The present review aims to characterize the general characteristics, physicochemical properties,
and health-related studies of onion, garlic, and cruciferous vegetable consumption, as well as the
influence of food processing on their content of active substances. The review combines distinct
chapters where information relevant for nutraceutical and functional food engineers, medical doctors,
as well as consumers can be obtained. We believe that the review will stimulate further studies related
to the implementation of the most accurate sources of sulfur compounds from chosen plants which
could support the therapy of cancer patients. Moreover, the review aims to present cost-e ective
anti-tumor means that would include the implementation in the diet of sulfur-based compounds
in foods.
- alliaceous vegetables
- cruciferous vegetables
- sulfur-based compounds
- food processing
- nutraceuticals
- functional food
The broad spectrum of the mechanism of action of immune-boosting natural compounds as well as the complex nature of the food matrices make researching the health benefits of various food products a complicated task. Moreover, many routes are involved in the action of most natural compounds that lead to the inhibition of chronic inflammation, which results in a decrease in the ability to remove a pathogen asymptomatically and is connected to various pathological events, such as cancer. A number of cancers have been associated with inflammatory processes. The current review strives to answer the question of whether plant-derived sulfur compounds could be beneficial in cancer prevention and therapy.
- Introduction
1. Introduction
Attempts to modulate the immune system started in the mid-1970s [1] [1] with the introduction of bacterial toxins. The main disadvantages of those therapies were connected to regulatory problems, the impurity of the exogenic modulatory cocktail, or the occurrence of multiple side effects. However, the discoveries in the field of natural components of the food that could display immunomodulatory properties led to the recognition of novel natural compounds that could preserve the homeostasis of the human immune system. These natural therapies include natural plant-derived food compounds that have been proved to be beneficial over synthetic or uncontrolled (such as neutralized pathogens) compounds [2]. The natural immune boosters present a really promising approach for immunomodulation and could be beneficial over synthetic drugs due to their lower cost, decreased or even eliminated toxicity, and few adverse effects. The study of natural immune boosters is relevant also because the imbalance in the immune system is involved in the pathogenesis and progression of several serious diseases, such as infectious and autoimmune diseases, allergies, obesity, metabolic syndrome, cardiovascular disorders, gastric ulcers, and cancer [3][4][3,4]. Nutraceuticals can exert indirect antitumor effects via immune enhancement [5]. In terms of cancer, an up to 40% decrease in cancer cases could be gained just via the administration of proper food and nutrition [6]. A connection between inflammation and cancer has been recognized since the 17th century, and over 150 years ago we were provided with a scientific rationale by Robert Virchow [7]. A number of cancers have been associated with inflammatory processes. This now has some implications for their prevention and treatment [8]. Indeed, there is solid evidence that the increased risk of malignancy is associated with the chronic inflammation caused by chemical and physical agents and different inflammatory reactions involving leucocytes and tumor cells. The focus has been on cytokines and chemokines; their expression; their mechanisms of action; and, consequently, their preventive and therapeutic targeting of inflammation [9]. The basic interactions between different nutrients, including those of plant origin; pro-inflammatory cytokines; and inflammation have been succinctly presented by Grimble [10,11][10][11]. In the context of the current review, the obvious question is whether plant-derived sulfur compounds can be beneficial for cancer treatment as a supportive therapy. Numerous plant compounds (saponins, alkaloids, phenolics, triterpenes, coumarins, polysaccharides, anthraquinones, etc.) exert antitumor properties, not only by a direct toxic effect on malignant cells but also by influencing macrophage phenotypic differentiation and, possibly, cytokine secretion [12]. Among them, the sulfur compounds found in the alliaceous and cruciferous families were studied for their potential in cancer treatment.
Cruciferous vegetables including broccoli (Brassica oleracea var. italica), cabbage (Brassica oleracea var. capitata), cauliflower (Brassica oleracea var. botrytis), and other plants of the family brassicaceae are rich sources of bioactive compounds, including vitamin C and E, dietary fiber, and the glycosides of the flavonoids quercetin and kaempferol. However, the components that make cruciferous vegetables so beneficial to human health and set them apart from other plants are the sulfur-containing glycosides (glucosinolates, GSLs). Dietary GSLs are considered as potential anticancer agents [13]. The bioactive substances found in cruciferous vegetables can inhibit inflammation by activating detoxification enzymes, clearing free radicals, and inducing immune functions.
Onion (Allium cepa) and garlic (Allium sativa) are some of the oldest cultivated plants in the world and are rich in flavonols and natural organosulfur compounds (OSCs). Their consumption was studied, as it was considered beneficial for heart diseases, tumors (such as esophageal, gastric, colorectal, breast, lung, skin and prostate cancers), and immunological processes, among others, especially due to the biological action of the sulfur-containing metabolic byproducts [14–16][14][15][16]. They act by the induction of phase II detoxification enzymes [17], take part in the scavenging of free radicals, and change the morphology of the cells that abort cell-to-cell contact [18]—effects that all together are known to contribute to cancer inhibition. On the other hand, the side effects of the excessive consumption of these plants have been described. Since the studies have been conducted mainly in animal models, the need for the full characterization of the proper dosages of OSC consumption remains an important task for further study [19]. Moreover, the need to precisely demonstrate the bioavailability of sulfur-containing compounds from various types of functional food remains a challenge.
