyanin accumulation of immature radish microgreens by hydrogen-rich water under short wavelength light. Sci. Hortic. Amst. 2019, 247, 75–85.
- Wei, X.; Chen, J.; Chen, H.; Wu, X.; Tian, J.; Su, N.; Cui, J. Hydrogen-rich water ameliorates the toxicity induced by Ca(NO3)2 excess through enhancing antioxidant capacities and re-establishing nitrate homeostasis in Brassica campestris spp. chinensis L. seedlings. Acta. Physiol. Plant 2021, 43, 50.
- Liu, F.; Zhang, X.; Li, D.; Zhai, J.; Bi, H.; Ai, X. Effect of exogenous hydrogen on photosynthetic carbon assimilation and nitrogen metabolism of cucumber seedlings under low temperature. Acta Hortic Sin. 2020, 47, 287–300. (In Chinese)
- Zhang, J.; Hao, H.; Chen, M.; Wang, H.; Feng, Z.; Chen, H. Hydrogen-rich water alleviates the toxicities of different stresses to mycelial growth in Hypsizygus marmoreus. Amb. Express. 2017, 7, 107.
- Zhang, Y.; Zhao, G.; Cheng, P.; Yan, X.; Li, Y.; Cheng, D.; Wang, R.; Chen, J.; Shen, W. Nitrite accumulation during storage of tomato fruit as prevented by hydrogen gas. Int. J. Food Prop. 2019, 22, 1425–1438.
- Zhu, J. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324.
- Zhao, G.; Cheng, P.; Zhang, T.; Abdalmegeed, D.; Xu, S.; Shen, W. Hydrogen-rich water prepared by ammonia borane can enhance rapeseed (Brassica napus L.) seedlings tolerance against salinity, drought or cadmium. Ecotox. Environ. Safe. 2021, 224, 112640.
- Wu, X.; Zhu, Z.B.; Chen, J.H.; Huang, Y.F.; Liu, Z.L.; Zou, J.W.; Chen, Y.H.; Su, N.N.; Cui, J. Transcriptome analysis revealed pivotal transporters involved in the reduction of cadmium accumulation in pak choi (Brassica chinensis L.) by exogenous hydrogen-rich water. Chemosphere 2019, 216, 684–697.
- Wu, X.; Su, N.; Yue, X.; Fang, B.; Zou, J.; Chen, Y.; Shen, Z.; Cui, J. IRT1 and ZIP2 were involved in exogenous hydrogen-rich water-reduced cadmium accumulation in Brassica chinensis and Arabidopsis thaliana. J. Hazard. Mater. 2021, 407, 124599.
- Li, C.; Huang, D.; Wang, C.; Wang, N.; Yao, Y.; Li, W.; Liao, W. NO is involved in H2-induced adventitious rooting in cucumber by regulating the expression and interaction of plasma membrane H+-ATPase and 14-3-3. Planta 2020, 252, 9.
- Zeng, J.; Zhang, M.; Sun, X.; Meijler, M.M. Molecular hydrogen is involved in phytohormone signaling and stress responses in plants. PLoS ONE 2013, 8, e71038.
- Jin, Q.; Zhu, K.; Cui, W.; Li, L.; Shen, W. Hydrogen-modulated stomatal sensitivity to abscisic acid and drought tolerance via the regulation of apoplastic pH in Medicago sativa. J. Plant Growth Regul. 2016, 35, 565–573.
- Felix, K.; Su, J.; Lu, R.; Zhao, G.; Cui, W.; Wang, R.; Mu, H.; Cui, J.; Shen, W. Hydrogen-induced tolerance against osmotic stress in alfalfa seedlings involves ABA signaling. Plant Soil 2019, 445, 409–423.
- Huang, D.; Bian, B.; Zhang, M.; Wang, C.; Li, C.; Liao, W. The role and proteomic analysis of ethylene in hydrogen gas-induced adventitious rooting development in cucumber (Cucumis sativus L.) explants. PeerJ 2020, 8, e8896.
- Wu, Q.; Su, N.; Huang, X.; Ling, X.; Yu, M.; Cui, J.; Shabala, S. Hydrogen-rich water promotes elongation of hypocotyls and roots in plants through mediating the level of endogenous gibberellin and auxin. Funct. Plant Biol. 2020, 47, 771.
- Song, Y.; Li, C.; Xie, P.; Cong, F.; Du, H. Effects of application on stage and methods of hydrogen-rich water on blossom of freesia (Fressia refracta) and related physiological mechanisms. J. Shanghai Jiao Tong Univ. (Agric. Sci.) 2017, 35, 10–16. (In Chinese)
- Hu, H.; Zhao, S.; Li, P.; Shen, W. Hydrogen gas prolongs the shelf life of kiwifruit by decreasing ethylene biosynthesis. Postharvest Biol. Tec. 2018, 135, 123–130.
- Wang, C.; Fang, H.; Gong, T.; Zhang, J.; Niu, L.; Huang, D.; Huo, J.; Liao, W. Hydrogen gas alleviates postharvest senescence of cut rose ‘Movie star’ by antagonizing ethylene. Plant Mol. Biol. 2020, 102, 271–285.
- Finke, C.E.; Leandri, H.F.; Karumb, E.T.; Zheng, D.; Hoffmann, M.R.; Fromer, N.A. Economically advantageous pathways for reducing greenhouse gas emissions from industrial hydrogen under common, current economic conditions. Energ. Environ. Sci. 2021, 14, 1517–1529.
- BNEF-Hydrogen-Economy-Outlook-Key-Messages-30-Mar-2020. Available online: https://data.bloomberglp.com/professional/sites/24/BNEF-Hydrogen-Economy-Outlook-Key-Messages-30-Mar-2020.pdf (accessed on 13 November 2021).
- Wang, X.B.; Schmidt, R.; Yergeau, É.; Constant, P. Field H2 infusion alters bacterial and archaeal communities but not fungal communities nor nitrogen cycle gene abundance. Soil Biol. Biochem. 2020, 151, 108018.