Opuntia species are cacti with high ecological, economic and conservation interest in semiarid environments, particularly in Mexico. Despite the economic and cultural importance of Opuntia, there is a significant lack of knowledge about the flower-visiting insects and their taxonomic identity. Although some Opuntia species could be visited by birds such as hummingbirds, the most dominant taxonomic group of pollinators are the insects.
- insecta
- pollination
- drylands
- core species
- conservation
1. Introduction
2. Opuntia (Cactaceae; Opuntioideae) Flower-Visiting Insects
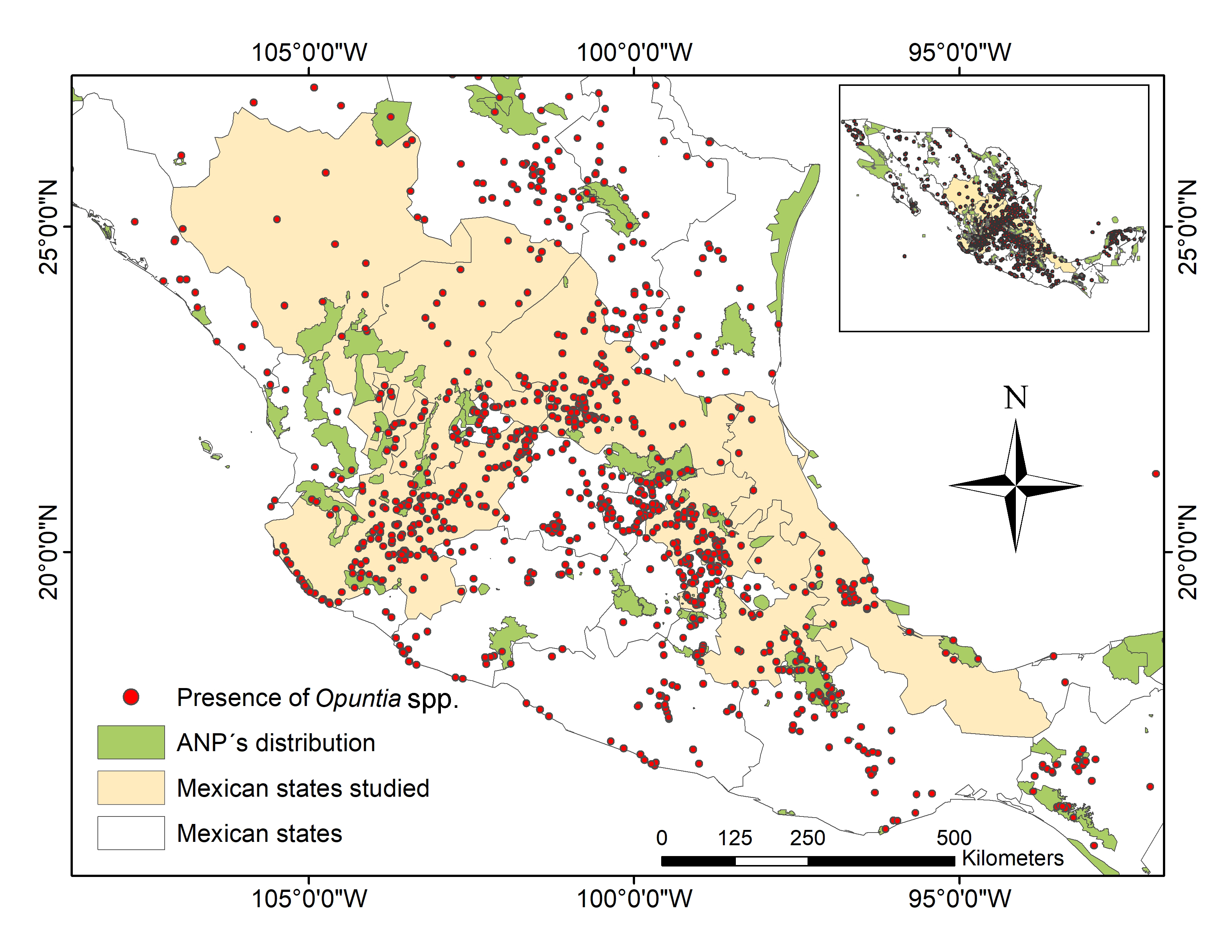
In Mexico, Opuntia species represent one of the most important crops, because they cover about 30% of the country’s land area, and they are mainly distributed throughout arid and semiarid regions (Figure 1) [33]. Mexico is the main producer of prickly pears, supporting 43% of annual world production, which is estimated at 1,060,000 t in an area of 100,000 ha [34]. The area planted with "nopal" in Mexico in 2019 was 45,746 ha, which was mainly in the Mexican High Plateau (central Mexico).
