Many species of the so-called exotic plants coexist with native species in a balanced way, but others thrive very quickly and escape human control, becoming harmful—these are called invasive alien species. In addition to overcoming geographic barriers, these species can defeat biotic and abiotic barriers, maintaining stable populations. Ailanthus altissima is no exception; it is disseminated worldwide and is considered high risk due to its easy propagation and resistance to external environmental factors. Currently, it has no particular use other than ornamental, even though it is used to treat epilepsy, diarrhea, asthma, ophthalmic diseases, and seborrhoea in Chinese medicine. Considering its rich composition in alkaloids, terpenoids, sterols, and flavonoids, doubtlessly, its use in medicine or other fields can be maximised.
1. Introduction
Since ancient times humans have taken advantage of the existing flora, especially trees, for various food, wood and non-wood products. Over the years, trees have been planted to provide these services. Consequently, humans’ ability to plant trees and transport goods worldwide has led to a greater expression of non-native trees
[1]. The spread of many non-native tree species has shown in recent years an increase in the number of publications highlighting its potential constraints either regionally or globally (e.g.,
Eucalyptus globulus [2] and
Acacia spp.
[3]). Other studies have also noted the conflicts that some invasive alien species (IAS), including some non-native species, can cause in the ecosystem
[4,5][4][5].
IAS are recognised for triggering various impacts, such as habitat modification, the alteration of community structure, and affecting ecosystem processes. Thus, they are considered one of the most significant global threats to biodiversity. However, these species can also be quite helpful to humans, providing a complex number of services, which makes it difficult to assess their positive and negative effects. For this reason, the importance of evaluating and investigating some IAS that have several adverse effects has been growing
[6].
The IAS
Ailanthus altissima (Mill.) Swingle, known as the “Tree of Heaven”, was introduced into Europe and worldwide in the 18th century. This plant spreads and is distributed mainly in cities, agricultural fields, and transportation corridors. In cities, it was reported to cause problems from its roots such as damaging infrastructures plus allergic reactions and respiratory problems in humans. It is more frequent in the southern and submeridional zones whereas, in Europe, it has a closed distribution area in the Mediterranean
[7]. Through this area, there can be modifications in the structure of the local vegetation and damage to the stability of the ecosystem
[8]. In some warm regions of Europe,
A. altissima can also cause damage to walls and other structures, as reported by Almeida et al.
[9], where this species appeared and spread on a wall of a historic monument in Coimbra, Portugal.
Nowadays, this species has been the subject of several pharmacological investigations, showing its positive effects. Traditional Chinese medicine uses it to treat certain disorders such as epilepsy, diarrhea, asthma, ophthalmic diseases, and seborrhoea. Traditional medicine practitioners use different parts of the plant for different types of health problems. The bark, for example, is prescribed for dysentery, menorrhagia and spermatorrhea. For intestinal problems that last a few months, it is advisable to boil some bark with water and then drink the liquid together with gin. Other bark remedies can be made with other ingredients and plants, such as root onions and Chinese pepper (
Zanthoxylum simulans) for rectal problems, especially after childbirth. In addition, it can also be used as an astringent, parasiticide, a narcotic substance, and a drug to relieve spasms
[6,7][6][7]. On the other hand,
A. altissima may have applications in agriculture as an environmentally friendly compound, since its extracts have strong herbicidal and insecticidal properties
[10].
2. Taxonomy and Morphology
Ailanthus altissima is an accepted name with original publication details in J. Wash. Acad. Sci. 6: 495 (1916) and many synonyms
[11]. It belongs to the family Simaroubaceae
[6], which consists mainly of trees and shrubs, distributed in tropical and warm regions, comprising 30 genera and 150 species. The term for the genus
Ailanthus is an Ambonese word derived from ailanto, meaning “tree of heaven”
[12]. About 5–10 species of trees from this genus are known and characterised by being deciduous with rapid growth, large spreading branches, pinnate leaves with pointed leaflets, where the terminal leaflet is usually present, small yellow to greenish flowers, and the fruits samaras stretched into a long wing, where the seed lies in the middle (
Figure 1C). In the root sprouts, the emerging leaves vary in the number of divisions (from unifoliate to compound pinnately) and are yellowish-green at first. In addition, the genus
Ailanthus has a fine, satiny wood and is known for its medicinal values
[12].
