Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Paweł Wroński and Version 2 by Conner Chen.
The most common type of bladder cancer (BC) is urothelial carcinoma arising from stratified epithelium-urothelium. Layers of cells are arranged into strata perched on the basement membrane (BM), then on lax connective tissue, followed by the muscular part of the bladder wall. The breeching of the urothelial BM, facilitated by an aberrant activation of matrix metalloproteinases (MMP) is particularly perilous. Inhibition of activation of these proteinases constitutes a logic opportunity to restrain progression. Recent studies revealed significant anticancer potential of natural phytochemicals. Especially, curcumin has emerged as a one of the most promising phytochemicals and showed its efficacy in several human malignancies.
- bladder cancer
- basement membrane proteins
- phytochemicals
- curcumin
- matrix metalloproteinases
[1][2][3][4][5][6][7][8][9][10][11][12][13][14][15][16][17][18][19][20][21][22][23][24][25][26][27][28][29][30][31][32][33][34][35][36][37][38][39][40][41][42][43][44][45][46][47][48][49][50][51][52][53][54][55][56][57][58][59][60][61][62][63][64][65][66][67][68][69][70][71][72][73][74][75][76][77][78][79][80][81][82][83][84][85][86][87][88][89][90][91][92][93][94][95][96][97][98][99][100][101][102][103][104][105][106][107][108][109][110][111][112][113][114][115][116]1. Biological Properties of Curcumin in Cancer Processes
For ages, herb- and plant-derived medicines have been a staple in therapy, even without exact knowledge of contained active components or the mechanism of action. Several extracted pure substances, their derivatives, or synthetic equivalents are widely used even today. Contemporary molecular biology and analyses of cell signaling pathways reveal mechanisms of action, and potentially new pharmacological uses of known old nutrients present in common food. Natural ingredients from food (nutrients, nutraceuticals, phytochemicals) or their derivatives can replace or synergistically reinforce the action of current medicines when combined with standard therapeutic regimens [1][47].
New findings provide a very promising perspective of selected compounds as substances with high therapeutic potential for human malignancies. The potential is promising for 2000 plants, each containing numerous molecules, which are under laboratory and clinical evaluation now [2][48]. Despite their demonstrated anti-cancer efficacy, the precise molecular mechanism of activity is not clearly established [3][49]. The activity of phytochemicals tested in vitro on experimental cell lines differs from effects observed on clinical settings for the multiplicity of additional interactions and interdependencies between different cells and tissues [4][50]. Last but not least, relatively inexpensive and easily available natural phytochemicals and derivatives may be difficult to compete with expensive, extolled drugs.
An example of a nutrient exerting potent biological activity is curcumin, currently the most intensively tested phytochemical. Widely used as a food additive, it shows a lack of any toxicity. [5][6][7][8][51,52,53,54]. Its curative properties, as used in traditional Asian medicine for the treatment of nonhealing wounds and gastrointestinal diseases, present safety even in high doses [9][10][55,56]. Recently, curcumin gained attention due its antioxidant, antiatherosclerotic, anticancer (antiproliferation, anti-invasive, and antimetastatic) activity [11][34].
Curcumin is an active compound from the turmeric rhizome Curcuma longa. Raw or dried-pulverized turmeric is commonly used as a spice in Asia [9][12][55,57]. Up to 235 bioactive compounds have been extracted from turmeric. The most abundant are curcuminoids: curcumin, demethoxycurcumin, and bisdemetoxycurcumin, in a proportion 7:2:1, respectively [13][58]. Nowadays, curcumin is used in food processing as a spice and as a natural pigment. Its chemical structure was first described by Polish chemists in 1910 [14][59]. Chemically, curcumin is a polyphenol-diferuloylmethane: (1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptane-3,5-dione) [6][9][52,55]. It has two constitutional isomers: enol and b-diketone tautomeric forms. The former exists predominantly in solutions, the latter is important for free radicals’ scavenging ability [9][15][55,60].
The degradation of the basement membrane—a prerequisite for tumor invasion—is mediated mainly by MMP-2 and -9. Abundant data present that the anticancer and antimetastatic effect of curcumin is linked to inhibition of MMP activity. This potential of curcuminoids is dose- and time-dependent, as demonstrated on in vitro cultured cells [9][16][55,61]. In addition to downregulation of MMPs, curcumin upregulates expression of tissue inhibitors of MMP, especially TIMP-2. Further, curcumin counteracts metastasis formation by the restriction of cancer cell adhesion molecules, allowing for binding to ECM [17][62]. Curcumin-related decreased expression of MMP-2 and -9 also inhibits angiogenesis [18][63]. The mechanisms of such an impressive range of activities have not been fully elucidated yet. Diverse anticancer and antioxidant properties seem to have common crucial elements linking these activities. Curcumin inhibits cell signaling pathways, and hampers expression of several cancer-related genes (for instance, COX-2, TNF, cyclin D1). Several papers present that its main impact point focuses on nuclear factor kappa B (NF-κB) and STAT pathways regulating the above genes (Figure 2) [9][55]. NF-κB is a ubiquitous inducible transcription factor present in all animal cells, and is the pivotal element of the pathway transmitting extracellular signals into the nucleus to stimulate the expression of numerous genes. Thus, curcumin appears to be a master regulator of almost all cellular processes involved in cell proliferation, survival, and response to external factors. It binds to promotors of targeted genes, and activates their transcription [19][64]. Substances which can control and modulate NF-κB, in turn, can control and modulate the function of genes and the cell’s fate. Curcumin has properties to suppress the activation of NF-κB (Figure 2). Binding any ligand to the cell surface receptor induces a specific kinase, IKK, which phosphorylates, and thus, inactivates an inhibitor of NF-κB-IκB. The promoters of MMP genes have binding sites for NF-κB. Then, NF-κB dimerizes and translocates to the nucleus to promote transcription of any given gene for cell membrane-bound metaloproteinase MT1-MMP. MT1- MMP proteolyse pro-MMP-2 into active MMP-2. Curcumin interrupts this pathway by blocking IκB kinase (IKK), and thwarts MMP-2 activation, and subsequent extracellular matrix degradation and invasion [20][65]. Concurrently, other NF-κB dependent genes and their products (MMP2,9, cyclin D1, CoX-2, Ras, iNOS, bcl-2, bclxl) are downregulated and suppressed, so cell proliferation and migration are inhibited. A similar protecting mechanism of curcumin has been described for restricting NF-κB benzopyrene (a cigarette smoke compound)-related activation of genes expression in selected lung carcinoma cell lines [21][66]. Additionally, curcumin through bcl-2/bclxl-related inhibition of NF-κB pathways activates caspase-mediated cell death [21][22][66,67]. An alternative pathway of curcumin suppression on MMP-2 runs through the Akt serine/threo-nine protein kinase axis, as proved by curcumin’s anti-lymphangiogenic effect on an experimental line of lymphatic endothelial cells (Figure 1) [22][67].
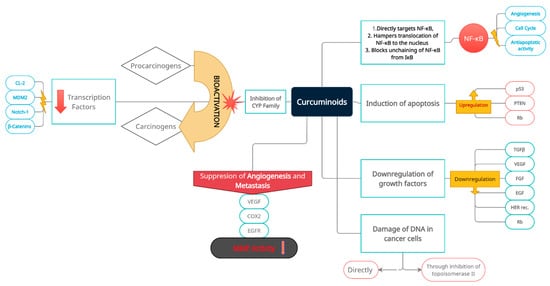
Figure 1. Multidirectional and multilevel preventive and inhibitory effects of curcuminoids on oncogenesis, with a special emphasis on induction and progression of bladder cancer, and its subsequent development. Prevention and inhibition of neoplastic transformation and progression is achieved through induction of apoptosis, inactivation of cancer-related transcriptional pathways (nuclear transcription factors and related oncogenes), cancer cell DNA damage (direct and indirect), inactivation of extracellular carcinogens, downregulation of growth factors, activation of cancer-suppressing genes, suppression of angiogenesis, and metastatic niche formation. Consecutive boxes of the diagram group intra- and extracellular molecular targets of curcumin.
A probable mechanism of MMP inhibition relies on zinc ions locking through the metal-binding moiety of curcumin [9][10][55,56]. Another inhibitory mechanism of curcumin on MMP-2 and -9, and on proliferation signaling pathways is in the decreasing expression of extracellular signal-regulated protein kinase (p-ERK1/2), associated with cell proliferation and survival. A transcription factor regulator p-ERK1/2 is involved in signal transduction from the cell membrane to the nucleus [9][23][55,68]. A mammalian target of rapamycin (mTOR) pathways is a next target for curcumin inhibitory activity, and through the phosphatidylinositol 3-kinase/Akt/IkBk kinase axis, MMP-2 and -9 expression are also significantly suppressed [9][55]. Further, curcumin also selectively targets MMP2 by restricting expression of mRNA for MMP2 [24][69]. Curcumin-induced apoptosis was experimentally presented in prostate cancer cell lines as propagated through TNF-a-related apoptosis-inducing ligand (TRAIL).