2.1. Timeline of
Opuntia
Studies
2.2. Insect Diversity: Are All the Species Efficient in Pollen Transport?
The studies conducted worldwide indicate that the main flower-visiting insect species in Opuntia spp. are bees (Hymenoptera, Apidae), beetles (Coleoptera), and some lepidopterans (Lepidoptera). However, the effectiveness of pollinator species varied broadly. Early studies [37][38] showed that both bees and beetles were the main groups of insects that pollinated Opuntia plants; however, beetles species were not found to be efficient for pollen transport because they feed on the stamens and petals [37][38]. Therefore, the probability of pollen adhering to their bodies is very low [45]. Moreover, beetles do not fly in search of more flowers, and thus, cross-pollination between plants is limited. In addition, the body size of insects plays a critical role in determining their effectiveness as pollinators [45]. Due to their smaller body size, bees of the genus Perdita sp. (2.0 mm to 10.0 mm) are not considered very effective pollinators, because they can slip into a flower without coming into contact with the stigma [38], thus limiting pollination. The recognized effective pollinators of Opuntia are medium and large-sized bees, such as species belonging to the genera Diadasia, Lithurge, Melissodes, Bombus, Agapostemon, and Megachile [38][46]. The few studies carried out in Argentina, Brazil and Canada were devoted to learning the structure of Opuntia flowers and the diversity of insects that visit them. For example, when bees touch the filaments, they stimulate the movement of sensitive stamens, causing the Opuntia flower to hide most of its pollen from flower visitors [23]. This floral adaptation benefits oligolectic (pollen-specialist) pollinating bees, which can reach the lower layers of the anthers, where 80% of the flower pollen is located [47]. According to the results obtained in the meta-analysis, oligolectic species (such as Ptilothrix fructifera, Lithurgus rufiventris and Cephalocolletes rugata) are the only effective pollinators in Opuntia [47][48][49]. Reliance on oligolectic bees for pollination has been similarly documented for a high number of plant species belonging to different plant families [50][51], including other Cactaceae [41][52]. Therefore, this seems to be quite likely for all Opuntia species as well. However, evidence in studies of other plant taxa highlights that the pollination effectiveness of different flower-visiting insects can also vary depending on other factors, such as season or geographical area, where other insects, including non-oligolectic bees, can act as effective pollinators [50]. For instance, in Lobularia maritima (L.) Desv. (Brassicaceae), the effectiveness of ants in pollination during the summer was comparable to the most effective insects in other periods such as spring [50]. Ants were also the main pollinators of Jatropha curcas L. (Euphorbiaceae) in Mediterranean croplands, unlike what is described for this species in its native distribution area [53]. As Opuntia spp. display a wide blooming period (ranging from early spring to the whole summer), and some of them are important crop species that are commonly grown out of their native area, it would be interesting to examine whether ants (as they are usually documented to be abundant flower visitors) or other animal species might also constitute effective pollinators in certain cases. However, it is also important to bear in mind that ants (as occurs with other common flower-visiting insects) could not generally participate in active Opuntia pollination, as these plants have extrafloral nectaries (EFN). Ecologically, extrafloral nectar is important as a sugared reward for ants to assure ant protection against herbivores [54][55][56]. Having EFN, plants also prevent ants from taking nectar directly from flowers, thus ensuring pollination by other more efficient species [57] in order to achieve successful cross-pollination between different Opuntia individuals. In this regard, ants could be even considered nectar thieves, because the pollen is not transported to other plant individuals as a result of their visit (mainly due to their small body size and absence of hair), and the plant does not benefit from the interaction from the sexual reproduction point of view (e.g., Komamura et al. [58]). Although there are few ant species able to transport pollen, the usual presence of metapleural gland secretion on the integument can also reduce the viability of pollen grains in these particular cases, as documented by Beattie et al. [59], Rostás et al. [60], and Rostás and Tautz [61], therefore, not contributing to the effective cross-pollination. In addition, Rostás et al. [60] found that ants visiting the flowers of Euphorbia seguieriana Neck. do not facilitate outcrossing because the worker ants were mainly plant dwellers; therefore, it was only