- Plant-Derived Sulfur Compounds—GSLs and OSCs
2. Plant-Derived Sulfur Compounds—GSLs and OSCs
2.1. Glucosinolates (GSLs)
Sulfur is an essential compound for the biosynthesis of various secondary metabolites, such as phytoalexins, GSLs, and alliins. These metabolic compounds play a vital role in the physiology and protection of plants against several environmental stresses [20]. More than 200 different types of GSLs have been studied, and 30 of them are identified in Brassica crops [20,21][20][21]. GSLs are major secondary metabolites synthesized by plants. GSLs are present in various species of the Brassica family. These are responsible for the nutritional characteristics of Brassica products, such as vegetables, oil, and meal [22]. Moreover, they are also responsible for the flavor and taste of Brassica products. The breakdown products of GSLs play a vital role in plant defense mechanisms against pathogens and insects. In addition, these breakdown products have gained much attention because of their anti-carcinogenic properties [23].
2.1.1. Chemical Structure and Biodegradation
GSLs are organic anions molecules comprised of β-D-thioglucoside linked to the (Z)-N-hydroximinosulfate group and a variable side chain derived from a variable amino acid R-group [24][24]. GSLs molecules are synthesized by the action of different chemical compounds, including two cytochromes P450, glucosyl transferase, C-S lyase, and sulfotransferase. GSLs can be divided into three classes—aliphatic, indole, and aromatic—which are derived from amino acid precursors such as methionine, tryptophan, and phenylalanine, respectively (Figure 1) [25,26][25][26]. GSLs with an aliphatic side chain derived from methionine are most prominent in the leaves of Brassica vegetables. Aliphatic GSLs are classified by the size of carbon side chains, such as 3-carbon, 4-carbon, and 5-carbon GSLs. They can also be classified by the structure of the side-chain, such as hydroxyalkeny, l methylsulfinylalkyl, methylsulfanylalkyl or methylsulfonylalkyl glucosinolates [27].
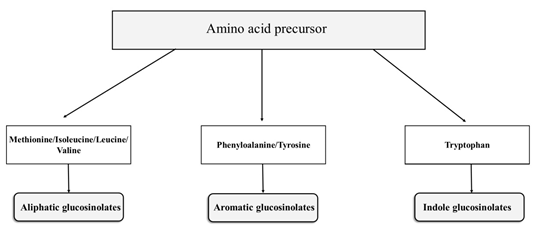
Figure 1. Glucosinolate classification based on the structure of their amino acid precursors. Among the common amino acid precursors of glucosinolates are aromatic amino acids, phenylalanine, or tyrosine; aliphatic amino acids include alanine, leucine, isoleucine, methionine, and valine, whereas indole glucosinolates are those derived from tryptophan.
GSLs are relatively stable compounds, but plant cells containing glucosinolates can be damaged due to the release of β-thioglucosidase or myrosinase enzymes during the cutting, mixing, chopping, and chewing of food (Figure 2). This enzyme is usually stored separately from GSLs, in different cells depending on the plant species [28,29][28][29] . The hydrolysis of GSLs by the enzyme myrosinase yields a β-D glucose molecule and an unstable aglycone (thiohydroximate-O-sulfonate). Upon the cellular injury of plant tissues, GSLs are enzymatically degraded by myrosinase (a thioglucosidase) to produce a variety of different breakdown products, such as glucose, sulfate, isothiocyanates, epithionitriles, nitriles, indolic alcohols, oxazolidinethions, amines, and thiocyanate. It is believed that both GSLs and their breakdown products play an important role in the defense mechanism of plants against the attacks of fungi, insects, and microorganisms [23,30][30]. Moreover, these are also presumed to have different health-promoting roles, which makes them beneficial compounds for human consumption.
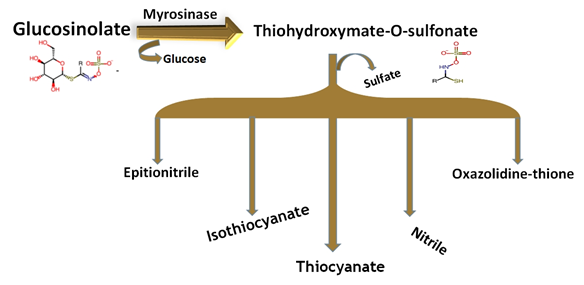
Figure 2. Breakdown of glucosinolates and the possible alternative breakdown products.
In order to preserve the beneficial properties of cruciferous plants, it is important to store and prepare them properly. The content of glucosinolates varies between cruciferous plant types, but the bioavailability of these compounds can be significantly increased by appropriate storage and preparation. The requirements for storage conditions vary in the available literature. Some reports indicate long-term storage at room temperature and in the refrigerator. Others suggest a significant decrease in GSLs after seven days of storage. An even greater decrease was noted after 6 h of storage for cruciferous vegetables that were previously chopped [31]. In the analyzed brassicaceae vegetables, the highest GSL content was found in purple cauliflower, while green cauliflower had the lowest content of those compounds [32]. However, the inappropriate preparation of cruciferous vegetables can significantly reduce the content of the mentioned compounds. Various processes used to prepare cruciferous vegetables for consumption, such as cutting, crushing, or cooking, disintegrate the cells and are a strong stress factor that stimulates the GSL hydrolysis reaction. As a result of boiling different brassicaceae vegetables for several minutes, a significant (36%) reduction in GSLs was observed. Therefore, in order to keep the content of the described compounds as high as possible, it is recommended to choose methods that require a small amount of water. GSL and its derivatives in cruciferous vegetables have a high potential for use as functional food. However, high-temperature cooking practices such as boiling, frying, blanching, and high-power microwave processing can degrade the bioavailability of GSLs, vitamins, phenolics, and other health-promoting compounds in brassicaceae vegetables [33,34][33][34]. Many studies indicate that the most efficient method for processing these vegetables is steam cooking, as it preserves many nutrients and other compounds beneficial to human health, including GSLs [35].