Because NF-κB is a component of pro-inflammatory signaling pathway, and is involved in the production and release of inflammatory cytokines such as TNF-alfa, which, in turn, activate MMP, this mode of activity can also potentiate the antitumor properties of curcumin [25][70]. Tumor-infiltrating lymphocytes express upregulation of MMP-9, which, in turn, promotes neoplastic proliferation and invasion. Curcumin inhibits this trait by suppressing the cancer propagatory influence of cytokines excreted from the inflammatory transformed peritumoral extracellular matrix [26][71]. Further, decrease of MMP-2 expression, and an increase of TIMP-1 intensified antiproliferative and anti-invasive effects on a breast cancer cell line [9][26][55,71]. Another mechanism of indirect downregulation of MMPs expression relies on the ability of curcumin to suppress ECM inducer (EMMPRIN, CD147), crucial for cancer cell adhesion, tumor invasion, and metastasis. This cell membrane glycoprotein is abundant in cancer cells, and stimulates production, secretion, and activation of MMPs by cells (monocytes and macrophages) present in tumors surrounding the EC stroma. CD147 also binds MMP at the cell surface, and helps the cell to directly degrade surrounding pericellular ECM and further invasion [27][72]. Curcumin inhibition of MMP-2 and -9 activity also restores blood–brain barrier integrity, as reported for cerebral ischemic injury [11][34]. Another positive involvement on vascular pathology has been presented. Curcumin reduces activity of MMP-2 and -9 during plaque formation, and suppresses NF-κB and MMP-9 expression in vascular smooth muscle cells, contributing to atherosclerosis formation (Figure 1) [9][55].
Unfortunately, important issues related to the use of curcuminoids still remain unresolved. Clinical application of curcumin is hampered by its somewhat unfavorable biological properties. Bioavailability of curcumin is low because of its limited water solubility at acidic and neutral pH, resulting in poor intestinal absorption, and also its rapid degradation by glucuronidation in the intestinal wall and in the liver [7][28][53,73]. To overcome barriers related to the limited bioavailability of curcumin, a number of modifications to the administration or its chemical structure have been tested. Research has been aimed at improving the absorption of curcumin, on slowing its metabolism, and on linking it with other substances to achieve synergy; thus, enhancing their therapeutic activities. Co-administration with piperine enhances intestinal absorption, and diminishes biodegradation (glucuronidation) of curcumin, and thus, increases curcumin bioavailability up to 20× [5][51]. Curcumin-piperine formulation as a nanoparticle is a useful solution, and has been extensively tested. [29][30][74,75]. The combination with phosphatidyl-choline increases oral absorption of curcumin in humans 5-fold [9][55]. Another formulation combines both molecules into liposomes as one stable water-soluble delivery system [5][51]. Experimental delivery systems couple curcumin with chemotherapeutics [31][32][76,77]. Liposomal formulation with polyethylene glycol, and complexes with phospholipids or dextrin have been tested for intravenous administration [5][51]. The addition of hydrophilic groups considerably improves the solubility of curcumin [30][31][32][33][75,76,77,78]. To make curcumin more relevant for bladder cancer treatment, intravenous infusion or intravesical instillations have been tested for the prevention of recurrence after tumor resection or BCG-therapy [32][33][77,78]. Other solutions enhancing curcumin absorption consider metal complexes, magnetic microspheres, or solid-lipid nanoparticles, which can be delivered straight to the targeted organs [29][30][31][74,75,76]. Published clinical trials present that complexing cyclodextrin with curcumin significantly improves intestinal absorption of curcuminoids [30][33][75,78]. Such commercial preparations are now available (brand name: Meriva, Curarti) [34][35][79,80].
Another field of clinical application is the synergistic activity of curcumin with existing anticancer agents. Co-administration of curcumin and paclitaxel significantly reduced expression of metalloproteinase-2, and decreased paclitaxel side effects in a PC3 xenografted prostate cancer model. In effect, such a formulation is proposed for hormone-refractory prostate cancer (HRPC) [9][55]. The synergistic action of gemcitabine plus curcumin has been presented in in vivo and in vitro models of pancreatic cancer [36][81]. In pancreatic cancer cell lines, inhibition of proliferation and apoptosis were substantially increased. Mice models of pancreatic cancer presented a significant reduction of tumor volume, suppression of NF-κB regulated genes (for cyclin D1, Bcl-2, BclxL, COX-2, matrix metalloproteinases, VEGF), and decreased microvessel density (Figure 1) [36][81].
Structural modifications of the curcumin molecule may facilitate its bioavailability, and increase treatment potential. New chemical analogues of curcumin form a very promising group of anticancer drugs. An analog FLLL32 blocks binding of transcription factors to DNA; induces its degradation; decreases VEGF, MMP-2, survivin expression; and promotes apoptosis in an experimental human osteosarcoma cell line [37][82]. Another analog is formulated by altering aromatic moiety to increase water solubility, metal binding, and MMP inhibition [4][50]. Contemporarily, a few dozen curcumin derivatives and dibenzoyl analogs have been tested for possible antioxidant, antiproliferative, and anti-inflammatory efficacy [4][50]. Hydrazinocurcumin, another synthetic curcumin derivative, significantly inhibited expression of MMP-2 and -9 by inhibition of the STAT3 signaling pathway as presented in breast cancer cell lines [9][55]. A very promising modification of curcumin aims at improving the binding of zinc ions. A novel MMP inhibitory formulation combines tetracyclines with curcumin [4][50]. Tetracycline molecules contain diketonic moiety, which binds zinc ions, and thus, may inhibit zinc-containing MMPs irrelevantly to antibacterial properties [8][54]. Similar metal-binding moieties contain curcumin molecules, and thus, complexing tetracycline with curcumin will combine and increase their inhibitory effect on MMPs by the zinc-binding properties of both being formulated into a new compound [8][10][54,56].
2. The Rationale for Curcumin Application in Bladder Cancer as a Potential Factor Limiting the Progression of the Disease
The characteristic clinical feature of bladder cancer is its propensity to recur and progress. Approximately three-quarters of cases are initially confined to the urothelium, not crossing the BM, as non-muscle-invasive cancer (NMIBC). The remaining one-quarter of cases manifest as muscle-invasive neoplasm (MIBC). A five-years recurrence rate of NMIBC reaches 70%, and one-third of them will progress to invasive MIBC. At the stage of invasive disease, the prognosis is much worse, as 15% patients will develop metastases [4][38][39][50,83,84]. The response rate for combined chemotherapy such as MVAC (methotrexate + vinblastine + doxorubicin + cisplatin) reaches up to 20%; however, 60% of patients do not respond to the treatment [5][51].
Surgical treatment of NMIBC is complemented by intravesical immunotherapy or chemotherapy with attenuated tuberculosis mycobacteria (BCG) or cytostatics (mitomycin), respectively. The aim of this treatment is to prevent the disease from becoming the muscle-invasive form. Immunotherapy and chemotherapy show efficacy, but a significant percentage of patients do not respond to such treatment, and develop severe adverse reactions. Therefore, research is targeted at the development of innovative high-potential medications replacing or enhancing the above adjuvant therapies to significantly improve the effectiveness of treatment [4][40][50,85]. Unfortunately, existing and innovative systemic and intravesical drugs are characterized by a high rate of side effects, along with a failure to achieve expected outcomes. Over 30% of patients do not respond to intravesical BG treatment [40][85]. Intravesical chemotherapy (mitomycin, epirubicin, thiotepa) causes local and systemic toxicity, whereas its actual therapeutic efficacy is limited [39][84]. Up to 50% of patients respond to adjuvant chemotherapy, but almost all are affected by its toxicity [38][40][83,85].
Natural plant-derived products have emerged as valuable alternatives with proven nontoxicity and suitability for the prevention and treatment of a number of ailments. Yet, only recently, meticulous research has revealed the complexity of the molecular mechanisms of action of commonly used nutraceuticals. Curcumin is an extensively tested nutrient for its proven, virtually unlimited therapeutic qualities. Its beneficial effects have been shown for a number of human malignancies during several clinical trials. A small series of tests have been carried out on bladder cancer. As bladder cancer therapy is the most capital-intensive among all cancer therapies, it is reasonable to reach for inexpensive, but effective, substances complementing and amending surgery—still accepted as the contemporary essential form of the treatment [41][42][86,87]. Existing clinical and laboratory experiments present multidirectional anticarcinogenic activities of curcumin on bladder cancer as monotherapy, or synergistically potentiating other chemotherapeutics. Moreover, experimental data presents curcumin bioactivity on all stages of carcinogenesis both intra- and extracellularly at concentrations of at least 40 μmol/L [41][86]. A number of experiments were conducted in vitro on experimental cell lines to explain intracellular mechanisms of curcumin activity [40][85]. Yet, despite having obtained many data, the exact mechanisms by which curcumin executes observed effects have not been fully explained [41][42][86,87].
Clonal assays presented and proved that curcumin is lethal to BC cell lines inducing apoptosis and arrest of the cell cycle in both G1/S and G2/M phases [40][43][85,88]. Curcumin also downregulated expression of essential antiapoptotic proteins (Bcl-2, Survivin, NF-κB) in parallel with upregulation of proapoptotic mediators (Bax, p53, caspase 3). These effects are stronger than those caused by cisplatin alone [43][44][45][88,89,90]. The apoptotic effect of gemcitabine and paclitaxel was intensified when co-administered with curcumin [36][40][44][81,85,89]. Concurrently, curcumin exhibits chemopreventive properties, due to in-vitro-presented inhibition of intracellular pathways activating external chemical carcinogens [39][84].