flying insects that were responsible for pollination, thus increasing the real sexual reproduction measured by success in seed germination rates. Therefore, the role of ants (or other animal species) in Opuntia pollination beyond oligolectic bees should be carefully studied in order to achieve a better understanding of the reproductive ecology of these plants. It is well known that cross-pollination mediated by insects in Opuntia species is crucial for crop productivity [62]. According to Ávila-Gómez et al. [63], Opuntia spp. crop productivity (e.g., number of fruits) is related to the number and species composition of bees. In addition, cross-pollination maintains the genetic diversity in plant populations. Nonetheless, this issue has not been studied for Opuntia spp., and further research must focus on understanding how these complex interactions could support the genetic variability of Opuntia spp. and the impact it might have on the plant–pollinator network structure.2.3. The Relationship between
Opuntia
and Insects in Mexico
Two studies were conducted in the Mapimi Biosphere Reserve, which is part of the Chihuahuan desert. These studies were focused on O. rastrera [40] and O. microdasys [64]; however, few records of flower visitors were obtained. In contrast, Morales-Trejo et al. [32] studied the insect diversity associated with O. pilifera, and they recorded the highest number of insects so far (Hymenoptera, Coleoptera, Hemiptera, Diptera, Neuroptera, and Lepidoptera were identified). Differences in the number of species recorded could be due to the different sampling methods employed. For instance, studies performed on O. rastrera and O. microdasyswere focused on visual records of the frequency that insects pollinated Opuntia flowers. The research conducted on O. pilifera employed a more complete set of harvesting methods mainly consisting of direct collection using entomological nets to capture flying insects and entomological forceps for species found inside the flowers [32]. Santa Anna-Aguayo et al. [65] used a sampling method that consisted of video camera recordings in order to study the behavior of the introduced species A. mellifera and the native species Lithurgus littoralis. They highlighted possible interference by competition between native and non-native bee species that visit the flowers of O. huajuapensis. The use of diverse sampling methods to study the same group of insects often produces different results. This may generate biased conclusions that in turn make it difficult to compare the diversity of species and interactions, and their implications for biodiversity conservation [66]. It is difficult to standardize a single method for insect sampling, and a recommendation to achieve more complete coverage of diversity is to coordinate all data produced by different sampling approaches [67]. In this regard, assimilating information collected using multiple methods, observations, and other sources into a composite database on Opuntia and insect interactions would provide a basis for a better understanding of plant–insect interactions between these groups. Moreover, focusing research efforts on interactions between threatened species of Opuntia and insects will help the conservation and management of these species.2.4. The Role of Core Species in the Community Structure
We found that O. ficus-indica, O. pilifera, and O. monacantha constitute the generalist core of the Opuntia spp. global network (Figure 2; 3), whereas only O. pilifera was core in the Mexican network (Figure 2). In a broad sense, the importance of Opuntia in the network can vary according to the number of studies carried out in each species as well as the sampling effort. This is probably the case for O. ficus-indica, which is a domesticated species with a long history of uses and has been cultivated in several countries [68][69].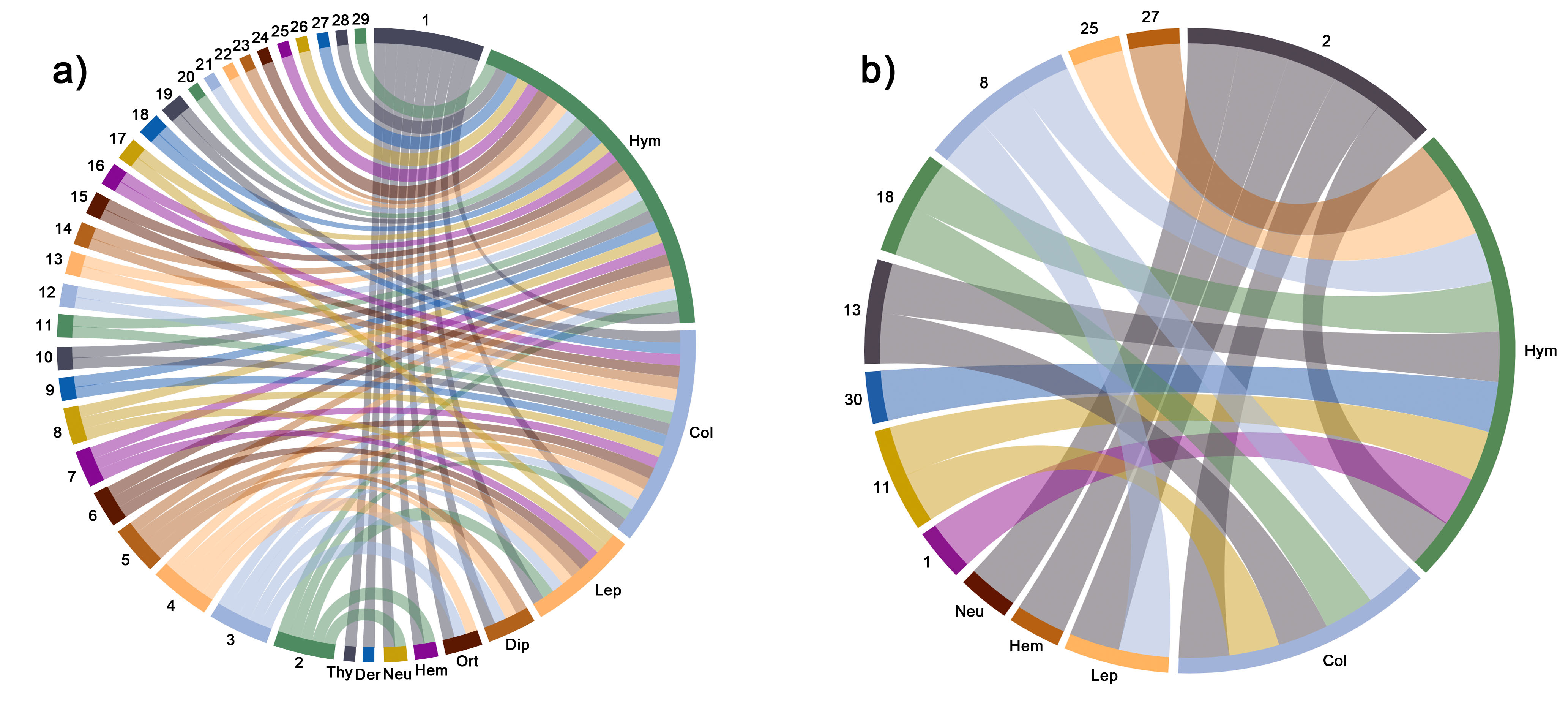
Figure 2. Circular network configuration of flower-visiting orders of insects associated with Opuntia species constructed with the databases of the retrieved articles between 1911 and 2020. Databases were sorted to construct a (a) global network and (b) network of Mexico. Numbers and abbreviations are defined as follows: Hym: Hymenoptera; Col: Coleoptera; Lep: Lepidoptera; Dip: Diptera; Ort: Orthoptera; Hem: Hemiptera; Neu: Neuroptera; Der: Dermaptera; Thy: Thysanoptera. 1: O. ficus-indica; 2: O. pilifera; 3: O: polyacantha; 4: O. fragilis; 5: O. stricta; 6: O. elata; 7: O. maxima; 8: O. tomentosa; 9: O. monacantha; 10: O. lindheimeri; 11: O. robusta; 12: O. anacantha; 13: O. rastrera; 14: O. basilaris; 15: O. humifusa; 16: O. macrorhiza; 17: O. quimilo; 18: O. microdasys; 19: O. littoralis; 20: O. viridirubra; 21: O. spinulifera; 22: O. sulphurea; 23: O. phaeacantha; 24: O. retrorsa; 25: O. streptacantha; 26: O. dillenii; 27: O. huajuapensis; 28: O. macrocentra; 29: O. engelmannii.