Several researchers have studied the thermostability and degradation of the GSLs contained in broccoli, red and white cabbage, and brussel sprouts. The results obtained indicate that the important determining factor is media type (food matrix or cooking water) [36]. Florkiewicz et al. [37] suggested sous-vide as the most appropriate and beneficial method of preparing cruciferous vegetables. Inappropriate thermal processing causes the degradation not only of GSLs, but also of indoles and other isothiocyanates. In turn, the research of Kapusta-Duch et al. [38] has shown a very high level (48.5; 75.8%) of indole decomposition as a result of boiling for 15 min at 100 °C. Under the same conditions, isothiocyanates (11.0; 42.4%) as well as ascorbic acid (33.24; 16%) also degrade significantly [39]. Whereas, Westphal et al. [40] evaluated the impact of high-pressure processing (HPP) on GSLs and their breakdown products. For instance, the activity of myrosinase, an enzyme which hydrolyzes GSLs and improves beneficial health effects, was not impaired in HPP-treated vegetables. However, the level of conversion was highly dependent on the parameters used. Additionally, the UV light seems to be not without significance as far as the activity of enzymes crucial for GSL metabolism is concerned [41]. The UV light stimulates, among other things, peroxidase activity, and affects the metabolism of flavonoids and promotes the antioxidant capacity of different cruciferous plants. Formica-Oliveira et al. [42] have confirmed that the use of UV radiation (2.2, 8.8, and 16.4 kJ/m2/day) has a positive effect on the content of GSLs in broccoli. One of the newer and more promising methods that can be used in the treatment of cruciferous vegetables while maintaining a high GSL content is pulsed electric field (PEF). The advantage of this technique is the possibility to penetrate into the plant tissue without using a very high temperature and damaging the tissue mechanically [43]. With the proper selection of parameters, this method may prove extremely useful, although only a few reports confirm its beneficial impact on the content of GSLs and other compounds, as well as their derivatives.
An important factor determining the bioavailability of GSLs is also human microflora. The most crucial role from the point of view of GSL metabolism is played by such enzymes as myrosinase and β-thioglucosidase. β-thioglucosidase is present in the human gut microbiome and influences human physiology and health, for example by regulating the metabolism of GSLs and their derivatives. In turn, the human homolog of myrosinase has not been described so far. The direct administration of isothiocyanates results in their higher bioavailability than that of GSLs, which indicates fundamental differences in such properties as lipophilicity. Therefore, a detailed knowledge of the factors determining the efficiency of the GSL to isothiocyanates conversion seems to be crucial [28]. A common method for assessing bioavailability is to measure the concentration of a particular compound (for example, sulforaphane (SFN)) or its metabolites in urine. In one of the clinical trials, the volunteers were given SFN- or glucoraphanin (GR)-rich beverages, and then the level of metabolites in urine was studied. The level of metabolites tested was significantly higher in SFN recipients. Other studies carried out on a group of healthy volunteers in China showed a high rate of SFN metabolite removal. Studies were also conducted to compare the degree of absorption of particular compounds from fresh and processed food. Four healthy volunteers who were given 3-day-old broccoli sprout extract hydrolyzed by myrosinase, which showed high levels of isothiocyanates. Other researchers have confirmed significantly higher levels of SFN in urine and blood plasma after the ingestion of fresh broccoli sprouts than commercially available myrosinase-free dietary supplements rich in glucoraphanin. However, another clinical trial carried out on a small group of individuals showed a similar level of bioavailability both when taking a commercially available supplement rich in GR but free of myrosinase and frozen broccoli sprout extract, suggesting a large individual variation in the absorption of the compounds described. It was also found that the factor determining the bioavailability of GSLs is the presence of myrosinase. This was confirmed by studies comparing the level of SFN metabolites after the consumption of powder prepared from fresh broccoli rich in GSLs without myrosinase and with it, respectively. It can undoubtedly be deduced that the way cruciferous vegetables are stored and prepared is of key importance for the preservation of compounds beneficial to the proper functioning of the human organism, such as GSLs and their metabolites. On the basis of the described reports, it can be summarized that the consumption of raw cruciferous vegetables, which allows one to maintain a high activity of endogenous myrosinase, is the most appropriate from the point of view of the bioavailability of the analyzed compounds. Since the pharmacokinetics of GSLs are complex and depend on many factors, such as the type of food matrix, the temperature, and the type of vegetable processing, as well as type of starting product (raw or processed vegetable) and the individual differentiation of the influence of the intestinal microflora, further research is necessary to be able to fully explain their potential in many fields of science and especially medicine [44].