Still, intravesical BCG immunotherapy remains the most effective method of treatment of bladder cancer. Prevention of tumor recurrence and of transition from non-muscle invasive to invasive tumor is the main goal of such therapy. Several in vitro and in vivo studies presented prominent synergistic effects of co-administration of BCG and curcumin [32][33][77,78]. This beneficial phenomenon is multidirectional through influence on different cellular signaling molecules. Curcumin potentiates proapoptotic effects of the tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) by upregulation of its main DR5 membrane receptor [46][47][91,92]. Because BCG acts mainly through stimulation of the expression of TRIAL by neutrophils, co-administration with curcumin will enhance and intensify the BCG-induced immune response in bladder cancer [32][33][46][77,78,91]. On experimental cancer cells lines, the percentage of apoptotic cells ranged from 43% to 74% for BCG alone and BCG+curcumin, respectively [46][91]. The ineffectiveness of BCG-therapy observed in a subset of patients can be explained by the TRAIL resistance of tumor cells, related to constitutive (over)activation of nuclear factor NF-κB [46][91]. As curcumin suppresses NF-κB and further expression of dependent genes, its addition to BCG can reverse this resistance, and potentiate apoptosis [47][92].
Another pathway contributing to BCG-therapy failure is the overexpression of proapoptotic protein Bcl-2, which inhibits TRAIL-induced apoptosis. As described, curcumin effectively inhibits Bcl-2 and enhances tumor cells death [48][93]. In vitro assays proved that expression of proliferation marker cyclin D1 decreased by 50% after treatment with BCG alone, and up to 90% concomitantly with curcumin [46][91]. A proapoptotic response was additionally supported, and intensified by the formation of reactive oxygen species under curcumin influence [40][41][85,86]. Several other cellular pathways are modulated by curcumin, such as an inhibition of nitric oxide synthase, tyrosine kinases, transcriptional factors c- jun/AP-1, arachidonic acid pathways, COX activity, and many more. Wnt/Beta-catenin signal transduction pathways are also targeted by curcumin by downregulation of catenin, thus, affecting crucial metastasis EMT induction in bladder cancer cells [41][86]. This activity overlaps with the inhibition of MMP-2 and -9 expression, also involved in EMT processes. Indeed, the expression of mesenchymal markers (vimentin, N-cadherin) decreases in the presence of curcumin in a dose-dependent manner, whereas the expression of epithelial differentiation markers (E-cadherin) increases. This indicates that curcumin suspends bladder cancer cells in an epithelial, polarized phenotype; and restricts ″mobile″ mesenchymal features, hampering cancer cell migration, bladder cancer invasiveness, and further metastasis [49][94]. Curcumin also suppresses beta-catenin overexpression in bladder cancer cells, and thus, reverses metastatic potential and migration of bladder cancer cells in a dose-dependent manner [41][42][86,87].
The above data and other published studies suggest that the suppression of the NF-κB pathway is most likely essential for the multidirectional activity of curcumin (anti-inflammatory, anti-proliferatory, anti-invasive, anti-angiogenic) [44][46][47][89,91,92]. This mechanism was depicted in a previous part of this paper, and was proved on experimental bladder cancer cell lines [44][89]. The evidence shows that several others pathways associated with PI3K/AKT/mTOR, ERK1/2-signaling, insulin receptor substrate-1, insulin-like growth factor-2, and trophoblast cell surface antigen-2 related to bladder carcinogenesis are also affected by curcumin [4][5][50,51]. Data from in vitro experiments are confirmed by in vivo studies. In animal models of human bladder cancer, both xenografts and chemically induced curcumin inhibited cancer cell implantation, tumor growth, and metastasis [41][86]. These effects were observed after gavage, and after intravesical instillation. To overcome poor intestinal absorption, alternative modes of application have been tested. In line with intravesical administration, intraperitoneal and intravenous injections proved to be systematically effective and safe in a rat bladder cancer model [39][84]. All above studies show that curcumin is an effective modality in the prevention of NMIBC bladder cancer recurrence and progression, both as a sole agent and as a synergistic additive chemosensitizer for existing therapies [4][5][50,51].
Intravesical instillations of curcumin in experimental mouse bladder cancer models resulted in tumor necrosis and a significant reduction of tumor size, but with no effect on the number of tumors per bladder [44][89]. The combination of curcumin and BCG appeared to be more effective than BCG or curcumin alone [46][91]. In addition to the impact on existing tumors, curcumin inhibited implantation of free cancer cells in a bladder cancer murine model through the influence on integrin adhesion receptors [50][51][95,96]. A beneficial additive effect has been presented for intravesical or intratumoral or oral administration of curcumin concurrent with standard intravesical administration of BCG, with doses up to 8000 mg/day [46][91]. Even though only a few such studies have been published, evidences suggest a high therapeutic capacity of curcumin in bladder cancer, and place curcumin as possible integral part of therapy [4][5][50,51]. The synergy of BCG and curcumin is reflected in cellular molecular events, such as in a decrease of cell proliferation proteins (Ki67, cyclin D1, c-myc), anti-apoptotic proteins (Bcl-2, Bcl-xl, survivin), and pro-angiogenic proteins (CD31, VEGF), and the inhibition of epigenetic controllers (HDAC) [5][51].
Curcumin also potentiates therapeutic effects, and alleviates side effects of bladder cancer adjuvant chemotherapy. It increases therapeutic efficiency, and reverses tumor resistance to gemcitabine [52][97]. The combination of oral curcumin with intraperitoneal cisplatin resulted in an increased therapeutic efficiency of chemotherapy, and an increased reduction of tumor size in mouse models [53][98]. This synergistic effect can be explained by the induction of reactive oxygen species-related activation of proapoptotic pathways, and the inhibition of antiapoptotic pathways [53][98]. Concomitantly, curcumin alleviates cisplatin-related nephrotoxicity.
Acute kidney injury is the most common severe side effect of cisplatin-based therapy. Nephrotoxicity is related to a series of dysfunctional cellular metabolic pathways which pathologically intensify renal inflammatory processes, along with devastating oxidative stress, necrosis, and apoptosis of proximal tubular epithelial cells (PTEC) [5][54][55][56][51,99,100,101]. Cisplatin directly and/or indirectly induces upregulation of key molecules involved in those pathways, among others: TNF-α, p53, Fas ligand/receptor system, COX-2, caspases, and nuclear transcription factor-kappa B (NF-κB). These pathways are interlinked through common molecules [54][55][56][99,100,101]. Overexpression of TNF-α activates humoral and cellular inflammatory processes, and induces the generation of reactive oxygen species, leading to renal damage. P53 activates Bax-related proapoptotic pathways, and impairs Bcl-2- and Bcl-xL-related antiapoptotic pathways. Also, the Fas ligand/receptor apoptotic system is pathologically induced by cisplatin-upregulated p53 [54][55][56][99,100,101]. Cisplatin-related activation of transcription factor NF-κB, together with upregulated p53, stimulates pathways suppressing nephroprotective cytokine HNF1β, and also through the promotion of TNF-α synthesis [54][56][99,101]. Such interactions induce renal tubular cells apoptosis and necrosis, resulting in acute kidney injury. Further, epithelial damage attracts an influx of immunocompetent cells, with subsequent aggressive renal damage. Studies revealed that selective suppression of the above molecules alleviates cisplatin-related tubular epithelium dysfunction and renal injury. Existing nephroprotective strategies (cimetidine, mannitol, amifostine, celecoxib, etc.) are not always effective [5][54][55][51,99,100]. Curcumin directly and indirectly blocks the aforementioned molecules and involved pathways (see Figure 1). Laboratory experiments clearly present that curcumin reduced acute kidney injury in mice, and downregulated the pro-apoptotic cisplatin-related response in renal tubular cells [54][56][99,101]. This curcumin-associated reno-protective effect is achieved precisely by targeting key multifunctional cytokines, such as p53, NF-κB, Bax, and the others mentioned above. Therefore, curcumin appears to be an important renoprotective complementary and supportive agent for cancer chemotherapy [54][99].
Immunologic escape of the tumor has also been modulated by curcumin through inhibition of expression of programmed cell death ligand 1 (PDL1) on both bladder cancer cells and tumor-infiltrating lymphocytes, as proved by in vivo and in vitro experiments [5][51]. Also, a clinical trial presented that curcumin intensified patients’ immunologic response by stimulation of interferon-gamma production, and the propagation of T- helper lymphocytes and cytotoxic NK cells [5][51].
To fully exploit anti-PDL1 activity of curcumin, a few studies have been conducted. The combined application of curcumin molecules and anti-PDL1 antibodies was tested in a bladder tumor mouse model [5][51]. This construct targets the programmed cell death protein (PD-1) receptor of lymphocytes. One clinical trial on patients with gynecological carcinomas combining pembrolizumab (clinically approved anti-PD1 receptor antibody) with RTG therapy and curcumin food supplementation (with additional vit. D, lansoprazole, aspirin, cyclophosphamide) was conducted to elevate the proportion of response for a PD-1-blockade + radiotherapy treatment regime [57][102].