 Figure 2. Examples of species that constituted part of the core in both global and Mexico networks. Upper figures show the flowering Opuntia species (a) O. ficus-indica, (b) O. pilifera, and (c) O. monacantha and lower figures show core bees (d) Diadasia rinconis, (e) Agapostemon texanus, and (f) Bombus terrestris. Note that the photos are not scaled.
For this reason, it is not surprising that O. ficus-indica constitutes part of the core species in the global network (Figure 2a; 3a). O. ficus-indica was first introduced to Europe via Spain during the fifteenth century, and it has been cultivated in many parts of the world for various purposes, including carminic acid extraction from Dactylopius coccus Costa, 1835 [18][70]. In this regard, Padrón et al. [20] found that O. maxima is an invader species that behaves as a core species in native communities, where it becomes naturalized. It is known that flowers of alien plants are usually compatible with the body size of native insects [71]. Accordingly, these plants may attract a broad number of native insect pollinators, acting as competitors of the native flora, modifying the network’s structure and the function of the original communities because pollen transported by insects may be dominated by pollen from alien plants [43][72]. In addition, some Opuntia species have no limits to their pollen production, so they might sustain a vast community of insects, thus increasing seed production. Therefore, the probability that these species would expand their range of distribution is high [19].
In the Mexican network (Figure 2b), O. pilifera alone constituted the core species. However, these results are probably influenced by the study conducted by Morales-Trejo et al. [32], who carried out extensive field observations with the goal of recording the highest possible number of insect species and individuals visiting the flowers of O. pilifera and finding seven orders of insects. They also found a temporal segregation of insects in the morning versus the afternoon. This variation can most likely be attributed to the environmental pulses that occur at different times of the day because of changes in the main abiotic factors across the day (such as humidity, temperature, solar irradiation, etc.), which can affect insect behavior and activity [73]. More comprehensive studies on the pollination ecology of Opuntia species from Mexico, ideally conducted with standardized insect harvesting methodologies, would be necessary to ascertain whether O. pilifera is still a core species in the Mexican network, as well as to elucidate the role of other Opuntia species as core species for this network.
In terms of the insect fauna recorded visiting Opuntia flowers, species belonging to the genera Diadasia, Lithurge, Melissodes, Bombus, Agapostemon, and Megachile, which were also the most effective pollinators, were core to the global network. Species of these genera have been documented as the most effective plant pollinators due to their body size [38][46]. The Eurasian honeybee, A. mellifera, had the highest core value. Despite the global economic importance of A. mellifera [74][75], it is an exotic species in the Americas [76], and its effects on the structure and functioning of native communities should also be considered. In this regard, Santa Anna-Aguayo et al. [65] found that females of L. littoralis avoid flowers that have been previously visited by A. mellifera. There is still little information about the competition effect of honeybees on native bees, and therefore, general conclusions cannot be stated. In spite of the fact that A. mellifera is included in the IUCN Red Lists [77][78], it could be of great interest to assess the effects of this species on the conservation of native pollinators in areas where it is not native, such as the American continent. It is important that future research prioritize the study of native bees to discover their conservation status, since competition with A. mellifera could be reducing their populations and affecting the reproductive capacity of Opuntia species.
Bumblebees (Bombus sp.) face important conservation challenges in North America due to their natural habitat transformation for large-scale intensive agricultural production [79][80][81] with many species included in the IUCN Red Lists. Therefore, to maintain (or recover) the conservation status and population levels of bumblebees, it could be useful to maintain the Opuntia species that they visit.
Figure 2. Examples of species that constituted part of the core in both global and Mexico networks. Upper figures show the flowering Opuntia species (a) O. ficus-indica, (b) O. pilifera, and (c) O. monacantha and lower figures show core bees (d) Diadasia rinconis, (e) Agapostemon texanus, and (f) Bombus terrestris. Note that the photos are not scaled.