In addition to the scientific challenges, the creation of supplements from properly prepared cruciferous plants to increase the degree of GSL assimilation is a great opportunity, especially since heat treatment is one of the factors negatively affecting the degree of absorption. In the United States, there is a growing increase in the sale of dietary supplements from cruciferous plants, which are prepared in the form of extracts and powders and then sold in capsule form. This again confirms the need for a deeper analysis of many factors influencing the transformation and metabolism of the compounds they contain, not only in healthy people but also in the interactions with drugs taken or the possible therapeutic potential. The Food and Drug Administration (FDA) has also issued a number of warnings about the information on the labels of these types of dietary supplements that provide anti-cancer properties and other beneficial effects but which have not yet been confirmed by clinical trials [44]. This is particularly important given that not all the mechanisms that influence the sulfur compounds contained in cruciferous vegetables are fully understood.
2.1.2. Metabolism Pathway of Glucosinolates in the Body
Currently, little is known about how GSLs and their derivatives are metabolized in the human organism, as most reports relate to cell culture and animal studies. Data in humans are extremely sparse and mostly relate to the level of metabolites in the urine. With many compounds, it is the case that, despite the high level of absorption, the bioavailability is limited and reduced due to the high metabolic rate. With regard to the literature data on GSL metabolites, they are most often the result of the level of nitrogen compounds, isothiocyanates, and other derivatives. Additionally, the measurement of the level of metabolites in urine confirms that the hydrolysis reaction is an important criterion for the bioavailability of GSLs. The level of metabolites resulting from prior hydrolysis was six times higher than that of GSLs [45]. The highest concentration and level of glutathione and glutathione s-transferase activity have been reported in the liver. An increased concentration of isothiocyanates results in a decrease in the glutathione activity. Even with isothiocyanates which are similar in structure, their half-lives can vary by up to several hours. The kidneys are the main organ in which the conversion of glutathione conjugates into mercapturic acids takes place. This reaction is catalyzed by γ-glutamyltransferase. There are also many indications that there is a relationship between the structure of the isothiocyanate side chain and the intensity of a particular metabolic pathway, which translates directly into metabolites in urine [46]. The bacterial microflora of the human gastrointestinal tract is also directly related to the metabolism of GSLs. In human feces were identified, for example, a strain of the Bacteroides thetaiotaomicron, which is capable of breaking down GSLs. The high level of bacterial myrosinase activity allows a significant increase in the efficiency of the GSL metabolism, which translates into a high level of their metabolites in the feces. The main factors determining the absorption of GSLs from the intestinal lumen are not endogenous enzymes, but myrosinase activity in the plant tissues, which also affects the absorption of nitriles into the circulation. In the absence of active myrosinase, some GSLs can be absorbed directly from the human digestive tract, but the significance of this pathway is not fully understood [47].
- Health Benefits of Glucosinolates
3. Health Benefits of Glucosinolates
The phytochemicals in cruciferous plants can exert different effects that affect the immune system. Inflammation leads to a number of changes in the organism, such as increased cellular proliferation; the inhibition of apoptosis; and, as a result, an increased risk of various types of cancer. Biologically active substances that occur in cruciferous plants inhibit the development of inflammation by, for example, activating detoxification enzymes, removing reactive oxygen species, and stimulating natural defense mechanisms [31]. Specific compounds of certain vegetables of the family brassicaceae block NF-κB activation; this is done through various mechanisms, such as the inhibition of TNF-α, IκBα, and p65 protein, which also regulates the pro-inflammatory signaling pathway. This effect was primarily mediated by SFN.
In addition to those effects described above, SFN shows a wide range of impacts. It blocks apoptosis, inhibits the release of reactive oxygen species, and affects the expression of vascular cell adhesion molecule (VCAM)-1 or key molecules in the IL-1 signaling cascade. Other important properties of this isothiocyanate include the suppression of such inflammatory reaction markers as interferon regulatory factor 3 (IRF3), as well as macrophage migration inhibitory factor (MIF). These beneficial effects of SFN are the result of the activation of various signaling pathways and mechanisms, which involve, among other Toll-interleukin-1 receptor domains, Toll-like receptors (TLRs) and interferon-beta. Among the numerous activities of SFN, its chemopreventive potential cannot be overlooked. Indeed, SFN suppresses cyclooxygenase-2 (COX-2), which has immunoreactivity properties. These effects were again mainly NF-κB- and ERK (extracellular-signal-regulated kinase)-mediated. Another important compound of cruciferous vegetables with a significant impact on the immune system is berteroin (5 methylthiopentyl isothiocyanate). It is found in the largest quantities in vegetables, such as cabbage and rucola. It has been shown that its frequent consumption reduces edema by modulating COX-2, NF-κB, and ERK activity. There are also reports indicating the beneficial effect of cruciferous vegetables on the immune system parameters in healthy people. These concerns, in particular, the concentrations of circulating inflammatory biomarkers, such as IL-6, CRP, and sTNFRI. However, further more complex studies are needed to confirm that a cruciferous vegetable-rich diet reduces high levels of inflammatory markers in populations where this pathological condition persists chronically [48].
Glucosinolates and their metabolites, such as isothiocyanates like sulforaphane (SFN), have several health-promoting activities (Table 1). Sulfur-containing glycosides (glucoisnolates) are not biologically active but are hydrolyzed by myrosinases to indoles, thiocyanates, and isothiocyanates, which exhibit biological potential. Some isothiocyanates (for example, SFN is one of the best known isothiocyanates), are useful in stress response and anti-inflammation due to their inhibition of transcription regulator (NF-κB). Others play an important role in activating the synthesis of regulatory protein Nrf2, which promotes antioxidants and phase II microsomal enzymes [49].