References
- Knecht, K.; Kinder, D.; Stockert, A. Biologically-based complementary and alternative medicine (CAM) use in cancer patients: The good, the bad, the misunderstood. Front. Nutr. 2020, 6, 196. References
- Guldiken, B.; Ozkan, G.; Catalkaya, G.; Ceylan, F.D.; Ekin Yalcinkaya, I.; Capanoglu, E. Phytochemicals of herbs and spices: Health versus toxicological effects. Food Chem. Toxicol. 2018, 119, 37–49. Kirkali, Z.; Chan, T.; Manoharan, M.; Algaba, F.; Busch, C.; Cheng, L.; Kiemeney, L.; Kriegmair, M.; Montironi, R.; Murphy, W.M.; et al. Bladder cancer: Epidemiology, staging and grading, and diagnosis. Urology 2005, 66 (Suppl. 1), 4–34. [Google Scholar] [CrossRef]
- Jobin, C.; Bradham, C.A.; Russo, M.P.; Juma, B.; Narula, A.S.; Brenner, D.A.; Sartor, R.B. Curcumin blocks cytokine- mediated NF-kappa B activation and proinflammatory gene expression by inhibiting inhibitory factor I-kappa B kinase activity. J. Immunol. 1999, 163, 3474–3483. Yuk, H.D.; Ku, J.H. Role of Systemic Inflammatory Response Markers in Urothelial Carcinoma. Front. Oncol. 2020, 10, 1473. [Google Scholar] [CrossRef]
- Hauser, P.J.; Han, Z.; Sindhwani, P.; Hurst, R.E. Sensitivity of bladder cancer cells to curcumin and its derivatives depends on the extracellular matrix. Anticancer Res. 2007, 27, 37–40. Maloney, I.; Parker, D.C.; Cookson, M.S.; Patel, S. Bladder cancer recovery pathways: A systematic review. Bladder Cancer 2017, 4, 269–281. [Google Scholar] [CrossRef]
- Rutz, J.; Janicova, A.; Woidacki, K.; Chun, F.K.; Blaheta, R.A.; Relja, B. Curcumin-A viable agent for better bladder cancer treatment. Int. J. Mol. Sci. 2020, 21, 3761. Richters, A.; Aben, K.K.H.; Kiemeney, L.A.L.M. The global burden of urinary bladder cancer: An update. World J. Urol. 2020, 38, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Chainani-Wu, N. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcuma longa). J. Altern. Complement. Med. 2003, 9, 161–168. Avritscher, E.B.C.; Cooksley, C.D.; Grossman, H.B.; Sabichi, A.L.; Hamblin, L.; Dinney, C.P.; Elting, L.S. Clinical model of lifetime cost of treating bladder cancer and associated complications. Urology 2006, 68, 3549–3553. [Google Scholar] [CrossRef]
- Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. Bolla, S.R.; Odeluga, N.; Jetti, R. Histology, Bladder. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK540963/ (accessed on 23 February 2021).
- Salehi, B.; Zucca, P.; Sharifi-Rad, M.; Pezzani, R.; Rajabi, S.; Setzer, W.N.; Varoni, E.M.; Iriti, M.; Kobarfard, F.; Sharifi-Rad, J. Phytotherapeutics in cancer invasion and metastasis. Phytother. Res. 2018, 32, 1425–1449. Babaian, K.N.; Adams, P.G.; McClure, C.; Tompkins, B.; McMurray, M. Bladder Cancer. Medscape. Updated 23 February 2021. Available online: https://emedicine.medscape.com/article/438262-overview (accessed on 23 February 2021).
- Kumar, D.; Kumar, M.; Saravanan, C.; Singh, S.K. Curcumin: A potential candidate for matrix metalloproteinase inhibitors. Expert Opin. Ther. Targets 2012, 16, 959–972. Marcos-Gragera, R.; Mallone, S.; Kiemeney, L.A.; Vilardell, L.; Malats, N.; Allory, Y.; Sant, M. EUROCARE-5 Working Group: Urinary tract cancer survival in Europe 1999–2007: Results of the population-based study EUROCARE-5. Eur. J. Cancer 2015, 15, 2217–2230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gu, Y.; Lee, H.M.; Hambardjieva, E.; Vranková, K.; Golub, L.M.; Johnson, F. Design, synthesis and biological activity of new polyenolic inhibitors of matrix metalloproteinases: A focus on chemically-modified curcumins. Curr. Med. Chem. 2012, 19, 4348–4358. Plachot, C.; Lelievre, S.A. Novel directions in tumour biology: From basement membrane-directed polarity to DNA methylation. In Cancer Modelling and Simulation, 1st ed.; Preziosi, L., Ed.; CRC Press: London, UK, 2003; ISBN 9781584883616. [Google Scholar]
- Swarnakar, S.; Ganguly, K.; Kundu, P.; Banerjee, A.; Maity, P.; Sharma, A.V. Curcumin regulates expression and activity of matrix metalloproteinases 9 and 2 during prevention and healing of indomethacin-induced gastric ulcer. J. Biol. Chem. 2005, 280, 9409–9415. Pozzi, A.; Yurchenco, P.D.; Iozzo, R.V. The nature and biology of basement membranes. Matrix Biol. 2017, 57–58, 1–11. [Google Scholar] [CrossRef]
- Bachmeier, B.E.; Killian, P.H.; Melchart, D. The role of curcumin in prevention and management of metastatic disease. Int. J. Mol. Sci. 2018, 19, 1716. Randles, M.J.; Humphries, M.J.; Lennon, R. Proteomic definitions of basement membrane composition in health and disease. Matrix Biol. 2017, 57–58, 12–28. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin. J. Med. Chem. 2017, 60, 1620–1637. Jayadev, R.; Sherwood, D.R. Basement membranes. Curr. Biol. 2017, 27, R207–R211. [Google Scholar] [CrossRef]
- Lampe, V.; Miłobędzka, J.; Kostanecki, S.V. Zur Kenntnis des Curcumins. Ber. Der Dtsch. Chem. Ges. 1910, 43, 2163–2170. Liotta, L.A.; Rao, C.N.; Wewer, U.M. Biochemical interactions of tumor cells with the basement membrane. Annu. Rev. Biochem. 1986, 55, 1037–1057. [Google Scholar] [CrossRef]
- Shen, L.; Ji, H.F. Theoretical study on physicochemical properties of curcumin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2007, 67, 619–623. Sekiguchi, R.; Yamada, K.M. Basement membranes in development and disease. Curr. Top. Dev. Biol. 2018, 130, 143–191. [Google Scholar] [CrossRef] [PubMed]
- Yodkeeree, S.; Chaiwangyen, W.; Garbisa, S.; Limtrakul, P. Curcumin, demethoxycurcumin, and bisdemethoxycurcumin differentially inhibit cancer cell invasion through the down-regulation of MMPs and uPA. J. Nutr. Biochem. 2009, 20, 87–95. Kalluri, R. Basement membranes: Structure, assembly and role in tumour angiogenesis. Nat. Rev. Cancer 2003, 3, 422–433. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Chattopadhyay, N.; Mitra, A.; Siddiqi, M.; Chatterjee, A. Curcumin exhibits antimetastatic properties by modulating integrin receptors, collagenase activity, and expression of Nm23 and E-cadherin. J. Environ. Pathol. Toxicol. Oncol. 2003, 22, 49–58. Marchisio, P.C.; Trusolino, L.; De Luca, M. Topography and biological role of integrins in human skin. Microsc. Res. Tech. 1997, 38, 353–360. [Google Scholar] [CrossRef]
- Mohan, R.; Sivak, J.; Ashton, P.; Russo, L.A.; Pham, B.Q.; Kasahara, N.; Raizman, M.B.; Fini, M.E. Curcuminoids inhibit the angiogenic response stimulated by fibroblast growth factor-2, including expression of matrix metalloproteinase gelatinase B. J. Biol. Chem. 2000, 275, 10405–10412. Wiseman, B.S.; Werb, Z. Stromal effects on mammary gland development and breast cancer. Science 2002, 296, 1046–1049. [Google Scholar] [CrossRef] [PubMed]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. Hamano, Y.; Zeisberg, M.; Sugimoto, H.; Lively, J.C.; Maeshima, Y.; Yang, C.; Hynes, R.O.; Werb, Z.; Sudhakar, A.; Kalluri, R. Physiological levels of tumstatin, a fragment of collagen IV α3 chain, are generated by MMP-9 proteolysis and suppress angiogenesis via αVβ3 integrin. Cancer Cell 2003, 3, 589–601. [Google Scholar] [CrossRef]
- Philip, S.; Kundu, G.C. Osteopontin induces nuclear factor kappa B-mediated promatrix metalloproteinase-2 activation through I kappa B alpha/IKK signaling pathways, and curcumin (diferulolylmethane) down-regulates these pathways. J. Biol. Chem. 2003, 278, 14487–14497. Horejs, C.M.; Serio, A.; Purvis, A.; Gormley, A.J.; Bertazzo, S.; Poliniewicz, A.; Wang, A.J.; Di Maggio, P.; Hohenester, E.; Stevens, M.M. Biologically-active laminin-111 fragment that modulates the epithelial-to-mesenchymal transition in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 5908–5913. [Google Scholar] [CrossRef]
