Perennial herbaceous Fallopia is native to East Asia, and was introduced to Europe and North America in the 19th century as an ornamental plant. Fallopia has been spreading quickly and has naturalized in many countries. It is listed in the world’s 100 worst alien species. Fallopia often forms dense monospecies stands through the interruption of the regeneration process of indigenous plant species. Allelopathy of Japanese knotweed (Fallopia japonica), giant knotweed (Fallopia sachalinensis), and Bohemian knotweed (Fallopia x bohemica) has been reported to play an essential role in its invasion. The exudate from their roots and/or rhizomes, and their plant residues inhibited the germination and growth of some other plant species. These knotweeds, which are non-mycorrhizal plants, also suppressed the abundance and species richness of arbuscular mycorrhizal fungi (AMF) in the rhizosphere soil. Such suppression was critical for most territorial plants to form the mutualism with AMF, which enhances the nutrient and water uptake, and the tolerance against pathogens and stress conditions.
1. Introduction
The genus
Fallopia (Polygonaceae) contains highly competitive invasive species such as, Japanese knotweed (
Fallopia japonica (Houtt.) Ronse Decraene; synonym:
Polygonum cuspidatum Sieb. et Zucc.), giant knotweed
(Fallopia sachalinensis (F. Schmidt) Ronse Decraene; synonym:
Polygonum sachalinensis F. Schmidt ex Maxim), Bohemian knotweed (
Fallopia x bohemica (Chrteket Chrtková) J.P. Bailey; synonym
Polygonum x bohemicum (Chrtek et Chrtková) P.F. Zika et A.L. Jacobson). Japanese knotweed and giant knotweed hybridize naturally by sexual reproduction and create the hybrid Bohemian knotweed
[1].
These knotweeds are fast-growing perennial herbaceous plants and form monospecific stands with dense canopy, and expand the stands through their extensive rhizome systems
[2]. Japanese knotweed grows 2–3 m in height, and has multiple branches with hollow stems. Its shoots were recorded to grow 4–15 cm per day
[3]. Its ovate leaves are 3–10 cm in length with a truncate base. Giant knotweed is 3–6 m in height, and has few branches with hollow stems. Its ovate leaves are 20–40 cm in length with a deeply cordate base. Characteristic of Bohemian knotweed has variations between both species
[2].
Japanese knotweed grows under a wide range of soil pH, and nutrient poor soil conditions such as volcanic slopes as a pioneer species, and in disturbed areas such as roadsides and pastures
[4][5][6][4,5,6]. Giant knotweed occurs in riparian corridors, coastal cliffs, road banks, and bare soils in human settlements
[3]. Those knotweeds show a strong preference for man-made disturbed habitats, and along roads, and watercourses are their most frequent habitats. However, Bohemian knotweed has the highest population in man-made habitats
[7][8][7,8].
Knotweeds spread sexually and asexually. The seeds of knotweeds are buoyant, and carried with water flow in streams and rivers for a long distance. The seeds are also easy to disperse with the wind
[9][10][9,10]. The seed distribution may contribute to establishing knotweed populations in new habitats. Knotweeds also propagate with their rhizome branches. The rhizomes grow quickly, and the apex of the rhizome branches develops into an aerial shoot and forms a new shoot clump
[4]. Rhizome fragments made by such as floods and human activities, as small as 1 cm in length and 0.7 g fresh in weight can regenerate new plants
[11]. The fragmentation and subsequent regeneration increase their propagation potential.
The native range of Japanese knotweed is China, Japan, Korea, and Taiwan, and giant knotweed is native to Sakhalin Island, North Japan, and Korea
[2]. Both species were introduced into Europe in the 19th century as ornamental plants and collections for botanical gardens, and were sold to gardens and parks in various countries
[2][12][2,12]. Japanese knotweed was also cultivated as a Chinese medicinal plant
[13]. Giant knotweed was recommended as riverbank stabilizer and livestock feed
[14]. Although hybrid species, Bohemian knotweed was first recorded in Europe around 1980s, it was considered to occur in the late 19th century and spread undetectably
[15]. The Japanese knotweed and giant knotweed were not sympatric in the native range. However, Bohemian knotweed was also observed in Japan in 1997
[16]. Japanese knotweed is the most widespread species in Europe, and followed by Bohemian knotweed and giant knotweed
[8][17][8,17].