For example, SFN is one of the best known isothiocyanates, mainly due to its anticancer properties, but also because of its ability to reduce the secretion of inflammation markers by immune system cells, as well as nuclear factor kappa B (NF-κB), which plays an important role in the rapid regulation of immune reactions. In vitro studies have confirmed the anti-inflammatory properties of a diet rich in cruciferous vegetables. Among others, reduced serum concentrations of pro-inflammatory cytokines, such as IL-1β, TNFα, and IL-6, were observed [50]. Several studies have confirmed that natural aryl hydrocarbon receptor (AhR) ligands are derived from food, such as broccoli. This AhR has been identified as one of the key immune receptors, and is expressed by not only different types of immune system cells, but also by certain types of cancer cells. The incorporation of the relevant ligands, including those found in cruciferous plants, results in activation, which transfers them to the nucleus. The resulting heterodimer exerts a number of different effects not only on the metabolic pathways, but also on genes whose expression modifies the course of immune reactions and the activity of various cytokines, including IL-22. The deficiency of the mentioned ligands has many negative consequences for the functioning of the whole organism. The digestive system is most exposed to these deficiencies, which manifest mainly in adverse effects on the bacterial intestinal flora, reduced granzyme production, and severe colitis [51]. Interestingly, mice studies have confirmed that a diet rich in cruciferous plants, which contain high levels of natural AhR ligands, mitigates these undesirable effects, as well as having a positive influence on the proper development of the immune system at an early stage of life. Isothionates can produce different compounds such as erucic and olic acids, which are considered to have strong antimicrobial effects. GSLs and their derivatives have also been reported to have chemopreventive actions [52].
Table 1. Health-promoting role of Glucosinolate.[53][54][55][56][57][58][59][60][61][62][63][64][65]
|
Glucosinolate |
Type of Compound |
Health Promoting Roles and Plant Protection |
Reference |
|
Sulforaphane |
Isothiocyanates |
Inhibition of transcription regulator (NF-κB), which is relevant against inflammation and for minimizing diabetes-related complications such as diabetic neuropathy. |
[13,53] |
|
Mediation of cell cycle arrest and apoptosis; inhibition of the activity of histone deacetylase; and increasing histone acetylation, which leads to the enhancement of protection against carcinogenesis. Induction of cytotoxicity. |
[54–56] |
||
|
Normalization of kidney genome and blood pressure owing to the decrease in oxidative stress in cardiovascular and kidney tissues. |
[57] |
||
|
Decreasing infarct size, brain edema, and cortical apoptosis, reducing the inflammation and tissue damage of the central nervous system due to the activation of the transcription factor Nrf2, and the upregulation of different target genes. |
[58] |
||
|
Reduction in the damage induced by high concentrations of substances that mimic the pathomechanism of autism spectrum disorders in mice models. |
[59] |
||
|
Treatment of Helicobacter pylori. |
[60] |
||
|
Reduces the risk of skin lesions caused by UV radiation, especially in high-risk patients. |
[44] |
||
|
Glucoiberin, Sinigrin, and Progoitrin |
Suppressing agents, protection of human and animal cells against carcinogenesis owing to the induction of Phase II detoxification enzymes or the inhibition of Phase I enzymes. |
[60–63] |
|
|
Indole-3-Carbinol |
Chemopreventive agent. |
[13] |
|
|
Benzyl Isothiocyanate |
Bactericidal and fungicidal properties and has proven effective in combating respiratory and urinary tract infections. |
[64] |
|
|
Chemopreventive agent. |
[13] |
||
|
Allyl Isothiocyanate, Allyl Thiocyanate, and Allyl Isocyanate |
Effective natural insecticides, efficiency in eliminating nematodes or flying insects. Possible mechanisms: the inhibition of the activity of the thiol groups of key enzymes, or the blocking of electron transport and ATP synthesis. |
[65] |
References
- Nauts, H.C. The Bibliography of Reports Concerning the Experimental Clinical Use of Coley Toxins. Cancer Research Institute: New York, NY, USA, 1985.
- Hadden, J.W. Immunostimulants. Trends Pharm. Sci. 1993, 14, 169–174.
- Upadhayay, U.P.P.D.D.; Ewam, P.C.V.V.; Ewam, U.P.C.V.V.; Sansthan, G.-A. Immunomodulatory and Therapeutic Potentials of Herbal, Traditional/Indigenous and Ethnoveterinary Medicines” Mahima,“Anu Rahal,” Rajib Deb,“Shyma, K. Latheef,” Hari Abdul Samad. Pak. J. Biol. Sci. 2012, 15, 754–774.
- Quintero-Fabián, S.; Ortuño-Sahagún, D.; Vázquez-Carrera, M.; López-Roa, R.I. Alliin, a garlic (Allium sativum) compound, prevents LPS-induced inflammation in 3T3-L1 adipocytes. Mediat. Inflamm. 2013, 2013.
- Tanaka, S.; Haruma, K.; Yoshihara, M.; Kajiyama, G.; Kira, K.; Amagase, H.; Chayama, K. Aged garlic extract has potential suppressive effect on colorectal adenomas in humans. J. Nutr. 2006, 136, 821S–826S.