- Shishodia, S.; Potdar, P.; Gairola, C.G.; Aggarwal, B.B. Curcumin (diferuloylmethane) down-regulates cigarette smoke-induced NF-kappaB activation through inhibition of IkappaBalpha kinase in human lung epithelial cells: Correlation with suppression of COX-2, MMP-9 and cyclin D1. Carcinogenesis 2003, 24, 1269–1279. Chiarugi, P.; Giannoni, E. Anoikis: A necessary death program for anchorage-dependent cells. Biochem. Pharmacol. 2008, 76, 1352–1364. [Google Scholar] [CrossRef]
- Matsuo, M.; Sakurai, H.; Koizumi, K.; Saiki, I. Curcumin inhibits the formation of capillary-like tubes by rat lymphatic endothelial cells. Cancer Lett. 2007, 251, 288–295. Slade, M.J.; Coope, R.C.; Gomm, J.J.; Coombes, R.C. The human mammary gland basement membrane is integral to the polarity of luminal epithelial cells. Exp. Cell Res. 1999, 247, 267–278. [Google Scholar] [CrossRef]
- Zou, J.; Lei, T.; Guo, P.; Yu, J.; Xu, Q.; Luo, Y.; Ke, R.; Huang, D. Mechanisms shaping the role of ERK1/2 in cellular senescence. Mol. Med. Rep. 2019, 2, 759–770. Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3, a004952. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Chakrabarti, J.; Banerji, A.; Chatterjee, A.; Das, B.R. Curcumin, a potential inhibitor of MMP-2 in human laryngeal squamous carcinoma cells HEp2. J. Environ. Pathol. Toxicol. Oncol. 2006, 25, 679–690. Clark, E.; Brugge, J. Integrins and signal transduction pathways: The road taken. Science 1995, 268, 233–239. [Google Scholar] [CrossRef]
- Xiao, C.J.; Yu, X.J.; Xie, J.L.; Liu, S.; Li, S. Protective effect and related mechanisms of curcumin in rat experimental periodontitis. Head Face Med. 2018, 14, 12. Bezakova, G.; Ruegg, M.A. New insights into the roles of agrin. Nat. Rev. Mol. Cell Biol. 2003, 4, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.L.; Iragavarapu-Charyulu, V.; Lopez, D.M. T cell-derived matrix metalloproteinase-9 in breast cancer: Friend or foe? Breast Dis. 2004, 20, 145–153. Leitinger, B. Discoidin domain receptor functions in physiological and pathological conditions. Int. Rev. Cell Mol. Biol. 2014, 310, 39–87. [Google Scholar] [CrossRef]
- Cao, J.; Han, Z.; Tian, L.; Chen, K.; Fan, Y.; Ye, B.; Huang, W.; Wang, C.; Huang, Z. Curcumin inhibits EMMPRIN and MMP-9 expression through AMPK-MAPK and PKC signaling in PMA induced macrophages. J. Transl. Med. 2014, 12, 266. O’Connor, K.L.; Shaw, L.M.; Mercurio, A.M. Release of cAMP gating by the alpha 6 beta 4 integrin stimulates lamellae formation and the chemotactic migration of invasive carcinoma cells. J. Cell Biol. 1998, 143, 1749–1760. [Google Scholar] [CrossRef]
- Hsu, C.H.; Cheng, A.L. Clinical studies with curcumin. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease. Advances in Experimental Medicine and Biology; Aggarwal, B.B., Surh, Y.J., Shishodia, S., Eds.; Springer: Boston, MA, USA, 2007; Volume 595. Rabinovitz, I.; Toker, A.; Mercurio, A.M. Protein kinase C-dependent mobilization of the alpha 6 beta 4 integrin 6 integrin from hemidesmosomes and its association with actin-rich cell protrusions drive the chemotactic migration of carcinoma cells. J. Cell Biol. 1999, 146, 1147–1160. [Google Scholar] [CrossRef] [PubMed]
- Patil, V.M.; Das, S.; Balasubramanian, K. Quantum chemical and docking insights into bioavailability enhancement of curcumin by piperine in pepper. J. Phys. Chem. A 2016, 120, 3643–3653. Nguyen-Ngoc, K.V.; Cheung, K.J.; Brenot, A.; Shamir, E.R.; Gray, R.S.; Hines, W.C.; Yaswen, P.; Werb, Z.; Ewald, A.J. ECM microenvironment regulates collective migration and local dissemination in normal and malignant mammary epithelium. Proc. Natl. Acad. Sci. USA 2012, 109, E2595–E2604. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. Nakamura, K.; Mori, M.; Enjoji, M. Distribution of basement membrane antigens in clinical gastric adenocarcinomas: An immunohistochemical study. J. Clin. Pathol. 1987, 40, 1418–1423. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Sen, R.; Paul, B.; Kazi, J.; Ganguly, S.; Debnath, M.C. Gemcitabine co-encapsulated with curcumin in folate decorated PLGA nanoparticles; a novel approach to treat breast adenocarcinoma. Pharm. Res. 2020, 37, 1–19. Ewald, A.J.; Huebner, R.J.; Palsdottir, H.; Lee, J.K.; Perez, M.J.; Jorgens, D.M.; Tauscher, A.N.; Cheung, K.J.; Werb, Z.; Auer, M. Mammary collective cell migration involves transient loss of epithelial features and individual cell migration within the epithelium. J. Cell Sci. 2012, 125, 2638–2654. [Google Scholar] [CrossRef] [PubMed]
- Redelman-Sidi, G.; Glickman, M.S.; Bochner, B.H. The mechanism of action of BCG therapy for bladder cancer-a current perspective. Nat. Rev. Urol. 2014, 11, 153–162. Tanos, B.; Rodriguez-Boulan, E. The epithelial polarity program: Machineries involved and their hijacking by cancer. Oncogene 2008, 27, 6939–6957. [Google Scholar] [CrossRef]
- Falke, J.; Parkkinen, J.; Vaahtera, L.; Hulsbergen-van de Kaa, C.A.; Oosterwijk, E.; Witjes, J.A. Curcumin as treatment for bladder cancer: A preclinical study of cyclodextrin-curcumin complex and BCG as intravesical treatment in an orthotopic bladder cancer rat model. Biomed. Res. Int. 2018, 2018, 9634902. Yoshizaki, K.; Yamada, Y. Gene evolution and functions of extracellular matrix proteins in teeth. Orthod. Waves 2013, 72, 1–10. [Google Scholar] [CrossRef]
- Xiang, D.-B.; Zhang, K.-Q.; Zeng, Y.-L.; Yan, Q.-Z.; Shi, Z.; Tuo, Q.-H.; Lin, L.-M.; Xia, B.-H.; Wu, P.; Liao, D.-F. Curcumin: From a controversial “panacea” to effective antineoplastic products. Medicine 2020, 99, e18467. Felbor, U.; Dreier, L.; Bryant, R.A.; Ploegh, H.L.; Olsen, B.R.; Mothes, W. Secreted cathepsin L generates endostatin from collagen XVIII. EMBO J. 2000, 19, 1187–1194. [Google Scholar] [CrossRef]
- Purpura, M.; Lowery, R.P.; Wilson, J.M.; Mannan, H.; Münch, G.; Razmovski-Naumovski, V. Analysis of different innovative formulations of curcumin for improved relative oral bioavailability in human subjects. Eur. J. Nutr. 2018, 57, 929–938. Swarnakar, S.; Ganguly, K.; Kundu, P.; Banerjee, A.; Maity, P.; Sharma, A.V. Curcumin regulates expression and activity of matrix metalloproteinases 9 and 2 during prevention and healing of indomethacin-induced gastric ulcer. J. Biol. Chem. 2005, 280, 9409–9415. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Guha, S.; Krishnan, S.; Diagaradjane, P.; Gelovani, J.; Aggarwal, B.B. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res. 2007, 67, 3853–3861. Hayakawa, T.; Yamashita, K.; Ohuchi, E.; Shinagawa, A. Cell growth- promoting activity of tissue inhibitor of metalloproteinases-2 (TIMP-2). J. Cell Sci. 1994, 107, 2373–2379. [Google Scholar] [CrossRef]
- Fossey, S.L.; Bear, M.D.; Lin, J.; Li, C.; Schwartz, E.B.; Li, P.K.; Fuchs, J.R.; Fenger, J.; Kisseberth, W.C.; London, C.A. The novel curcumin analog FLLL32 decreases STAT3 DNA binding activity and expression, and induces apoptosis in osteosarcoma cell lines. BMC Cancer 2011, 11, 112. Mueller, S.C.; Ghersi, G.; Akiyama, S.K.; Sang, Q.X.; Howard, L.; Pineiro-Sanchez, M.; Nakahara, H.; Yeh, Y.; Chen, W.T. A novel protease-docking function of integrin at invadopodia. J. Biol. Chem. 1999, 274, 24947–24952. [Google Scholar] [CrossRef] [PubMed]