For this reason, it is not surprising that O. ficus-indica constitutes part of the core species in the global network (Figure 2a; 3a). O. ficus-indica was first introduced to Europe via Spain during the fifteenth century, and it has been cultivated in many parts of the world for various purposes, including carminic acid extraction from Dactylopius coccus Costa, 1835 [18][70]. In this regard, Padrón et al. [20] found that O. maxima is an invader species that behaves as a core species in native communities, where it becomes naturalized. It is known that flowers of alien plants are usually compatible with the body size of native insects [71]. Accordingly, these plants may attract a broad number of native insect pollinators, acting as competitors of the native flora, modifying the network’s structure and the function of the original communities because pollen transported by insects may be dominated by pollen from alien plants [43][72]. In addition, some Opuntia species have no limits to their pollen production, so they might sustain a vast community of insects, thus increasing seed production. Therefore, the probability that these species would expand their range of distribution is high [19].
In the Mexican network (Figure 2b), O. pilifera alone constituted the core species. However, these results are probably influenced by the study conducted by Morales-Trejo et al. [32], who carried out extensive field observations with the goal of recording the highest possible number of insect species and individuals visiting the flowers of O. pilifera and finding seven orders of insects. They also found a temporal segregation of insects in the morning versus the afternoon. This variation can most likely be attributed to the environmental pulses that occur at different times of the day because of changes in the main abiotic factors across the day (such as humidity, temperature, solar irradiation, etc.), which can affect insect behavior and activity [73]. More comprehensive studies on the pollination ecology of Opuntia species from Mexico, ideally conducted with standardized insect harvesting methodologies, would be necessary to ascertain whether O. pilifera is still a core species in the Mexican network, as well as to elucidate the role of other Opuntia species as core species for this network.
In terms of the insect fauna recorded visiting Opuntia flowers, species belonging to the genera Diadasia, Lithurge, Melissodes, Bombus, Agapostemon, and Megachile, which were also the most effective pollinators, were core to the global network. Species of these genera have been documented as the most effective plant pollinators due to their body size [38][46]. The Eurasian honeybee, A. mellifera, had the highest core value. Despite the global economic importance of A. mellifera [74][75], it is an exotic species in the Americas [76], and its effects on the structure and functioning of native communities should also be considered. In this regard, Santa Anna-Aguayo et al. [65] found that females of L. littoralis avoid flowers that have been previously visited by A. mellifera. There is still little information about the competition effect of honeybees on native bees, and therefore, general conclusions cannot be stated. In spite of the fact that A. mellifera is included in the IUCN Red Lists [77][78], it could be of great interest to assess the effects of this species on the conservation of native pollinators in areas where it is not native, such as the American continent. It is important that future research prioritize the study of native bees to discover their conservation status, since competition with A. mellifera could be reducing their populations and affecting the reproductive capacity of Opuntia species.
Bumblebees (Bombus sp.) face important conservation challenges in North America due to their natural habitat transformation for large-scale intensive agricultural production [79][80][81] with many species included in the IUCN Red Lists. Therefore, to maintain (or recover) the conservation status and population levels of bumblebees, it could be useful to maintain the Opuntia species that they visit.
3. Conclusions and Future Perspectives
References
- Hernandez-Hernandez, T.; Brown, J.W.; Schlumpberger, B.O.; Eguiarte, L.E.; Magallón, S. Beyond aridification: Multiple explanations for the elevated diversification of cacti in the new world Succulent Biome. New Phytol. 2014, 202, 1382–1397.
- Anderson, E.F. The Cactus Family; Timber Press: Portland, OR, USA, 2001; p. 776.
- Hunt, D. The New Cactus Lexicon; DH Books: Milborne Port, UK, 2006; Volumes 1 and 2, p. 925.
- Guerrero, P.C.; Majure, L.C.; Cornejo-Romero, A.; Hernández-Hernández, T. Phylogenetic relationships and evolutionary trends in the cactus family. J. Hered. 2019, 110, 4–21.
- González-Medrano, F. Las Zonas Áridas y Semiáridas de México y su Vegetación; Instituto Nacional de Ecología–Secretaria del Medio Ambiente y Recursos Naturales: Mexico City, Mexico, 2012; p. 194.
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 1–7.
- Jiménez-Sierra, C.L. Las cactáceas mexicanas y los riesgos que enfrentan. Revista Digital Universitaria 2011, 12, 5–13.
- Majure, L.C.; Puente, R. Phylogenetic relationships and morphological evolution in Opuntia s. str. and closely related members of tribe Opuntieae. Succ. Plant Res. 2014, 8, 9–30.
- Majure, L.C.; Puente, R.; Griffith, M.P.; Judd, W.S.; Soltis, P.S.; Soltis, D.E. Phylogeny of Opuntia s.s. (Cactaceae): Clade delineation, geographic origins, and reticulate evolution. Am. J. Bot. 2012, 99, 847–864.
- Aliscioni, N.L.; Delbón, N.E.; Gurvich, D.E. Spine function in Cactaceae, a review. J. Prof. Assoc. Cactus 2021, 23, 1–11.
- González-Elizondo, M.S.; González-Elizondo, M.; López-Enríquez, I.L.; Tena-Flores, J.A.; González-Gallegos, J.G.; Ruacho-González, L.; Melgoza-Castillo, A.; Villarreal-Quintanilla, J.A.; Estrada-Castillón, A.E. Diagnóstico del conocimiento taxonómico y florístico de las plantas vasculares del norte de México. Bot. Sci. 2017, 95, 760–779.