Figure 1. Some examples of A. altissima in the Castelo Branco region (Portugal); (A) an old plant; (B) a younger plant; (C) the samaras with seeds in the middle; (D) the trunk of the first plant and (E) part of a leaf with its leaflets.
Regarding the morphology of the species in the study,
A. altissima (
Figure 1) is described as a tree growing up to 30 m in height with a trunk of greyish, smooth, or cracked bark, and in the oldest specimens and branches, a reddish-brown colour. Its leaves (30–100 cm) are alternate, compound, odd-pinnate (on the shoots) and paripinnate (arranged at the ends of the branches), and glabrous or with scattered hairs on the upper side of the margin. Each leaf can have 5–12 pairs of leaflets (or 4 to 35 leaflets
[13]) of very variable size, petiolate, from narrowly lanceolate to ovate-lanceolate, with gradually narrowed apex, and an entire margin. Each of these leaflets has two to four glandular teeth near the base, with 1–8 openings as extrafloral nectaries—an important characteristic to identify
A. altissima [6,13,14][6][13][14].
A. altissima is a tree with both female and male terminally located inflorescences, where the latter are larger, pluriflor, and emit an unpleasant scent. In addition, unlike the female flowers, the male flowers have 10 well-functioning stamens, each with a fertile anther; the female flowers have abortive anthers. It is due to these facts that
A. altissima is considered a dioecious tree
[7,13,15][7][13][15]. Flowers have a tiny cupular and five-lobed calyx, a corolla with five sepals, lanceolate and green, five distinct petals that are hairy at the base, greenish to yellowish-green, and an annular and lobed gland disc (different types of nectaries); female flowers have five free carpels that generally develop into five separate winged fruits. The fruits (samaras) can vary from greenish-yellow to reddish-brown, and are arranged in clusters and dispersed by the wind
[6,7,13,14][6][7][13][14]. In the samaras, the seeds are found and these, depending on the location of the trees, vary considerably in colour, size, weight, and thickness
[6].
Concerning roots, the main root can develop several lateral roots, from pre-existing primordia or root sprouts. The root system has a highly variable and asymmetric spatial extent, in which lateral roots have been reported to reach a length of 27 m from the parent tree. The nectaries’ primary function is to eliminate excess sugar from the plant.
A. altissima has floral and extrafloral nectaries, which excrete different forms of sugar until October. These nectaries are located on the leaves, pseudostipules, and cataphylls. Trees of the genus
Ailanthus have a rapid stem elongation, which leads to the fact that these trees can grow to heights of 25–45 m
[7,12][7][12].
3. Biology and Ecology
According to Kowarik and Säumel’s review
[7],
A. altissima is a short-lived tree that can reach more than 100 years. These authors also mentioned the existence of trees in Germany that were 130, 121, and 113 years old. In terms of germination and seed establishment, information varies widely in the literature. This species is characterised by not forming a long-term seed bank but can establish temporary seed banks in the soil. It has also been observed that a seed in contact with water influences its initiation, duration, and germination rate. In addition, the germination rate was reported to decrease with altitude. The same was observed on a germination experiment when comparing soil types (sand with gravel and peat substrates). The germination rate is much lower on sand and higher on peat substrates
[7]. Considering seed establishment, the combination of litter, weed competition, and insect herbivory were studied. It was concluded that litter (without weed competition) delays germination but does not affect seedling biomass.
On the other hand, with weed competition, seedling growth was reduced but neutralised by the litter. However, the litter showed damage to the seedling performance by increasing herbivory
[19,20][16][17]. In terms of stem elongation,
A. altissima is one of the fastest-growing trees in North America and Great Britain. The height and diameter increase until the trees reach 5 to 10 years, and then from 10 to 20 years, it starts to decrease. In Hu’s
[13] review, he states that as young as 1 year old, seedlings can reach 1–2 m in height. On average, the height increment in trees between 20 and 25 years can be less than 8 cm per year
[7].