Japanese knotweed and giant knotweed were introduced into North America in the 19th century. The presence of Bohemian knotweed was first recognized in 2001 from Seattle, USA
[18]. The presence of the knotweeds has been confirmed in 42 states including Alaska and 8 provinces in Canada
[19][20][19,20]. These knotweeds were also confirmed in Australia, New Zealand, South Africa, and other countries
[15][16][15,16]. Knotweed species have already naturalized in many countries as invasive noxious weeds, and are listed in the world’s 100 worst invasive alien species
[21].
The characteristics of life-history of plants, such as high growth and high reproduction rate, and phenotypic plasticity are involved in the naturalization of invasive plants into the introduced range
[22][23][24][22,23,24]. As described above, knotweeds grow fast and reproduce sexually and asexually. Those species can grow under poor soil conditions and in disturbed places. The single stem of Japanese knotweed bore about 200,000 seeds, and the germination rate was 50–80%
[25][26][25,26]. Soil collected from the invaded site of Bohemian knotweed contained about 800 seeds/m
2 [27]. Genetic diversity of each knotweed species is not clear. However, knotweeds showed large epigenetic differentiation and phenotypic plasticity in response to several environmental conditions
[28][29][28,29]. Epigenetics diversity provides clonal plants with the potential for acclimation in various environmental conditions
[30]. Hybridization of Japanese knotweed and giant knotweed probably gives genetic diversity to a hybrid plant, Bohemian knotweed
[1][31][32][1,31,32].
High defense capacity against pathogens and herbivores contributes for the invasive plants to naturalize into introduced range
[33][34][35][33,34,35]. Japanese knotweed has a lower abundance and diversity of invertebrate herbivores compared to native plant species of introduced range, and received less leaf damage than their native plant species
[36]. Knotweed contains flavones, quinones, and stilbenes
[13][37][13,37], and some of these compounds may act as defensive agents against herbivores and pathogens. In addition, aqueous extracts of rhizomes of Japanese knotweed altered soil fauna and reduced nematode population
[38].
2. Allelopathy of Knotweeds
Allelopathy is the interaction between one plant and other neighboring plants through the specific secondary metabolites which are defined as allelochemicals
[39][42]. The allelochemicals are released into their rhizosphere soil and neighboring environments either by root exudation, decomposition of plant residues, rainfall leachates, or volatilization from living plant parts
[40][41][42][43,44,45].
Plant-to-plant interaction is a complex combination of competition for resources such as water, nutrients, and light, along with allelopathic interaction through allelochemicals
[43][44][46,47]. Thus, it is essential to eliminate such competitive effects from experimental systems to clarify allelopathy
[45][46][47][48,49,50]. Bohemian knotweed reduced the growth and survival rate of native plant species,
Eupatorium perfoliatum L. and
Acer saccharinum L. in field conditions. Supplemental nutrient and light to the growth conditions for those native plants recovered the inhibitory effects of the knotweed, but the recovery was limited. Therefore, allelopathy of Bohemian knotweed may contribute to a certain extent of the reduction in the growth and survival rate of the native plant species
[48][51].
A rhizome (contain single node) of Bohemian knotweed and each of six native plant species (four forbs;
Geraniun robertianum L.,
Lamium maculatum L.,
Silene dioica (L.) Clairv. and
Symphytum officinalis L.; and two grasses;
Lolium perenne L. and
Poa trivialis L.) were grown together in 7 L pots filled with soil for three weeks. The rhizome of Bohemian knotweed significantly suppressed the growth of four forbs but not grasses. Activated carbons mixed with soil in the pots reduced the inhibitory effects of the knotweed on the forbs
[37]. Activated carbon absorbs organic compounds in the soil
[49][52]. Therefore, the result indicates that allelochemicals released from the rhizome of Bohemian knotweed may be involved in the inhibition. The evidence of allelopathy of knotweeds has been accumulated over three decades. In this section, allelopathic potential of the exudation, extracts, and plant residues of knotweeds was summarized (
Table 1).