- Fund, W.C.R.; Research, A.I. for C. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective. Amer Inst for Cancer Research: Washington, D.C., USA, 2007; Vol. 1; ISBN 0972252223.
- Munn, L.L. Cancer and inflammation. Wires Syst. Biol. Med. 2017, 9, 1370, doi:10.1002/wsbm.1370.
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545, doi:10.1016/S0140-6736(00)04046-0.
- Todoric, J.; Antonucci, L.; Karin, M. Targeting inflammation in cancer prevention and therapy. Cancer Prev. Res. 2016, 9, 895–905.
- Grimble, R.F. Interaction between nutrients, pro-inflammatory cytokines and inflammation. Clin. Sci. 1996, 91, 121–130, doi:10.1042/cs0910121.
- Grimble, R.F. Nutritional modulation of cytokine biology. Nutrition 1996, 14, 634–640, doi: org/10.1016/S0899-9007(98)00010-0.
- Grigore, A. Targeting Tumor-Associated Macrophages by Plant Compounds. In Macrophages; IntechOpen: London, UK, 2020.
- Fuentes, F.; Paredes-Gonzalez, X.; Kong, A.N.T. Dietary Glucosinolates Sulforaphane, Phenethyl Isothiocyanate, Indole-3-Carbinol/3,3′-Diindolylmethane: Antioxidative Stress/Inflammation, Nrf2, Epigenetics/Epigenomics and In Vivo Cancer Chemopreventive Efficacy. Curr. Pharm. Rep. 2015, 1, 179–196, doi: 10.1007/s40495-015-0017-y.
- Melino, S.; Sabelli, R.; Paci, M. Allyl sulfur compounds and cellular detoxification system: Effects and perspectives in cancer therapy. Amino Acids 2011, 41, 103–112.
- Antony, M.L.; Singh, S. V Molecular mechanisms and targets of cancer chemoprevention by garlic-derived bioactive compound diallyl trisulfide. Indian J. Exp. Biol. 2011, 49, 805.
- Ohkubo, S.; Dalla Via, L.; Grancara, S.; Kanamori, Y.; García-Argáez, A.N.; Canettieri, G.; Arcari, P.; Toninello, A.; Agostinelli, E. The antioxidant, aged garlic extract, exerts cytotoxic effects on wild-type and multidrug-resistant human cancer cells by altering mitochondrial permeability. Int. J. Oncol. 2018, 53, 1257–1268.
- Chang, H.-S.; Ko, M.; Ishizuka, M.; Fujita, S.; Yabuki, A.; Hossain, M.A.; Yamato, O. Sodium 2-propenyl thiosulfate derived from garlic induces phase II detoxification enzymes in rat hepatoma H4IIE cells. Nutr. Res. 2010, 30, 435–440.
- Modem, S.; DiCarlo, S.E.; Reddy, T.R. Fresh garlic extract induces growth arrest and morphological differentiation of MCF7 breast cancer cells. Genes Cancer 2012, 3, 177–186.
- Desai, G.; Schelske-Santos, M.; Nazario, C.M.; Rosario-Rosado, R. V.; Mansilla-Rivera, I.; Ramírez-Marrero, F.; Nie, J.; Myneni, A.A.; Zhang, Z.-F.; Freudenheim, J.L.; et al. Onion and Garlic Intake and Breast Cancer, a Case-Control Study in Puerto Rico. Nutr. Cancer 2020, 72, 791–800, doi:10.1080/01635581.2019.1651349.
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100.
- Franco, P.; Spinozzi, S.; Pagnotta, E.; Lazzeri, L.; Ugolini, L.; Camborata, C.; Roda, A. Development of a liquid chromatography—electrospray ionization—tandem mass spectrometry method for the simultaneous analysis of intact glucosinolates and isothiocyanates in Brassicaceae seeds and functional foods. J. Chromatogr. A 2016, 1428, 154–161.
- Hirani, A.H.; Li, G.; Zelmer, C.D.; McVetty, P.B.E.; Asif, M.; Goyal, A. Molecular genetics of glucosinolate biosynthesis in Brassicas: Genetic manipulation and application aspect. Crop. Plant. 2012, 189–216.
- Bischoff, K.L. Glucosinolates. In Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2016; pp. 551–554.
- Mikkelsen, M.D.; Naur, P.; Halkier, B.A. Arabidopsis mutants in the C-S lyase of glucosinolate biosynthesis establish a critical role for indole-3-acetaldoxime in auxin homeostasis. Plant. J. 2004, 37, 770–777.
- Ishida, M.; Nagata, M.; Ohara, T.; Kakizaki, T.; Hatakeyama, K.; Nishio, T. Small variation of glucosinolate composition in Japanese cultivars of radish (Raphanus sativus L.) requires simple quantitative analysis for breeding of glucosinolate component. Breed. Sci. 2012, 62, 63–70.
- Lee, M.-K.; Chun, J.-H.; Byeon, D.H.; Chung, S.-O.; Park, S.U.; Park, S.; Arasu, M.V.; Al-Dhabi, N.A.; Lim, Y.-P.; Kim, S.-J. Variation of glucosinolates in 62 varieties of Chinese cabbage (Brassica rapa L. ssp. pekinensis) and their antioxidant activity. Lwt-Food Sci. Technol. 2014, 58, 93–101.