- Flores Valdez, C.A.; Aguirre Rivera, J.R. El Nopal como Forraje; Universidad Autónoma Chapingo: Texcoco, México, 1979; p. 91.
- Anaya-Pérez, M.A.; Bautista-Zane, R. El nopal forrajero en México: Del siglo XVI al siglo XX. Agric. Soc. Desarro. 2008, 5, 167–183.
- Ortega-Baes, P.; Sühring, S.; Sajama, J.; Sotola, E.; Alonso-Pedano, M.; Bravo, S.; Godínez-Alvarez, H. Diversity and conservation in the Cactus family. In Desert Plants. Biology and Biotechnology, 1st ed.; Ramawat, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 157–173.
- Dubeux, J.C.B., Jr.; Dos Santos, M.V.F.; Da Cunha, M.V.; Dos Santos, D.C.; De Almeida Souza, R.T.; De Mello, A.C.L.; De Souza, T.C. Cactus (Opuntia and Nopalea) nutritive value: A review. Anim. Feed Sci. and Tech. 2021, 275, 114890.
- Ciriminna, R.; Chavarría-Hernández, N.; Rodríguez-Hernández, A.I.; Pagliaro, M. Toward unfolding the bioeconomy of nopal (Opuntia spp.). Biofuel Bioprod. Bior. 2019, 13, 1417–1427.
- Le Houerou, H.N. Utilization of fodder trees and shrubs in the arid and semiarid zones of West Asia and North Africa. Arid Soil Res. Rehabil. 2000, 14, 101–135.
- Silva, M.A.; Albuquerque, T.G.; Pereira, P.; Ramalho, R.; Vicente, F.; Oliveira, M.B.P.; Costa, H.S. Opuntia ficus-indica (L.) Mill.: A multi-benefit potential to be exploited. Molecules 2021, 26, 951.
- Bartomeus, I.; Vilà, M. Breeding system and pollen limitation in two supergeneralist alien plants invading Mediterranean shrublands. Aust. J. Bot. 2009, 57, 109–115.
- Padrón, B.; Traveset, A.; Biedenweg, T.; Díaz, D.; Nogales, M.; Olesen, J.M. Impact of alien plant invaders on pollination networks in two archipelagos. PLoS ONE 2009, 4, e6275.
- Instituto Nacional de Estadística y Geografía (INEGI). Características Principales del Cultivo del Nopal en el Distrito Federal Caso Milpa Alta; Censo Agropecuario 2007; Instituto Nacional de Estadística y Geografía: Aguascalientes, Mexico, 2007; p. 68.
- Rebman, J.P.; Pinkava, D.J. Opuntia cacti of North America: An overview. Fla. Entomol. 2001, 84, 474–483.
- Reyes-Agüero, J.A.; Valiente-Banuet, A. Reproductive biology of Opuntia: A review. J. Arid Environ. 2006, 64, 549–585.
- Inglese, P.; Mondragon, C.; Nefzaoui, A.; Saenz, C.; Taguchi, M.; Makkar, H.; Louhaichi, M. Ecologia del Cultivo, Manejo y Usos del Nopal; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018; p. 229. Available online: https://hdl.handle.net/20.500.11766/9380 (accessed on 22 October 2021).
- Mandujano, M.C.; Carrillo-Ángeles, I.; Martínez Peralta, C.; Golubov, J. Reproductive biology of Cactaceae. In Desert Plants. Biology and Biotechnology, 1st ed.; Ramawat, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 197–230.
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112.
- Taki, H.; Kevan, P.G. Does habitat loss affect the communities of plants and insects equally in plant–pollinator interactions? Preliminary findings. Biodivers. Conserv. 2007, 16, 3147–3161.
- Senapathi, D.; Biesmeijer, J.C.; Breeze, T.D.; Kleijn, D.; Potts, S.G.; Carvalheiro, L.G. Pollinator conservation—The difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12, 93–101.
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353.
- Van der Kooi, C.J.; Vallejo-Marín, M.; Leonhardt, S.D. Mutualisms and (A) symmetry in plant-pollinator interactions. Curr. Biol. 2021, 31, 91–99.
- Morales-Trejo, J.J.; Sandoval-Ruiz, C.A.; Fascinetto-Zago, P.; Cruzado-Lima, A.L.; Vázquez-Hernández, C. Abundancia y diversidad de visitadores florales de Opuntia pilifera en Zapotitlán Salinas, Puebla. Entomol. Mex. 2014, 1, 1144–1148.
- Pimienta, B.E.; Del Castillo, R.F. Reproductive biology. In Cacti: Biology and Uses; Nobel, P.S., Ed.; University of California Press: Los Angeles, CA, USA, 2002; pp. 75–90.
- Gallegos-Vázquez, C.; Méndez-Gallegos, S.D.J.; Mondragón, J.C. Producción Sustentable de Tuna en San Luis Potosí; Colegio de Postgraduados–Fundación Produce San Luis Potosí: San Luis Potosí, Mexico, 2013; p. 203.
- Potgieter, J.; D’Aquino, S. Fruit production and post-harvest management. In Ecologia del Cultivo, Manejo y Usos del Nopal; Inglese, P., Mondragon, C., Nefzaoui, A., Saenz, C., Taguchi, M., Makkar, H., Louhaichi, M., Eds.; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2018; pp. 51–71. Available online: https://hdl.handle.net/20.500.11766/9380 (accessed on 13 December 2021).