- Mari, A.; D’Andrea, D.; Abufaraj, M.; Foerster, B.; Kimura, S.; Shariat, S.F. Genetic determinants for chemo- and radiotherapy resistance in bladder cancer. Transl. Androl. Urol. 2017, 6, 1081–1089. Rebustini, I.T.; Myers, C.; Lassiter, K.S.; Surmak, A.; Szabova, L.; Holmbeck, K.; Pedchenko, V.; Hudson, B.G.; Hoffman, M.P. MT2-MMP-dependent release of collagen IV NC1 domains regulates submandibular gland branching morphogenesis. Dev. Cell 2009, 17, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Amling, C.L. Diagnosis and management of superficial bladder cancer. Curr. Probl. Cancer 2001, 4, 219–278. O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.; Folkman, J. Endostatin: An endogenous inhibitor of angiogenesis and tumor growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef]
- Herr, H.W.; Wartinger, D.D.; Fair, W.R.; Oettgen, H.F. Bacillus Calmette-Guerin therapy for superficial bladder cancer: A 10-year followup. J. Urol. 1992, 147, 1020–1023. Ortega, N.; Werb, Z. New functional roles for non-collagenous domains of basement membrane collagens. J. Cell Sci. 2002, 115, 4201–4214. [Google Scholar] [CrossRef]
- Tian, B.; Wang, Z.; Zhao, Y.; Wang, D.; Li, Y.; Ma, L.; Li, X.; Li, J.; Xiao, N.; Tian, J.; et al. Effects of curcumin on bladder cancer cells and development of urothelial tumors in a rat bladder carcinogenesis model. Cancer Lett. 2008, 264, 299–308. Madsen, D.H.; Bugge, T.H. The source of matrix-degrading enzymes in human cancer: Problems of research reproducibility and possible solutions. J. Cell Biol. 2015, 209, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, Y.; Jia, Z.; Gao, Y.; Zhao, C.; Yao, Y. Curcumin inhibits bladder cancer progression via regulation of β-catenin expression. Tumor Biol. 2017, 39, 1010428317702548. Nielsen, B.S.; Sehested, M.; Duun, S.; Rank, F.; Timshel, S.; Rygaard, J.; Johnsen, M.; Dano, K. Urokinase plasminogen activator is localized in stromal cells in ductal breast cancer. Lab. Investig. 2001, 81, 1485–1501. [Google Scholar] [CrossRef]
- Botteman, M.F.; Pashos, C.L.; Redaelli, A.; Laskin, B.; Hauser, R. The health economics of bladder cancer: A comprehensive review of the published literature. Pharmacoeconomics 2003, 21, 1315–1330. Grindel, B.; Li, Q.; Arnold, R.; Petros, J.; Zayzafoon, M.; Muldoon, M.; Stave, J.; Chung, L.W.; Farach-Carson, M.C. Perlecan/HSPG2 and matrilysin/MMP-7 as indices of tissue invasion: Tissue localization and circulating perlecan fragments in a cohort of 288 radical prostatectomy patients. Oncotarget 2016, 7, 10433–10447. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.J.; Deng, N.; Zou, Z.H.; Chen, G.X. The effect of curcumin on bladder tumor in rat model. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 884–889. Farach-Carson, M.C.; Brown, A.C.; Lynam, M.; Safran, J.B.; Carson, D.D. A novel peptide sequence in perlecan domain IV supports cell adhesion, spreading and FAK activation. Matrix Biol. 2008, 27, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Chendil, D.; Ranga, R.S.; Meigooni, D.; Sathishkumar, S.; Ahmed, M.M. Curcumin confers radiosensitizing effect in prostate cancer cell line PC-3. Oncogene 2004, 23, 1599–1607. Mongiat, M.; Sweeney, S.; San Antonio, J.D.; Fu, J.; Iozzo, R.V. Endorepellin, a novel inhibitor of angiogenesis derived from the C terminus of perlecan. J. Biol. Chem. 2003, 278, 4238–4249. [Google Scholar] [CrossRef] [PubMed]
- Kamat, A.M.; Tharakan, S.T.; Sung, B.; Aggarwal, B.B. Curcumin potentiates the antitumor effects of Bacillus Calmette-Guerin against bladder cancer through the downregulation of NF-kappaB and upregulation of TRAIL receptors. Cancer Res. 2009, 69, 8958–8966, Erratum in Cancer Res. 2018, 78, 5182. Gonzalez, E.M.; Reed, C.C.; Bix, G.; Fu, J.; Zhang, Y.; Gopalakrishnan, B.; Greenspan, D.S.; Iozzo, R.V. BMP-1/Tolloid-like metalloproteases process endorepellin, the angiostatic C-terminal fragment of perlecan. J. Biol. Chem. 2005, 280, 7080–7087. [Google Scholar] [CrossRef] [PubMed]
- Khanbolooki, S.; Nawrocki, S.T.; Arumugam, T.; Andtbacka, R.; Pino, M.S.; Kurzrock, R.; Logsdon, C.D.; Abbruzzese, J.L.; McConkey, D.J. Nuclear factor-kappaB maintains TRAIL resistance in human pancreatic cancer cells. Mol. Cancer Ther. 2006, 9, 2251–2260. Passos-Bueno, M.R.; Suzuki, O.T.; Armelin-Correa, L.M.; Sertie, A.L.; Errera, F.I.; Bagatini, K.; Kok, F.; Leite, K.R. Mutations in collagen 18A1 and their relevance to the human phenotype. An. Acad. Bras. Cienc. 2006, 78, 123–131. [Google Scholar] [CrossRef]
- Kruyt, F.A. TRAIL and cancer therapy. Cancer Lett. 2008, 263, 14–25. Knecht, K.; Kinder, D.; Stockert, A. Biologically-based complementary and alternative medicine (CAM) use in cancer patients: The good, the bad, the misunderstood. Front. Nutr. 2020, 6, 196. [Google Scholar] [CrossRef]
- Duan, W.; Chang, Y.; Li, R.; Xu, Q.; Lei, J.; Yin, C.; Li, T.; Wu, Y.; Ma, Q.; Li, X. Curcumin inhibits hypoxia inducible factor-1α-induced epithelial-mesenchymal transition in HepG2 hepatocellular carcinoma cells. Mol. Med. Rep. 2014, 10, 2505–2510. Guldiken, B.; Ozkan, G.; Catalkaya, G.; Ceylan, F.D.; Ekin Yalcinkaya, I.; Capanoglu, E. Phytochemicals of herbs and spices: Health versus toxicological effects. Food Chem. Toxicol. 2018, 119, 37–49. [Google Scholar] [CrossRef]
- Sindhwani, P.; Hampton, J.A.; Baig, M.M.; Keck, R.; Selman, S.H. Curcumin prevents intravesical tumor implantation of the MBT-2 tumor cell line in C3H mice. J. Urol. 2001, 166, 1498–1501. Jobin, C.; Bradham, C.A.; Russo, M.P.; Juma, B.; Narula, A.S.; Brenner, D.A.; Sartor, R.B. Curcumin blocks cytokine- mediated NF-kappa B activation and proinflammatory gene expression by inhibiting inhibitory factor I-kappa B kinase activity. J. Immunol. 1999, 163, 3474–3483. [Google Scholar] [PubMed]
- Zhang, L.; Yang, G.; Zhang, R.; Dong, L.; Chen, H.; Bo, J.; Xue, W.; Huang, Y. Curcumin inhibits cell proliferation and motility via suppression of TROP2 in bladder cancer cells. Int. J. Oncol. 2018, 53, 515–526. Hauser, P.J.; Han, Z.; Sindhwani, P.; Hurst, R.E. Sensitivity of bladder cancer cells to curcumin and its derivatives depends on the extracellular matrix. Anticancer Res. 2007, 27, 37–40. [Google Scholar]
- Cho, C.-J.; Yang, C.-W.; Wu, C.-L.; Ho, J.-Y.; Yu, C.-P.; Wu, S.-T.; Yu, D.-S. The modulation study of multiple drug resistance in bladder cancer by curcumin and resveratrol. Oncol. Lett. 2019, 18, 6869–6876. Rutz, J.; Janicova, A.; Woidacki, K.; Chun, F.K.; Blaheta, R.A.; Relja, B. Curcumin-A viable agent for better bladder cancer treatment. Int. J. Mol. Sci. 2020, 21, 3761. [Google Scholar] [CrossRef]
- Park, B.H.; Lim, J.E.; Jeon, H.G.; Seo, S.I.; Lee, H.M.; Choi, H.Y.; Jeon, S.S.; Jeong, B.C. Curcumin potentiates antitumor activity of cisplatin in bladder cancer cell lines via ROS-mediated activation of ERK1/2. Oncotarget 2016, 7, 63870–63886. Chainani-Wu, N. Safety and anti-inflammatory activity of curcumin: A component of tumeric (Curcuma longa). J. Altern. Complement. Med. 2003, 9, 161–168. [Google Scholar] [CrossRef]
- dos santos, N.A.; Carvalho Rodrigues, M.A.; Martins, N.M.; dos Santos, A.C. Cisplatin-induced nephrotoxicity and targets of nephroprotection: An update. Arch. Toxicol. 2012, 86, 1233–1250. Kocaadam, B.; Sanlier, N. Curcumin, an active component of turmeric (Curcuma longa), and its effects on health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2889–2895. [Google Scholar] [CrossRef] [PubMed]