A. altissima is a different tree from many other tree species. One of the requirements for the buds to open is high temperature, leading the seasonal development to start later and last longer (until late fall)
[7]. As mentioned before, altitude influences the germination rate and in the case of the flowering stage this is also an essential factor to take into account. The flowers appear at different stages depending on altitude. In North America, they appear from mid-April to July; in the French Mediterranean, they start in mid-May; and in Central Europe, flowering occurs mainly in July. The ripening occurs in September-October, and soon after this, there is the abscission of the samaras, which can vary in time depending on the individual and the years. The fruits can be almost all released in some years before the end of February, and in others, a large part of them can remain on the tree until the beginning of May
[6,7][6][7].
The reproduction of
A. altissima occurs by seeds and by root suckers. In sexual reproduction, the seedlings give rise to flowers after 3 weeks of germination and large amounts of light. The flowers usually develop non-viable seeds, at this stage. However,
Ailanthus is pollinated by bees, beetles, and other insects
[7].
In vegetative reproduction, what occurs is the growth of new shoots from a mother plant. These shoots are known to have a great length and can reach up to 27 m or more. Alternatively,
A. altissima can propagate itself through vegetative regeneration. It consists of originating shoots from pre-existing buds in the hypocotyl, axillary buds of the cataphylls, the roots, and adventitious buds on a cutting section
[6,7][6][7]. The dispersal of
A. altissima seeds, which give rise to new individuals, is achieved through the dispersal of samaras. This dispersion can occur from the following types of vectors: the wind, in which the samaras are moved individually or aggregated in groups by water (through river corridors) and animals (such as rodents and birds)
[7].
The response to ecological factors is another critical issue to mention when it comes to this IAS. It was found that
A. altissima is tolerant to several abiotic factors, contributing to its extensive spread. Among the several factors, the following stand out: temperature, frost, drought, soils, and air pollution
[6]. First, this species is temperature tolerant, although it adapts better to high temperatures, equivalent to its distribution. Regarding low temperatures, it was observed in a study that old trees survived at −33 °C; however, they suffered some damage. Kowarik and Säumel also mentioned that trees in the early stages of life might be more susceptible to the cold
[7]. Drought is another factor to which
Ailanthus is tolerant, presenting mechanisms that contest the lack of water, such as the closing of the stomata and the reallocation of food reserves to the lateral roots from the main root. Another fundamental factor is the soil.
A. altissima grows in various soil types: from barren, rocky soils to saline and alkaline soils; it also tolerates dry, humid, and nutrient-rich or nutrient-poor soils.
A. altissima’s tolerance to pollution is attributed to the high antioxidant capacity of the leaves. It is pretty tolerant of major components of air pollution such as SO
2. However, it is sensitive to ozone
[7,21][7][18] and cold, which can inhibit some seedlings and limit their occurrence and distribution.
Furthermore, this species of
Ailanthus has allelopathic effects on other plant species by producing a wide variety of active compounds and becoming resistant to herbivores and pathogens. All these ecological factors and the biological characteristics of
A. altissima, make it a very competitive species and justify its wide distribution worldwide. Due to its tolerance to several factors, it is established in different habitats. It is widely dispersed and able to establish seedlings at long distances
[6].