- Bembower, W. Pollination notes from the Cedar Point region. Ohio Nat. 1911, 9, 378–383.
- Cockerell, T.D.A. Two new subgenera of north American bees. Am. Mus. Novit 1922, 47, 1–5.
- Grant, V.; Grant, K.A. Pollination of Opunta basilaris and O. littoralis. Plant Syst. Evol. 1979, 132, 321–325.
- Grant, V.; Grant, K.A.; Hurd, P.D. Pollination of Opuntia lindheimeri and related species. Plant Syst. Evol. 1979, 132, 313–320.
- Del Castillo, R.; González-Espinosa, M. Una interpretación evolutiva del polimorfismo sexual de Opuntia robusta (Cactaceae). Agrociencia 1988, 71, 185–196.
- Mandujano, M.D.C.; Montaña, C.; Eguiarte, L.E. Reproductive ecology and inbreeding depression in Opuntia rastrera (Cactaceae) in the Chihuahuan Desert: Why are sexually derived recruitments so rare? Am. J. Bot. 1996, 83, 63–70.
- Fachardo, A.L.S.; Sigrist, M.R. Pre-zygotic reproductive isolation between two synchronopatric Opuntia (Cactaceae) species in the Brazilian Chaco. Plant Biol. 2020, 22, 487–493.
- Pretto, F.; Celesti-Grapow, L.; Carli, E.; Blasi, C. Influence of past land use and current human disturbance on non-native plant species on small Italian islands. Plant Ecol. 2010, 210, 225–239.
- Lo Verde, G.; La Mantia, T. The role of native flower visitors in pollinating Opuntia ficus-indica (L.) Mill., naturalized in Sicily. Acta Oecologica 2011, 37, 413–417.
- Bartomeus, I.; Vilà, M.; Santamaría, L. Contrasting effects of invasive plants in plant–pollinator networks. Oecologia 2008, 155, 761–770.
- Jauker, F.; Speckmann, M.; Wolters, V. Intra-specific body size determines pollination effectiveness. Basic Appl. Ecol. 2016, 17, 714–719.
- Osborn, M.M.; Kevan, P.G.; Lane, M.A. Pollination biology of Opuntia polyacantha and Opuntia phaeacantha (Cactaceae) in southern Colorado. Plant Syst. Evol. 1988, 159, 85–94.
- Cota-Sánchez, J.H.; Almeida, O.J.G.; Falconer, D.J.; Choi, H.J.; Bevan, J. Intriguing thigmonastic (sensitive) stamens in the plains prickly pear Opuntia polyacantha (Cactaceae). Flora 2013, 208, 381–389.
- Schlindwein, C.; Wittmann, D. Stamen movements in flowers of Opuntia (Cactaceae) favour oligolectic pollinators. Plant Syst. Evol. 1997, 204, 179–193.
- Lenzi, M.; Orth, A.I. Floral visitors of the Opuntia monacantha (Cactaceae) in sandbank of the Florianópolis, SC, Brazil. Acta Biológica Paranaense 2011, 40, 19–32.
- Gómez, J.M. Effectiveness of ants as pollinators of Lobularia maritima: Effects on main sequential fitness components of the host plant. Oecologia 2000, 122, 90–97.
- Maubecin, C.C.; Boero, L.; Sérsic, A.N. Specialisation in pollen collection, pollination interactions and phenotypic variation of the oil-collecting bee Chalepogenus cocuccii. Apidologie 2020, 51, 710–723.
- Arroyo-Pérez, E.; Jiménez-Sierra, C.L.; Zavala Hurtado, J.A.; Flores, J. Shared pollinators and sequential flowering phenologies in two sympatric cactus species. Plant Ecol. Evo. 2021, 154, 28–38.
- Samra, S.; Samocha, Y.; Eisikowitch, D.; Vaknin, Y. Can ants equal honeybees as effective pollinators of the energy crop Jatropha curcas L. under Mediterranean conditions? GCB Bioenergy 2014, 6, 756–767.
- Pickett, C.H.; Clark, W.D. The function of extrafloral nectaries in Opuntia acanthocarpa (Cactaceae). Am. J. Bot. 1979, 66, 618–625.
- LeVan, K.E.; Hung, K.L.J.; McCann, K.R.; Ludka, J.T.; Holway, D.A. Floral visitation by the Argentine ant reduces pollinator visitation and seed set in the coast barrel cactus, Ferocactus viridescens. Oecologia 2014, 174, 163–171.
- Mauseth, J.D.; Rebmann, J.P.; Machado, S.R. Extrafloral nectaries in cacti. Cactus Succul. J. 2016, 88, 156–171.
- Wagner, D.; Kay, A. Do extrafloral nectaries distract ants from visiting flowers? An experimental test of an overlooked hypothesis. Evol. Ecol. Res. 2002, 4, 293–305.
- Komamura, R.; Koyama, K.; Yamauchi, T.; Konno, Y.; Gu, L. Pollination contribution differs among insects visiting Cardiocrinum cordatum flowers. Forests 2021, 12, 452.
- Beattie, A.J.; Turnbull, C.; Knox, R.B.; Williams, E.G. Ant inhibition of pollen function—A possible reason why ant pollination is rare. Am. J. Bot. 1984, 71, 421–426.