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. Salehi, B.; Zucca, P.; Sharifi-Rad, M.; Pezzani, R.; Rajabi, S.; Setzer, W.N.; Varoni, E.M.; Iriti, M.; Kobarfard, F.; Sharifi-Rad, J. Phytotherapeutics in cancer invasion and metastasis. Phytother. Res. 2018, 32, 1425–1449. [Google Scholar] [CrossRef]
- Volarevic, V.; Djokovic, B.; Jankovic, M.G.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N. Molecular mechanisms of cisplatin-induced nephrotoxicity: A balance on the knife edge between renoprotection and tumor toxicity. J. Biomed. Sci. 2019, 26, 25. Kumar, D.; Kumar, M.; Saravanan, C.; Singh, S.K. Curcumin: A potential candidate for matrix metalloproteinase inhibitors. Expert Opin. Ther. Targets 2012, 16, 959–972. [Google Scholar] [CrossRef] [PubMed]
- Tuyaerts, S.; van Nuffel, A.M.T.; Naert, E.; van Dam, P.A.; Vuylsteke, P.; de Caluwé, A.; Aspeslagh, S.; Dirix, P.; Lippens, L.; de Jaeghere, E.; et al. PRIMMO study protocol: A phase II study combining PD-1 blockade, radiation and immunomodulation to tackle cervical and uterine cancer. BMC Cancer 2019, 19, 506. Zhang, Y.; Gu, Y.; Lee, H.M.; Hambardjieva, E.; Vranková, K.; Golub, L.M.; Johnson, F. Design, synthesis and biological activity of new polyenolic inhibitors of matrix metalloproteinases: A focus on chemically-modified curcumins. Curr. Med. Chem. 2012, 19, 4348–4358. [Google Scholar] [CrossRef]
- Bachmeier, B.E.; Killian, P.H.; Melchart, D. The role of curcumin in prevention and management of metastatic disease. Int. J. Mol. Sci. 2018, 19, 1716. [Google Scholar] [CrossRef]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Lampe, V.; Miłobędzka, J.; Kostanecki, S.V. Zur Kenntnis des Curcumins. Ber. Der Dtsch. Chem. Ges. 1910, 43, 2163–2170. [Google Scholar]
- Shen, L.; Ji, H.F. Theoretical study on physicochemical properties of curcumin. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2007, 67, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Yodkeeree, S.; Chaiwangyen, W.; Garbisa, S.; Limtrakul, P. Curcumin, demethoxycurcumin, and bisdemethoxycurcumin differentially inhibit cancer cell invasion through the down-regulation of MMPs and uPA. J. Nutr. Biochem. 2009, 20, 87–95. [Google Scholar] [CrossRef]
- Ray, S.; Chattopadhyay, N.; Mitra, A.; Siddiqi, M.; Chatterjee, A. Curcumin exhibits antimetastatic properties by modulating integrin receptors, collagenase activity, and expression of Nm23 and E-cadherin. J. Environ. Pathol. Toxicol. Oncol. 2003, 22, 49–58. [Google Scholar]
- Mohan, R.; Sivak, J.; Ashton, P.; Russo, L.A.; Pham, B.Q.; Kasahara, N.; Raizman, M.B.; Fini, M.E. Curcuminoids inhibit the angiogenic response stimulated by fibroblast growth factor-2, including expression of matrix metalloproteinase gelatinase B. J. Biol. Chem. 2000, 275, 10405–10412. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Philip, S.; Kundu, G.C. Osteopontin induces nuclear factor kappa B-mediated promatrix metalloproteinase-2 activation through I kappa B alpha/IKK signaling pathways, and curcumin (diferulolylmethane) down-regulates these pathways. J. Biol. Chem. 2003, 278, 14487–14497. [Google Scholar] [CrossRef]
- Shishodia, S.; Potdar, P.; Gairola, C.G.; Aggarwal, B.B. Curcumin (diferuloylmethane) down-regulates cigarette smoke-induced NF-kappaB activation through inhibition of IkappaBalpha kinase in human lung epithelial cells: Correlation with suppression of COX-2, MMP-9 and cyclin D1. Carcinogenesis 2003, 24, 1269–1279. [Google Scholar] [CrossRef]
- Matsuo, M.; Sakurai, H.; Koizumi, K.; Saiki, I. Curcumin inhibits the formation of capillary-like tubes by rat lymphatic endothelial cells. Cancer Lett. 2007, 251, 288–295. [Google Scholar] [CrossRef]
- Zou, J.; Lei, T.; Guo, P.; Yu, J.; Xu, Q.; Luo, Y.; Ke, R.; Huang, D. Mechanisms shaping the role of ERK1/2 in cellular senescence. Mol. Med. Rep. 2019, 2, 759–770. [Google Scholar] [CrossRef]
- Mitra, A.; Chakrabarti, J.; Banerji, A.; Chatterjee, A.; Das, B.R. Curcumin, a potential inhibitor of MMP-2 in human laryngeal squamous carcinoma cells HEp2. J. Environ. Pathol. Toxicol. Oncol. 2006, 25, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.J.; Yu, X.J.; Xie, J.L.; Liu, S.; Li, S. Protective effect and related mechanisms of curcumin in rat experimental periodontitis. Head Face Med. 2018, 14, 12. [Google Scholar] [CrossRef]
- Owen, J.L.; Iragavarapu-Charyulu, V.; Lopez, D.M. T cell-derived matrix metalloproteinase-9 in breast cancer: Friend or foe? Breast Dis. 2004, 20, 145–153. [Google Scholar] [CrossRef]
- Cao, J.; Han, Z.; Tian, L.; Chen, K.; Fan, Y.; Ye, B.; Huang, W.; Wang, C.; Huang, Z. Curcumin inhibits EMMPRIN and MMP-9 expression through AMPK-MAPK and PKC signaling in PMA induced macrophages. J. Transl. Med. 2014, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.H.; Cheng, A.L. Clinical studies with curcumin. In The Molecular Targets and Therapeutic Uses of Curcumin in Health and Disease. Advances in Experimental Medicine and Biology; Aggarwal, B.B., Surh, Y.J., Shishodia, S., Eds.; Springer: Boston, MA, USA, 2007; Volume 595. [Google Scholar] [CrossRef]
- Patil, V.M.; Das, S.; Balasubramanian, K. Quantum chemical and docking insights into bioavailability enhancement of curcumin by piperine in pepper. J. Phys. Chem. A 2016, 120, 3643–3653. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Sen, R.; Paul, B.; Kazi, J.; Ganguly, S.; Debnath, M.C. Gemcitabine co-encapsulated with curcumin in folate decorated PLGA nanoparticles; a novel approach to treat breast adenocarcinoma. Pharm. Res. 2020, 37, 1–19. [Google Scholar] [CrossRef]
- Redelman-Sidi, G.; Glickman, M.S.; Bochner, B.H. The mechanism of action of BCG therapy for bladder cancer-a current perspective. Nat. Rev. Urol. 2014, 11, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Falke, J.; Parkkinen, J.; Vaahtera, L.; Hulsbergen-van de Kaa, C.A.; Oosterwijk, E.; Witjes, J.A. Curcumin as treatment for bladder cancer: A preclinical study of cyclodextrin-curcumin complex and BCG as intravesical treatment in an orthotopic bladder cancer rat model. Biomed. Res. Int. 2018, 2018, 9634902. [Google Scholar] [CrossRef]
- Xiang, D.-B.; Zhang, K.-Q.; Zeng, Y.-L.; Yan, Q.-Z.; Shi, Z.; Tuo, Q.-H.; Lin, L.-M.; Xia, B.-H.; Wu, P.; Liao, D.-F. Curcumin: From a controversial “panacea” to effective antineoplastic products. Medicine 2020, 99, e18467. [Google Scholar] [CrossRef] [PubMed]
- Purpura, M.; Lowery, R.P.; Wilson, J.M.; Mannan, H.; Münch, G.; Razmovski-Naumovski, V. Analysis of different innovative formulations of curcumin for improved relative oral bioavailability in human subjects. Eur. J. Nutr. 2018, 57, 929–938. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Guha, S.; Krishnan, S.; Diagaradjane, P.; Gelovani, J.; Aggarwal, B.B. Curcumin potentiates antitumor activity of gemcitabine in an orthotopic model of pancreatic cancer through suppression of proliferation, angiogenesis, and inhibition of nuclear factor-kappaB-regulated gene products. Cancer Res. 2007, 67, 3853–3861. [Google Scholar] [CrossRef] [PubMed]
- Fossey, S.L.; Bear, M.D.; Lin, J.; Li, C.; Schwartz, E.B.; Li, P.K.; Fuchs, J.R.; Fenger, J.; Kisseberth, W.C.; London, C.A. The novel curcumin analog FLLL32 decreases STAT3 DNA binding activity and expression, and induces apoptosis in osteosarcoma cell lines. BMC Cancer 2011, 11, 112. [Google Scholar] [CrossRef] [PubMed]