4. Chemical Characterization
Plants of the
Ailanthus genus are phytochemically characterised by being rich in alkaloids, terpenoids, sterols, flavonoids, and other compounds. These compounds nowadays are widely studied since they are responsible for several pharmacological activities
[12]. The bark of
A. altissima has always been used in traditional medicine and, nowadays, it is known for being very rich in several compounds. It contains oleoresin, resin, ceryl alcohol, ailanthin, isoquercetin, tannins, ceryl palmitate, and saponins
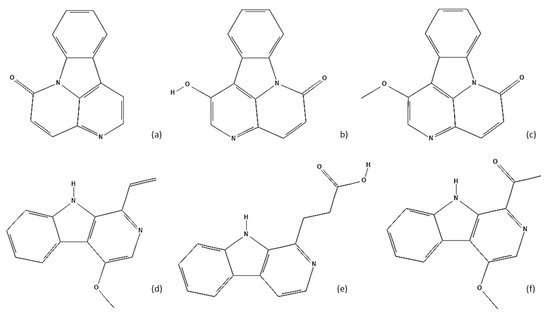
[7]. Two hundred and twenty-one compounds have been isolated and identified from the dry bark, such as alkaloids, quassinoids, phenylpropanoids, triterpenoids, and volatile oils. The main active compounds of dry bark are alkaloids (
Figure 2), 32 of which have already been isolated. Quassinoids (
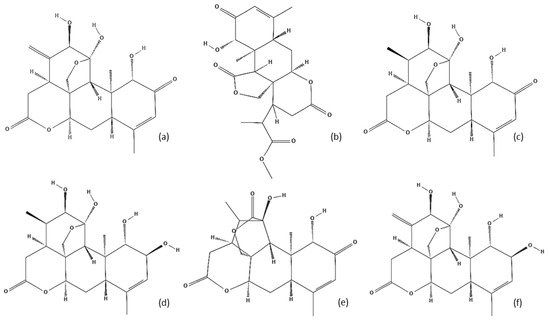
Figure 3) are also one of the characteristic compounds of dry bark, with 40 isolated to date
[22][19]. As reported by Pijush Kundu
[12], the root bark essentially has alkaloids and terpenoids. Some ligands, coumarins, phenylpropanoids, and new terpenoids have also been identified and isolated
[12,23,24,25][12][20][21][22]. The leaves of this species are characterised by having some percentage of tannins, quercetin, isoquercetin, alkaloids, and mainly flavonoids. These were widely used in traditional Indian medicine to treat seborrhoea and scabies
[7,12][7][12].
Figure 2. Some examples of alkaloids identified in A. altissima: (a) Canthin-6-one; (b) 1-Hydroxycanthin-6-one; (c) 1-Methoxycanthin-6-one; (d) 4-methoxy-1-vinyl-β-carboline; (e) β-carboline-1-propionic acid; (f) 1-Acetyl-4-methoxy-β-carboline.
Figure 3. Some examples of the main A. altissima quassinoids identified: (a) Ailanthone; (b) Ailantinol A; (c) Chaparrinone; (d) Chaparrin; (e) Shinjudilactone; (f) Shinjulactone A.
Regarding the fruits of
A. altissima, not much is known about its constituents, but they was widely used in China as a medicine for bleeding and as an antibacterial. On the other hand, in India it was used as an emmenagogue and to treat eye diseases
[12,26][12][23]. Jian-Cheng Ni et al.
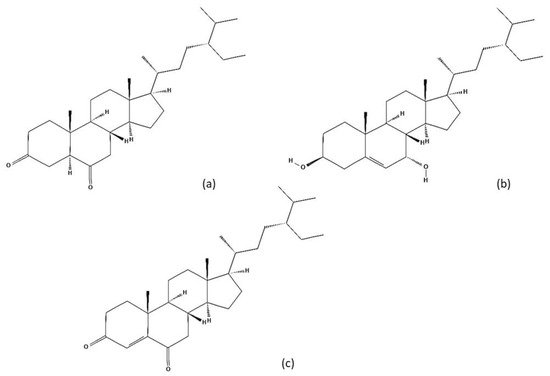
[27][24] mentioned that previous studies have shown that only four quassinoid glycosides and several stigmasterols (
Figure 4)
[28][25] were identified from fruits. However, they elucidated the structure and isolated four new compounds: two phenylpropionamides, piperidine, and a phenolic derivative. In addition to these new compounds, 13 phenols, 10 flavonoids, and a phenylpropionamide were also isolated. According to Clair and Bory
[29][26], the composition of
A. glandulosa (syn. of
A. altissima) extrafloral nectar diverge according to the nectary type. These authors also declared that the three essential nectars’ sugars are sucrose, fructose, and glucose in the leaf nectaries. It is the high amount of fructose that characterizes this species. Furthermore, bound lipids (monogalactosyldiacylglycerol) and oleic, palmitic, and linoleic acids have been isolated in the secretion of glandular trichomes in cataphylls and in young stems
[7,29][7][26].
Figure 4. Some examples of steroids identified in A. altissima: (a) 5α-Stigmastane-3,6-dione; (b) Stigmast-5-ene-3β,7α-diol; (c) Stigmast-4-ene-3,6-dione.