- Rostás, M.; Bollmann, F.; Saville, D.; Riedel, M. Ants contribute to pollination but not to reproduction in a rare calcareous grassland forb. PeerJ 2018, 6, e4369.
- Rostás, M.; Tautz, J. Ants as pollinators of plants and the role of floral scents. In All Flesh is Grass. Cellular Origin, Life in Extreme Habitats and Astrobiology, 1st ed.; Dubinsky, Z., Seckbach, J., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 16, pp. 149–161.
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821.
- Ávila-Gómez, E.S.; Meléndez-Ramírez, V.; Castellanos, I.; Zuria, I.; Moreno, C.E. Prickly pear crops as bee diversity reservoirs and the role of bees in Opuntia fruit production. Agric. Ecosyst. Environ. 2019, 279, 80–88.
- Piña, H.H.; Montaña, C.; Mandujano, M. Fruit abortion in the Chihuahuan-Desert endemic cactus Opuntia microdasys. Plant Ecol. 2007, 193, 305–313.
- Santa Anna-Aguayo, A.I.; Schaffner, C.M.; Golubov, J.; López-Portillo, J.; García-Franco, J.; Herrera-Meza, G.; Martínez, A.J. Behavioral repertoires and interactions between Apis mellifera (Hymenoptera: Apidae) and the native bee Lithurgus littoralis (Hymenoptera: Megachilidae) in flowers of Opuntia huajuapensis (Cactaceae) in the Tehuacan desert. Fla. Entomol. 2017, 100, 396–402.
- McCravy, K.W. A review of sampling and monitoring methods for beneficial arthropods in agroecosystems. Insects 2018, 9, 170.
- Montgomery, G.A.; Belitz, M.W.; Guralnick, R.P.; Tingley, M.W. Standards and best practices for monitoring and benchmarking insects. Front. Ecol. Evol. 2021, 8, 579193.
- Vigueras, A.L.; Portillo, L. Uses of Opuntia species and the potential impact of Cactoblastis cactorum (Lepidoptera: Pyralidae) in Mexico. Fla. Entomol. 2001, 84, 493–498.
- Griffith, M.P. The origins of an important cactus crop, Opuntia ficus-indica (Cactaceae): New molecular evidence. Am. J. Bot. 2004, 91, 1915–1921.
- Barbera, G.; Carimi, F.; Inglese, P. Past and present role of the indian-fig prickly-pear (Opuntia ficus-indica (L) Miller, Cactaceae) in the agriculture of Sicily. Econ. Bot. 1992, 46, 10–20.
- Stang, M.; Klinkhamer, P.G.; Van Der Meijden, E. Size constraints and flower abundance determine the number of interactions in a plant–flower visitor web. Oikos 2006, 112, 111–121.
- Lopezaraiza-Mikel, M.E.; Hayes, R.B.; Whalley, M.R.; Memmott, J. The impact of an alien plant on a native plant–pollinator network: An experimental approach. Ecol. Lett. 2007, 10, 539–550.
- Ness, J.H. Hot spots and hot moments for on-plant foraging by ants within the flora of warm North American Deserts. Am. Midl. Nat. 2020, 183, 145–163.
- Vithanage, V. The role of the European honeybee (Apis mellifera L.) in avocado pollination. J. Hortic. Sci. 1990, 65, 81–86.
- Rizzardo, R.A.; Milfont, M.O.; Silva, E.; Freitas, B.M. Apis mellifera pollination improves agronomic productivity of anemophilous castor bean (Ricinus communis). Anais da Academia Brasileira de Ciências 2012, 84, 1137–1145.
- Moritz, R.F.; Härtel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Ecoscience 2005, 12, 289–301.
- De la Rúa, P.; Paxton, R.J.; Moritz, R.F.A.; Roberts, S.; Allen, D.J.; Pinto, M.A.; Cauia, E.; Fontana, P.; Kryger, P.; Bouga, M.; et al. Apis mellifera. The IUCN Red List of Threatened Species 2014, e.T42463639A42463665. Available online: https://ec.europa.eu/environment/nature/conservation/species/redlist/downloads/European_bees.pdf (accessed on 13 March 2021).
- Paudel, Y.P.; Mackereth, R.; Hanley, R.; Qin, W. Honeybees (Apis mellifera L.) and pollination issues: Current status, impacts, and potential drivers of decline. J. Agric. Sci. 2015, 7, 93–109.
- Grixti, J.C.; Wong, L.T.; Cameron, S.A.; Favret, C. Decline of bumble bees (Bombus) in the North American Midwest. Biol. Conserv. 2009, 142, 75–84.
- Jacobson, M.M.; Tucker, E.M.; Mathiasson, M.E.; Rehan, S.M. Decline of bumble bees in northeastern North America, with special focus on Bombus terricola. Biol. Conserv. 2018, 217, 437–445.
- Hatfield, R.; Jepsen, S.; Thorp, R.; Richardson, L.; Colla, S.; Foltz Jordan, S. Bombus pensylvanicus. The IUCN Red List of Threatened Species 2015, e.T21215172A21215281. Available online: https://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T21215172A21215281.en (accessed on 25 February 2021).