Table 1. Allelopathic activities of roots, rhizomes, and above-ground parts of knotweeds.
| Source |
|
Knotweed |
Target Plant Species |
Inhibition |
Reference |
| Root, rhizome |
|
|
|
|
|
| |
Exudation |
Japanese knotweed |
Salix viminalis, Salix atrocinerea, Populus nigra |
Growth |
[50][53] |
| |
|
Giant knotweed |
Lactuca sativa |
Growth |
[51][54] |
| |
Rhizome extract |
Japanese knotweed
Giant knotweed
Bohemian knotweed |
Leucosinapis alba |
Growth |
[52][55] |
| |
|
Japanese knotweed
Bohemian knotweed |
Raphanus sativus |
Growth
Germination |
[53][54][56,57] |
| |
|
Japanese knotweed |
Atrichum angustatum
Thuidium delicatulum |
Biomass |
[55][58] |
| Above-ground part |
|
|
|
|
|
| |
Leaf residue |
Japanese knotweed
Giant knotweed
Bohemian knotweed |
Leucosinapis alba
Brassica napa |
Germination |
[56][59] |
| |
Leaf extract |
Japanese knotweed
Giant knotweed
Bohemian knotweed |
Urtica dioica
Calamagrostis epigejos
Lepidium sativum |
Germination |
[57][60] |
| |
Soaking water |
Japanese knotweed |
Triticum aestivum
Sinapis arvensis |
Germination |
[58][61] |
| |
Extract (whole part) |
Japanese knotweed
Giant knotweed |
Brassica napa, Avena sativa
Helianthus annuus |
Growth |
[59][62] |
2.1. Exudation
Japanese knotweed was grown in “donor pots”, and test plants (
Salix viminalis L.,
Salix atrocinerea Brot. and
Populus nigra L.) were grown in “target pots”. Test plants were irrigated with drain solution from the donor pots and with additional nutrient-enriched solution, and incubated for four months. The system could discriminate resource competition between Japanese knotweed and test plants. The growth of those test plants was suppressed by the drain solution from Japanese knotweed. The drain solution contained polyphenol compounds, but those compounds were not identified
[50][53]. Giant knotweed (donor plant) and
Lactuca sativa L. seedlings (receiver plant) were incubated with root exudate recirculating system
[60][63] for 10–14 days. Exudate from roots and rhizomes of giant knotweed significantly inhibited the growth of
L. sativa seedlings
[51][54]. According to those observations, exudation from rhizome and/or roots of those knotweeds may contain allelochemicals, which cause the suppression of the growth of the test plant species. Although the organs that synthesize those allelochemicals are unknown, certain allelochemicals may be released from rhizomes and/or roots of knotweeds. Therefore, the evaluation of allelopathic potential of extracts of rhizome and/or roots is necessary.
Aqueous rhizome extracts of Japanese knotweed, giant knotweed, and Bohemian knotweed inhibited the root and hypocotyl growth of
Leucosinapis alba (L.) Spach. However, the inhibitory effects of three extracts were not significantly different
[52][55]. Aqueous rhizome extracts of Japanese knotweed and Bohemian knotweed also delayed the germination of
Raphanus sativus L., and suppressed their growth. The roots of
R. sativus showed symptoms of oxidative stress such as abnormal shapes of nuclei, plasma membrane, mitochondria, and endoplasmic reticulum
[53][54][56,57]. Aqueous rhizome extracts of Japanese knotweed (
Polygonum cuspidatum s.l.) inhibited the biomass of the mosses,
Atrichum angustatum (Brid.) Bruch and Schimp. and
Thuidium delicatulum (Hedw.) Schimp.
