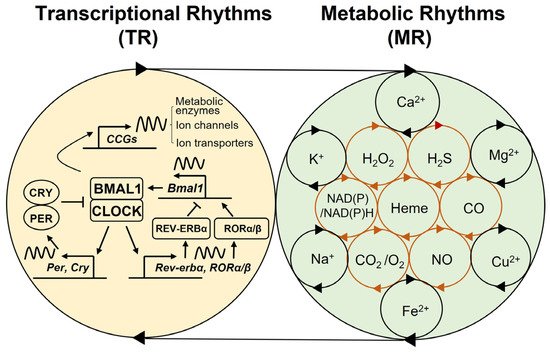
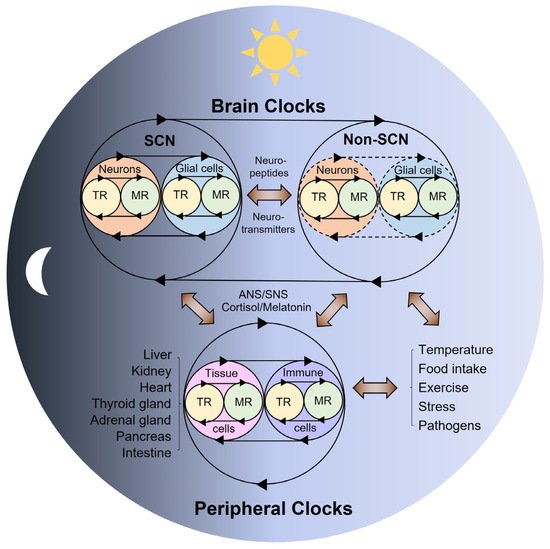
Figure 2. Coupled-tissue oscillators. Reciprocal crosstalk between the brain and peripheral clocks. The coupled TR and MR oscillators are thought to be commonly present across all body cells. Neurons and glial cells (e.g., astrocytes, microglia) interact to form the SCN central clock and non-SCN clocks in the brain. These autonomous brain clocks communicate with each other via neurotransmitters or neuropeptides, and with multiple peripheral tissue clocks via systemic innervations (ANS/SNS) or hormonal signals (e.g., cortisol, melatonin) in response to light–dark cycles. On the other hand, peripheral organs possess tissue autonomous clocks that can respond to non-photic physiological and environmental cues (e.g., temperature, food intake, exercise, stress) and provide feedback that influences the brain clocks via immune, metabolic, and endocrine signals. TR—transcriptional rhythms; MR—metabolic rhythms; ANS—autonomic nervous system; SNS—sympathetic nervous system.
The intracellular oscillators, in approximately 20,000 neurons, comprise the hypothalamic suprachiasmatic nucleus (SCN), a central clock in the rodent brain. The SCN in humans has been found to contain a total number of neurons close to 100,000, though these numbers decline with age
[46][47][48][46,47,48]. The SCN consists of different subtypes of neurons that express the neurotransmitter c-aminobutyric acid (GABA), an inhibitory neurotransmitter in the brain, alongside a range of neuropeptides such as vasoactive intestinal peptide (VIP), arginine vasopressin (AVP), and their cognate receptors (VPAC2 and AVPR1A/B)
[49][50][51][52][49,50,51,52]. These GABAergic/peptidergic SCN neurons interact among themselves or with the other neurons in extra-SCN brain regions, constituting the main output pathway of the clock. Notably, a recent study has demonstrated that the VIP-VPAC2 neuropeptidergic axis plays a central role in mediating the endogenous pacemaking function of the SCN circadian circuit
[53]. In line with this, the suprachiasmatic VIP neurons (SCN
VIP) have been shown to be required for normal circadian behaviors via functional connectivity between SCN
VIP neurons and dorsomedial hypothalamic neurons
[54]. Interestingly, recent single-cell RNA sequencing (scRNASeq) studies with mouse SCN slice revealed novel neuronal phenotypes and interaction networks involved in the central clock, including the identification of five SCN neuronal subtypes, with cluster-specific marker genes of VIP, AVP, gastrin-releasing peptide (GRP), cholecystokinin (CCK), and the cell adhesion regulator C1ql3
[55][56][55,56]. Additional scRNASeq study has also identified a subgroup of cells expressing Prokineticin2 (Prok2) and its cognate receptor (ProkR2) found to be topologically and functionally distinct pacemaking element of the central clock
[57]. Thus, these studies highlight diverse cellular sub-populations within the neuropeptidergic topology of the SCN, which may differently contribute to the central pacemaker function. Notably, recent studies have shown that astrocytes harbor distinct rhythmic properties, such as anti-phasic Ca
2+ rhythms, that direct the circadian rhythmicity of SCN neurons and behavior
[58][59][60][58,59,60]. This work suggests the importance of bipartite intercellular communication between astrocytes and neurons in modulating SCN pacemaker functions, beyond neuronal regulation of the central clock.