The quassinoids class has been the main target of many studies due to their bioactivities and phytotoxic impacts. The most widely studied active compound in this group of terpenoids is ailanthone. This has been identified as an effective phytotoxic agent, capable of being used as a herbicide
[7,30][7][27]. In addition to ailantthone, other derivatives have been isolated and studied (such as ailanthione, ailanthinol B, and chaparrin). There is an increasing demand for the isolation of new quassinoid derivatives to investigate their potential applications, mainly in pharmacology
[31,32,33][28][29][30].
Table 1 summarises the main compounds by classes of compounds and which parts of the plants were used to identify the chemical compounds. These results are based on the review by Pijush Kundu and Subrata Laskar
[12].
Table 1. The main compounds identified in A. altissima grouped by the major classes and which part of the plant was used for their isolation.
| |
Compounds |
Parts Used (Source) |
| Alkaloids |
1-Ethyl-4-methoxy-β-carboline |
Root bark |
| β-Carboline-1-propionic acid |
Root bark |
| Methyl-4-methoxy-β-caboline carboxylate |
Root bark |
| 1-(1′-2′-Dihydroxyethyl)-4-methoxy-β-carboline |
Root bark |
| 1-(2′-Hydroxyethyl)-4-methoxy-β-carboline/Crenatine |
Root bark |
| 4-Methoxy-1-vinyl-β-carboline/Dehydrocrenatine |
Dried leaves |
| 1-Methoxycarbonyl-4,8-dimethoxy-β-caboline |
Leaves |
| 1-Methoxycarbonyl-β-carboline/1-caromethoxy-β-carboline |
Leaves |
| 1-(1-Hydroxy-2-methoxy)-ethyl-4-methoxy-β-carboline |
Root bark |
| 1-Carbamoyl-β-carboline |
Root bark |
| 1-Acetyl-4-methoxy-β-carboline |
Root bark |
| Canthine-6-one |
Root bark, leaves and wood |
| 1-Hydroxycanthine-6-one |
Root bark |
| 1-Methoxy-canthine-6-one |
Root bark, dried leaves and wood |
| 5-Hydroxymethylcanthine-6-one |
Root bark |
| Canthine-6-one-3-N-oxide |
Wood |
| 1-Methoxycanthine-6-one-3-N-oxide |
Root bark |
| Terpenoids |
Ailanthone |
Stem bark |
| Chapparinone |
Seed |
| Chapparin |
Aerial part |
| Shinjulactone and Shinjulactone B |
Stem bark |
| Shinjulactone A |
Seed |
| Shinjulactone C, D, E, F, M, N, G, H, I, J, K and L |
Root bark |
| ∆ | 13(18) | -Dehydroglaucarubinone |
Stem bark |
| ∆ | 13(18) | -Dehydroglaucarubolone |
Stem bark |
| Ailantinol A e B |
Stem bark |
| Ailantinol C, D, E, F, G and H |
Aerial part |
| Shinjuglycoside A, B, C and D |
Seed |
| Shinjuglycoside E and F |
Root bark |
| Shinjudilactone |
Root bark |
| Cycloart-25-ene-3β-24R-diol and Cycloart-25-ene-3β-24S-diol |
Fruits |
| 9,19-Cyclolanost-23(Z)ene-3β,25-diol |
Fruits |
| Ailantholide |
Seed |
| 3-epi-ursolic acid |
Fruits |
| 12β, 20 (S)-dihydroxy dammar-24-en-3-one |
Fruits |
| Steroids |
β-Sitosterol |
Leaves and fruits |
| β-Sitosterol glucoside/Daucosterol/Sitosterol-3-Oβ-D-glucoside |
Leaves and fruits |
| Ailanthusterol A and B |
Seed |
| 5α-Stigmastane-3,6-dione |
Fruits |
| 5α-Stigmastane-3β, 6β-diol |
Fruits |
| 6α-Hydroxy-stigmast-4-en-3-one |
Fruits |
| Stigmast-4-ene-3β, 6α-diol |
Fruits |
| Stigmast-4-ene-3,6-dione |
Fruits |
| 6β-Hydroxy-stigmast-4-ene-3-one |
Fruits |
| Stigmast-4-ene-3β, 6β-diol |
Fruits |
| 3β-Hydroxy-stigmast-5-en-7-one |
Fruits |
| Stigmast-5-ene-3β, 7α-diol |
Fruits |
| Stigmast-5-ene-3β, 7α, 20ζ-triol |
Fruits |
| Flavonoids |
Quercetin |
Leaves |
| Kaempferol |
Leaves |
| Isoquercetin/Quercetin-3-O-glucoside |
Leaves |
| Kaempferol-3-O-glucoside |
Leaves |
| Rutin |
Leaves |
| Luteolin-7-O-β-(6" galloylglucopyranoside) |
Leaves |
| Miscellaneous compounds |
1-O-β-D-Glucopyranosyl-(2S,3R,4E,9E)-2-(2'-R-hydroxyhexadecenoy)-4,9-octadecadiene-1,3-diol |
Fruits |
| Ceryl alcohol |
Leaves |
| Ethyl gallate |
Leaves |
| Altissimacoumarin A and B |
Bark |
| Coumarin |
Bark |
| Isofraxidin |
Bark |
| Scopoletin |
Bark |
5. Biological Properties
For a long time, in the history of science, plants and parts of them were explored and used for medicinal purposes.