[55][58]. Those findings indicate that the rhizomes of knotweeds may contain allelochemicals, and those allelochemicals are extractable.
2.2. Plant Residues
Japanese, giant, and bohemian knotweeds are perennial herbaceous plants, but above-ground parts of the knotweeds die back at the first frost in the winter season
[21]. Those above-ground parts decay and accumulate as a litter layer on the soil. During the decomposition process of the litter, some of the secondary metabolites may be liberated into rhizosphere soil and act as allelochemicals
[40][41][42][43,44,45]. Therefore, evaluation of the allelopathic potential of above-ground parts of knotweeds is also necessary.
Leaves of Japanese knotweed, giant knotweed, and Bohemian knotweed were shattered into small pieces and mixed with soil, and the seeds of
Leucosinapis alba (L.) Spach,
Brassica napa L.,
Chenopodium album L., and
Echinochloa crus-galli (L.) P.Beauv were sown into the mixture. The germination of
L. alba and
B. napa was significantly suppressed by the residues of those knotweed leaf residues. The inhibitory activity of the residue of Japanese knotweed was the highest. However, all residues did not significantly inhibit the germination of
C. album and
E. crus-galli [56][59]. Aqueous leaf extracts of Japanese knotweed, giant knotweed, and Bohemian knotweed inhibited the germination of
Urtica dioica L.,
Calamagrostis epigejos (L.) Roth, and
Lepidium sativum L. However, the inhibitory effect of Japanese knotweed was the least
[57][60]. Senescent above-ground parts of Japanese knotweed were soaked in water. Seeds of
Triticum aestivum L. and
Sinapis arvensis L. were sown into the soil, and irrigated with the soaking water of the Japanese knotweed every two days for two weeks. The irrigation of the soaking water resulted in significantly reduction in their germination
[58][61]. Aqueous extracts of whole plants of Japanese knotweed and giant knotweed also inhibited the growth of
Brassica napa L.,
Avena sativa L., and
Helianthus annuus L.
[59][62]. Those findings indicate that above-ground parts of those knotweeds may contain allelochemicals. Those compounds may be released into the soil during the decomposition process of the plant residues. However, the efficiency of allelopathic potential among Japanese knotweed, giant knotweed, and Bohemian knotweed is not able to compared because of the limited information.
3. Invasion and Allelopathy of Knotweeds
Perennial plants are able to release allelochemicals into the rhizosphere soil over several years through the decomposition process of plant parts including fallen leaves, and the exudation from their rhizomes and roots, and those allelochemicals may be able to accumulate in the soil
[61][62][63][64][65][66][67][80,81,82,83,84,85,86]. The invasion of perennial herbaceous species, knotweed significantly reduced the plant diversity and abundance of native herbs, shrubs, and juvenile trees in the introduced range
[68][69][87,88]. The invasion of knotweed also suppressed long-term native tree regeneration and shifted from tree-dominated riparian forests to knotweed-dominated herbaceous shrublands
[70][89].
According to the novel weapon hypothesis, some invasive plant species may have particularly strong allelopathic activity and success in their invasion into the introduced range. Allelopathy of the invasive plants is mediated by allelochemicals that are new to the plant species in the introduced range. Those indigenous plants species in the introduced range are susceptible to the allelochemicals. The plants that co-evolved with invasive plants had the opportunity to obtain the defense systems against those allelochemicals. However, the plant species in the introduced range have not obtained the defense systems that obviate those allelochemicals
[71][72][40,73]. Root powder of Japanese knotweed suppressed the germination of
Ulmus minor Mill. (native in Europe), but did not suppress the germination of
Ulmus parvifolia Jacq. (native in East Asia, same as knotweed)
[73][90]. The concentrations of piceid (
10) and resveratrolside (
11) in Japanese knotweed from Switzerland (introduced range) were higher than in those from China (native range)
[74][67]. Those observations may support the novel weapon hypothesis for the invasive plant species.