- Mari, A.; D’Andrea, D.; Abufaraj, M.; Foerster, B.; Kimura, S.; Shariat, S.F. Genetic determinants for chemo- and radiotherapy resistance in bladder cancer. Transl. Androl. Urol. 2017, 6, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Amling, C.L. Diagnosis and management of superficial bladder cancer. Curr. Probl. Cancer 2001, 4, 219–278. [Google Scholar] [CrossRef]
- Herr, H.W.; Wartinger, D.D.; Fair, W.R.; Oettgen, H.F. Bacillus Calmette-Guerin therapy for superficial bladder cancer: A 10-year followup. J. Urol. 1992, 147, 1020–1023. [Google Scholar] [CrossRef]
- Tian, B.; Wang, Z.; Zhao, Y.; Wang, D.; Li, Y.; Ma, L.; Li, X.; Li, J.; Xiao, N.; Tian, J.; et al. Effects of curcumin on bladder cancer cells and development of urothelial tumors in a rat bladder carcinogenesis model. Cancer Lett. 2008, 264, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wang, Y.; Jia, Z.; Gao, Y.; Zhao, C.; Yao, Y. Curcumin inhibits bladder cancer progression via regulation of β-catenin expression. Tumor Biol. 2017, 39, 1010428317702548. [Google Scholar] [CrossRef] [PubMed]
- Botteman, M.F.; Pashos, C.L.; Redaelli, A.; Laskin, B.; Hauser, R. The health economics of bladder cancer: A comprehensive review of the published literature. Pharmacoeconomics 2003, 21, 1315–1330. [Google Scholar] [CrossRef]
- Pan, Z.J.; Deng, N.; Zou, Z.H.; Chen, G.X. The effect of curcumin on bladder tumor in rat model. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 884–889. [Google Scholar]
- Chendil, D.; Ranga, R.S.; Meigooni, D.; Sathishkumar, S.; Ahmed, M.M. Curcumin confers radiosensitizing effect in prostate cancer cell line PC-3. Oncogene 2004, 23, 1599–1607. [Google Scholar] [CrossRef]
- Kamat, A.M.; Tharakan, S.T.; Sung, B.; Aggarwal, B.B. Curcumin potentiates the antitumor effects of Bacillus Calmette-Guerin against bladder cancer through the downregulation of NF-kappaB and upregulation of TRAIL receptors. Cancer Res. 2009, 69, 8958–8966, Erratum in Cancer Res. 2018, 78, 5182. [Google Scholar] [CrossRef]
- Khanbolooki, S.; Nawrocki, S.T.; Arumugam, T.; Andtbacka, R.; Pino, M.S.; Kurzrock, R.; Logsdon, C.D.; Abbruzzese, J.L.; McConkey, D.J. Nuclear factor-kappaB maintains TRAIL resistance in human pancreatic cancer cells. Mol. Cancer Ther. 2006, 9, 2251–2260. [Google Scholar] [CrossRef]
- Kruyt, F.A. TRAIL and cancer therapy. Cancer Lett. 2008, 263, 14–25. [Google Scholar] [CrossRef]
- Duan, W.; Chang, Y.; Li, R.; Xu, Q.; Lei, J.; Yin, C.; Li, T.; Wu, Y.; Ma, Q.; Li, X. Curcumin inhibits hypoxia inducible factor-1α-induced epithelial-mesenchymal transition in HepG2 hepatocellular carcinoma cells. Mol. Med. Rep. 2014, 10, 2505–2510. [Google Scholar] [CrossRef]
- Sindhwani, P.; Hampton, J.A.; Baig, M.M.; Keck, R.; Selman, S.H. Curcumin prevents intravesical tumor implantation of the MBT-2 tumor cell line in C3H mice. J. Urol. 2001, 166, 1498–1501. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, G.; Zhang, R.; Dong, L.; Chen, H.; Bo, J.; Xue, W.; Huang, Y. Curcumin inhibits cell proliferation and motility via suppression of TROP2 in bladder cancer cells. Int. J. Oncol. 2018, 53, 515–526. [Google Scholar] [CrossRef]
- Cho, C.-J.; Yang, C.-W.; Wu, C.-L.; Ho, J.-Y.; Yu, C.-P.; Wu, S.-T.; Yu, D.-S. The modulation study of multiple drug resistance in bladder cancer by curcumin and resveratrol. Oncol. Lett. 2019, 18, 6869–6876. [Google Scholar] [CrossRef]
- Park, B.H.; Lim, J.E.; Jeon, H.G.; Seo, S.I.; Lee, H.M.; Choi, H.Y.; Jeon, S.S.; Jeong, B.C. Curcumin potentiates antitumor activity of cisplatin in bladder cancer cell lines via ROS-mediated activation of ERK1/2. Oncotarget 2016, 7, 63870–63886. [Google Scholar] [CrossRef]
- dos santos, N.A.; Carvalho Rodrigues, M.A.; Martins, N.M.; dos Santos, A.C. Cisplatin-induced nephrotoxicity and targets of nephroprotection: An update. Arch. Toxicol. 2012, 86, 1233–1250. [Google Scholar] [CrossRef]
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Volarevic, V.; Djokovic, B.; Jankovic, M.G.; Harrell, C.R.; Fellabaum, C.; Djonov, V.; Arsenijevic, N. Molecular mechanisms of cisplatin-induced nephrotoxicity: A balance on the knife edge between renoprotection and tumor toxicity. J. Biomed. Sci. 2019, 26, 25. [Google Scholar] [CrossRef] [PubMed]
- Tuyaerts, S.; van Nuffel, A.M.T.; Naert, E.; van Dam, P.A.; Vuylsteke, P.; de Caluwé, A.; Aspeslagh, S.; Dirix, P.; Lippens, L.; de Jaeghere, E.; et al. PRIMMO study protocol: A phase II study combining PD-1 blockade, radiation and immunomodulation to tackle cervical and uterine cancer. BMC Cancer 2019, 19, 506. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Qu, C.; Xie, F.; Chen, L.; Liu, L.; Liang, X.; Wu, X.; Wang, P.; Meng, Z. Curcumin suppresses epithelial-to-mesenchymal transition and metastasis of pancreatic cancer cells by inhibiting cancer-associated fibroblasts. Am. J. Cancer Res. 2017, 7, 125–133. [Google Scholar]
- Shi, J.; Zhang, X.; Shi, T.; Li, H. Antitumor effects of curcumin in human bladder cancer in vitro. Oncol. Lett. 2017, 14, 1157–1161. [Google Scholar] [CrossRef]
- Bava, S.V.; Puliyappadamba, V.T.; Deepti, A.; Nair, A.; Karunagaran, D.; Anto, R.J. Sensitization of taxol-induced apoptosis by curcumin involves down-regulation of nuclear factor-kappaB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J. Biol. Chem. 2005, 280, 6301–6308, Erratum in J. Biol. Chem. 2018, 293, 12283. [Google Scholar] [CrossRef]
- Fan, S.; Xu, Y.; Li, X.; Tie, L.; Pan, Y.; Li, X. Opposite angiogenic outcome of curcumin against ischemia and Lewis lung cancer models: In silico, in vitro and in vivo studies. Biochim. Biophys. Acta 2014, 1842, 1742–1754. [Google Scholar] [CrossRef]
- Soni, V.K.; Mehta, A.; Ratre, Y.K.; Tiwari, A.K.; Amit, A.; Singh, R.P.; Sonkar, S.C.; Chaturvedi, N.; Shukla, D.; Vishvakarma, N.K. Curcumin, a traditional spice component, can hold the promise against COVID-19? Eur. J. Pharmacol. 2020, 886, 173551. [Google Scholar] [CrossRef]
- Rattis, B.A.C.; Ramos, S.G.; Celes, M.R.N. Curcumin as a potential treatment for COVID-19. Front. Pharmacol. 2021, 12, 675287. [Google Scholar] [CrossRef]
- Zahedipour, F.; Hosseini, S.A.; Sathyapalan, T.; Majeed, M.; Jamialahmadi, T.; Al-Rasadi, K.; Banach, M.; Sahebkar, A. Potential effects of curcumin in the treatment of COVID-19 infection. Phytother. Res. 2020, 34, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Perera, P.K.; Meedeniya, A.C.B. Curcumin as a potential target for COVID19: A Concept Letter. Acad. Lett. 2021, 2, 3484. [Google Scholar] [CrossRef]
- Mijatović, S.; Bramanti, A.; Nicoletti, F.; Fagone, P.; Kaluderović, G.N.; Maksimović-Ivanić, D. Naturally occurring compounds in differentiation based therapy of cancer. Biotechnol. Adv. 2018, 36, 1622–1632. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Veena, M.S.; Basak, S.K.; Han, E.; Tajima, T.; Gjertson, D.W.; Starr, J.; Eidelman, O.; Pollard, H.B.; Srivastava, M.; et al. Curcumin treatment suppresses IKKβ kinase activity of salivary cells of patients with head and neck cancer: A pilot study. Clin. Cancer Res. 2011, 17, 5953–5961. [Google Scholar] [CrossRef]
- Hariri, M.; Gholami, A.; Mirhafez, S.R.; Bidkhori, M.; Sahebkar, A. A pilot study of the effect of curcumin on epigenetic changes and DNA damage among patients with non-alcoholic fatty liver disease: A randomized, double-blind, placebo-controlled, clinical trial. Complement. Ther. Med. 2020, 51, 102447. [Google Scholar] [CrossRef]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed]
More