- Li, G.; Quiros, C.F. In planta side-chain glucosinolate modification in Arabidopsis by introduction of dioxygenase Brassica homolog BoGSL-ALK. Appl. Genet. 2003, 106, 1116–1121.
- Halkier, B.A.; Gershenzon, J. Biology and biochemistry of glucosinolates. Annu. Rev. Plant. Biol. 2006, 57, 303–333.
- Collett, M.G.; Stegelmeier, B.L.; Tapper, B.A. Could nitrile derivatives of turnip (Brassica rapa) glucosinolates be hepato-or cholangiotoxic in cattle? J. Agric. Food Chem. 2014, 62, 7370–7375.
- Ahuja, I.; Rohloff, J.; Bones, A.M. Defence mechanisms of Brassicaceae: Implications for plant-insect interactions and potential for integrated pest management. A review. Agron. Sustain. Dev. 2010, 30, 311–348.
- Nair, A.; Chattopadhyay, D.; Saha, B. Plant-derived immunomodulators. In New Look to Phytomedicine; Elsevier: Amsterdam, The Netherlands, 2019; pp. 435–499.
- Jantan, I.; Ahmad, W.; Bukhari, S.N.A. Plant-derived immunomodulators: An insight on their preclinical evaluation and clinical trials. Front. Plant. Sci. 2015, 6, 655.
- Marszałek, K.; Woźniak, Ł.; Skąpska, S.; Mitek, M. A Comparative Study of the Quality of Strawberry Purée Preserved by Continuous Microwave Heating and Conventional Thermal Pasteurization During Long-Term Cold Storage. Food Bioprocess. Technol. 2016, 9, 1100–1112, doi:10.1007/s11947-016-1698-x.
- Hallmann, E.; Kazimierczak, R.; Marszałek, K.; Drela, N.; Kiernozek, E.; Toomik, P.; Matt, D.; Luik, A.; Rembiałkowska, E. The Nutritive Value of Organic and Conventional White Cabbage (Brassica Oleracea, L. Var. Capitata) and Anti-Apoptotic Activity in Gastric Adenocarcinoma Cells of Sauerkraut Juice Produced Therof. J. Agric. Food Chem. 2017, 65, 8171–8183, doi:10.1021/acs.jafc.7b01078.
- Shin, I.-S.; Hong, J.; Jeon, C.-M.; Shin, N.-R.; Kwon, O.-K.; Kim, H.-S.; Kim, J.-C.; Oh, S.-R.; Ahn, K.-S. Diallyl-disulfide, an organosulfur compound of garlic, attenuates airway inflammation via activation of the Nrf-2/HO-1 pathway and NF-kappaB suppression. Food Chem. Toxicol. 2013, 62, 506–513.
- Colín-González, A.L.; Santamaría, A. Garlic, gastrointestinal protection and oxidative stress. In Gastrointestinal Tissue; Elsevier: Amsterdam, The Netherlands, 2017; pp. 275–288.
- Florkiewicz, A.; Ciska, E.; Filipiak-Florkiewicz, A.; Topolska, K. Comparison of Sous-vide methods and traditional hydrothermal treatment on GLS content in Brassica vegetables. Eur. Food Res. Technol. 2017, 243, 1507–1517.
- Kapusta-Duch, J.; Kusznierewicz, B.; Leszczyńska, T.; Borczak, B. Effect of cooking on the contents of glucosinolates and their degradation products in selected Brassica vegetables. J. Funct. Foods 2016, 23, 412–422.
- Kim, S.R.; Jung, Y.R.; An, H.J.; Kim, D.H.; Jang, E.J.; Choi, Y.J.; Moon, K.M.; Park, M.H.; Park, C.H.; Chung, K.W. Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-κB signaling. Plos One 2013, 8, e73877.
- Westphal, A.; Riedl, K.M.; Cooperstone, J.L.; Kamat, S.; Balasubramaniam, V.M.; Schwartz, S.J.; Böhm, V. High-pressure processing of broccoli sprouts: Influence on bioactivation of glucosinolates to isothiocyanates. J. Agric. Food Chem. 2017, 65, 8578–8585.
- Fu, E.; Tsai, M.; Chin, Y.; Tu, H.; Fu, M.M.; Chiang, C.; Chiu, H. The effects of diallyl sulfide upon P orphyromonas gingivalis lipopolysaccharide stimulated proinflammatory cytokine expressions and nuclear factor-kappa B activation in human gingival fibroblasts. J. Periodontal Res. 2015, 50, 380–388.
- Formica-Oliveira, A.C.; Martínez-Hernández, G.B.; Díaz-López, V.; Artés, F.; Artés-Hernández, F. Use of postharvest UV-B and UV-C radiation treatments to revalorize broccoli byproducts and edible florets. Innov. Food Sci. Emerg. Technol. 2017, 43, 77–83.
- Zhou, Y.; Zhuang, W.; Hu, W.; Liu, G.; Wu, T.; Wu, X. Consumption of large amounts of Allium vegetables reduces risk for gastric cancer in a meta-analysis. Gastroenterology 2011, 141, 80–89.
- Palliyaguru, D.L.; Yuan, J.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the power of plants to people. Mol. Nutr. Food Res. 2018, 62, 1700965.