Along with endogenous circadian pacemaking activity, the SCN central clock also mediates the periodic synchronization of internal body rhythms with external day and night cycles by communicating retinal light information received from the retinohypothalamic tract (RHT) to peripheral clock systems
[26]. In a hierarchical organization model, the SCN master clock coordinates the circadian phases of individual subordinate clocks in other brain regions via rhythmic release of neurotransmitters and neuropeptides as well as in peripheral tissues via systemic hormonal secretion and neural innervations, thus generating rhythmic output physiology and behaviors that are in keeping with the daily changes in environment and needs
[61][62][63][64][61,62,63,64]. For example, the SCN coordinates the rhythmic anti-phasic secretion of the night sleep hormone melatonin in the pineal gland and the morning stress hormone glucocorticoid in the adrenal glands via the sympathetic nervous system, ensuring daily rhythms in sleep/wake, as well as neural, metabolic, and immune functions
[65][66][67][65,66,67]. In addition, the brain master clock controls peripheral clock functions in the heart, kidney, pancreas, lung, intestine, and thyroid glands by circadian regulation of the autonomic nervous system
[68][69][70][71][72][73][68,69,70,71,72,73].
Besides SCN-driven clock entrainment, a growing number of studies have reported that non-SCN brain regions and peripheral tissues possess their own autonomous, entrainable oscillators that influence not only circadian functions in the SCN and neighboring local clocks, but also whole body rhythms via neural, hormonal, and metabolic feedback mechanisms
[13][74][75][76][77][78][79][80][13,74,75,76,77,78,79,80]. The SCN receives a myriad of nonphotic input, arousal, feeding behavior, locomotor activity, immune function, blood pressure, and melatonin, which are all able to adjust and synchronize the SCN
[81][82][83][84][81,82,83,84]. The SCN can receive this feedback through its large array of reciprocal neuronal connections with different hypothalamic regions, such as the arcuate nucleus (ARC), intergeniculate leaflet (IGL), nucleus tractus solitarius (NTS), dorsal raphe, and dorsomedial hypothalamus, allowing these nuclei to convey nonphotic feedback to the SCN and thus adjusting circadian rhythmicity
[85][86][87][88][89][85,86,87,88,89].
Beyond photic entrainment, multiple physiological and environmental cues (e.g., food intake, gut microbial products, redox cofactors, metal ions, metabolic gases) control non-SCN and peripheral clock functions, which, in turn, impact the entire host clock system via neural and immune–metabolic circuits
[12][22][72][90][91][92][93][94][95][12,22,72,90,91,92,93,94,95]. These findings suggest that systemic circadian rhythms are achieved through multi-modal regulation of tightly coupled body clocks according to daily changes that occur in the internal and external environments.
3. Conclusions
In recent decades, extensive chronobiological research has expanded our understanding of the functional roles and mechanisms of the circadian clockwork in human health and diseases. Thus, research trends in chronobiology have undergone a paradigm shift in many ways, particularly changing from hierarchical models to more integrated ones for understanding the circadian clock mechanism. In this regard, the overall evidence points to bidirectional crosstalk between transcriptional and metabolic rhythms, neuronal and glial clocks, SCN and non-SCN clocks, as well as brain and peripheral clocks (Figure 1, Figure 2). Our growing knowledge of the interactive nature of clock regulatory systems is expected to not only diversify our understanding of circadian physiology and pathophysiology but also increase our capacity to harness chronobiological knowledge to improve the prevention and treatment of multiple circadian-related disorders.