In addition, the invasion of Japanese knotweed reduced the abundance and species richness of arbuscular mycorrhizal fungi (AMF)
[75][91]. Root powder of Japanese knotweed also inhibited AMF vesicle formation in
Ulmus spp. [73][90]. Mycorrhizal colonization is crucial for most territorial plants. AMF fungi are widespread and important mycorrhiza for plant symbionts. AMF fungi increase the ability of plants to absorb nutrient and water, and enhance the protection potential against pathogen attacks and several stress conditions
[76][92]. Knotweeds are non-mycorrhizal plants and do not form arbuscular mycorrhiza
[77][78][93,94]. Knotweeds may be able to degrade the fungal mutualism of nearby plants without negatively impacting their own nutrient and water acquisition. The reduction in AMF populations in the soil of the knotweed introduced range weakens the ability of residential plants for the competition and regeneration, and leads knotweed to dominant communities in the introduced range. Allelochemicals have the potential to inhibit plant mutualism with AMF fungi
[79][95]. Therefore, allelochemicals released from the knotweeds may cause the reduction in AMF communities in the soil, and contribute to the knotweed invasion. The suppression by allelochemicals on AMF communities were also found in non-mycorrhizal species,
Alliaria petiolate (M. Bieb.) Cavara and Grande (Brassicaceae)
[80][81][96,97].
Soaking water of giant knotweed (aboveground parts) suppressed the population of soil fungal pathogens
[58][61]. Bohemian knotweed and indigenous plant species were grown in the soil collected from knotweed-uninvaded areas. The community of soil bacteria shifted the balance in favor of the knotweed, and promoted the growth and regeneration of the knotweed more than those of indigenous plant species. This promotion effect was reduced by adding activated carbons to the soil
[31]. The observation suggests that certain compounds released from the knotweed may change the balance of soil bacteria in favor of the knotweed. However, further investigation is necessary to identify those allelochemicals.
Many of the phytotoxic substances from the invasive plants have been reported to have multiple functions such as allelopathic, anti-pathogen, anti-herbivore activity, and provide the invasive plants with the advantage in the increasing their population in the introduced range
[24][82][83][24,41,98]. Large numbers of secondary metabolites have been isolated from knotweeds, such as stilbenes, quinones, flavonoids, coumarins, and lignans
[13]. Among them, for instance, emodin was reported to work as a defense compound for pathogens, herbivores, and abiotic stress factors
[84][99]. Resveratrol and piceid may also be involved in the interaction of plant–pathogen and plant–herbivore
[85][86][100,101]. Japanese knotweed plant itself was also reported to possess antipathogen and antifungal effects
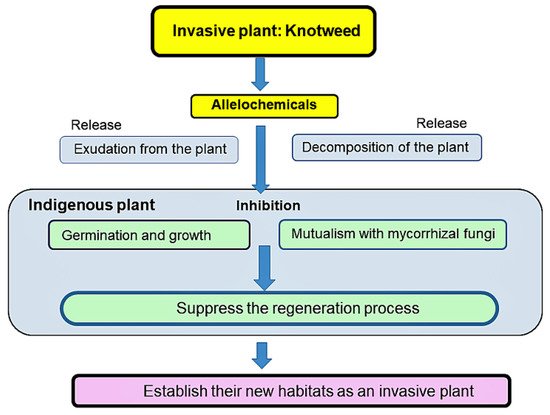
[13][87][13,76]. Some of those compounds may enhance competitive ability of knotweeds and make the plant invasive. As describe previously, knotweeds may interrupt the regeneration process of indigenous plant species by decreasing their germination and growth directory, and indirectly via the suppression of mycorrhizal colonization to indigenous plant species (
Figure 13). In addition, the elevated temperature from 2000 to 2008 results in an increase to 35–53% of the habitat being suitable for knotweed in southern Ontario, Canada
[88][102], indicating that global warming may increase the threat of the invasion of the species into the additional introduced areas.
Figure 13. A possible scheme of knotweed to establish the new habitats.