- Shapiro, T.A.; Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Talalay, P. Chemoprotective glucosinolates and isothiocyanates of broccoli sprouts: Metabolism and excretion in humans. Cancer Epidemiol. Prev. Biomark. 2001, 10, 501–508.
- Zhang, Y.S.; Kolm, R.H.; Mannervik, B.; Talalay, P. Reversible conjugation of isothiocyanates with glutathione catalyzed by human glutathione transferases. Biochem. Biophys. Res. Commun. 1995, 206, 748–755.
- Combourieu, B.; Elfoul, L.; Delort, A.-M.; Rabot, S. Identification of new derivatives of sinigrin and glucotropaeolin produced by the human digestive microflora using 1H NMR spectroscopy analysis of in vitro incubations. Drug Metab. Dispos. 2001, 29, 1440–1445.
- Navarro, J.L.; Tárrega, A.; Sentandreu, M.A.; Sentandreu, E. Partial purification and characterization of polyphenol oxidase from persimmon. 2014, 157, 283–289, doi:10.1016/j.foodchem.2014.02.063.
- Wagner, A.E.; Terschluesen, A.M.; Rimbach, G. Health promoting effects of brassica-derived phytochemicals: From chemopreventive and anti-inflammatory activities to epigenetic regulation. Oxid. Med. Cell. Longev. 2013, 2013.
- Thejass, P.; Kuttan, G. Inhibition of angiogenic differentiation of human umbilical vein endothelial cells by diallyl disulfide (DADS). Life Sci. 2007, 80, 515–521, doi:https://doi.org/10.1016/j.lfs.2006.09.045.
- Abdull Razis, A.F.; De Nicola, G.R.; Pagnotta, E.; Iori, R.; Ioannides, C. A glucosinolate-rich extract of Japanese Daikon perturbs carcinogen-metabolizing enzyme systems in rat, being a potent inducer of hepatic glutathione S-transferase. 2013, 52, 1279–1285.
- Cavaiuolo, M.; Ferrante, A. Nitrates and glucosinolates as strong determinants of the nutritional quality in rocket leafy salads. Nutrients 2014, 6, 1519–1538.
- Negi, G.; Kumar, A.; S Sharma, S. Nrf2 and NF-κB modulation by sulforaphane counteracts multiple manifestations of diabetic neuropathy in rats and high glucose-induced changes. Curr. Neurovasc. Res. 2011, 8, 294–304.
- Gamet-Payrastre, L. Signaling pathways and intracellular targets of sulforaphane mediating cell cycle arrest and apoptosis. Curr. Cancer Drug Targets 2006, 6, 135–145.
- Dinkova-Kostova, A.T. Chemoprotection against cancer by isothiocyanates: A focus on the animal models and the protective mechanisms. In Natural Products in Cancer Prevention and Therapy; Springer: Berlin, Germany, 2012; pp. 179–201.
- Sturm, C.; Wagner, A.E. Brassica-derived plant bioactives as modulators of chemopreventive and inflammatory signaling pathways. Int. J. Mol. Sci. 2017, 18, 1890.
- Senanayake, G.V.K.; Banigesh, A.; Wu, L.; Lee, P.; Juurlink, B.H.J. The dietary phase 2 protein inducer sulforaphane can normalize the kidney epigenome and improve blood pressure in hypertensive rats. Am. J. Hypertens. 2012, 25, 229–235.
- Tanito, M.; Masutani, H.; Kim, Y.-C.; Nishikawa, M.; Ohira, A.; Yodoi, J. Sulforaphane induces thioredoxin through the antioxidant-responsive element and attenuates retinal light damage in mice. Invest. Ophthalmol. Vis. Sci. 2005, 46, 979–987.
- Lynch, R.; Diggins, E.L.; Connors, S.L.; Zimmerman, A.W.; Singh, K.; Liu, H.; Talalay, P.; Fahey, J.W. Sulforaphane from broccoli reduces symptoms of autism: A follow-up case series from a randomized double-blind study. Glob. Adv. Heal. Med. 2017, 6, 2164957 × 17735826, doi: 10.1177/2164957X17735826
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229.
- Feng, Y.; Zhu, X.; Wang, Q.; Jiang, Y.; Shang, H.; Cui, L.; Cao, Y. Allicin enhances host pro-inflammatory immune responses and protects against acute murine malaria infection. Malar. J. 2012, 11, 268.
- Nok, A.J.; Williams, S.; Onyenekwe, P.C. Allium sativum-induced death of African trypanosomes. Parasitol. Res. 1996, 82, 634–637.
- Coppi, A.; Cabinian, M.; Mirelman, D.; Sinnis, P. Antimalarial activity of allicin, a biologically active compound from garlic cloves. Antimicrob. Agents Chemother. 2006, 50, 1731–1737.
- Drobnica, Ľ.; Zemanova, M.; Nemec, P.; Antoš, K.; Kristian, P.; Štullerová, A.; Knoppova, V. Antifungal Activity of Isothiocyanates and Related Compounds: I. Naturally Occurring Isothiocyanates and Their Analogues. Appl. Microbiol. 1967, 15, 701–709.
- Tsao, R.; Peterson, C.J.; Coats, J.R. Glucosinolate breakdown products as insect fumigants and their effect on carbon dioxide emission of insects. Bmc Ecol. 2002, 2, 5.