A. altissima offers a promising natural alternative for food safety and bioconservation as well as for its antioxidant properties. Several studies report the pharmacological effects: antimicrobial (antibacterial and antiviral), antioxidant, cytotoxic, anti-inflammatory, antipyretic, analgesic, anti-progestogenic, and many others
[34,35,36,37][31][32][33][34].
5.1. Antimicrobial Activity
Research on the bactericidal efficiency of phytochemicals as viable alternatives for chemical antibiotics has been conducted. An essential feature of extracts from plant origin is their antimicrobial activity, which contributes to alternative synthetic antibiotics
[38][35].
Natural products derived from
A. altissima may contribute to the development of new antimicrobial agents used as growth inhibitors of
Listeria monocytogenes,
Staphylococcus aureus,
Bacillus subtilis,
Escherichia coli,
Pseudomonas aeruginosa, and some important foodborne pathogens and spoilage bacteria
[39][36].
Methanolic extracts from leaves and hydrodistilled residues were strong and efficient against Gram-positive bacteria:
S. aureus,
B. subtilis,
Enterococcus faecium, and
Streptococcus agalactiae [40][37]. The same was reported by Rahman
[39][36]; methanolic extracts of
A. altissima leaves were most effective against Gram-positive bacteria, namely
L. monocytogenes (ATCC 19,116, ATCC 19,118 and ATCC 19,166),
S. aureus (ATCC 6538 and KCTC 1916), and
B. subtilis (ATCC 6633), and two Gram-negative bacteria,
P. aeruginosa (KCTC 2004) and
E. coli (ATCC 8739). The zones of inhibition of methanol extract and its derived different polar subfractions against the tested bacteria were found in the 12.1–23.2 mm range, and the minimum inhibitory concentration (MIC) values were recorded between 62.5 and 500 mg/mL
[39][36]. However, their extracts were ineffective against Gram-negative bacteria, such as
E. coli (ATCC 43,888),
Salmonella enteritidis (KCTC 12,021), and
Salmonella typhimurium (ATCC 2525). The MIC values of the methanolic extract against the tested bacteria were found in the 125–500 mg/mL range. In addition, the MeOH extract and its ethyl acetate (EtOAc) subfraction were compared with the standard antibiotics, tetracycline and streptomycin; in some cases, these showed greater antibacterial activity compared to streptomycin, but in other cases tetracycline showed greater activity than the solvent fractions. Aissani et al.
[41][38] observed in aqueous and methanolic extracts of the bark and wood strong antimicrobial activity against
P. aeruginosa (ATCC 9027) strains and isolated strains with an inhibition zone of 12 ± 0.3 mm. Zhao et al.
[28][25] showed that the extract from fruits was weakly active against
E. coli,
S. aureus,
P. aeruginosa, and
S. typhiuriun, with inhibition zones of 6.87–7.51 mm, using the concentration of ethanol extract of 1.2 mg/mL. The plant’s chemical composition can be variable due to variations in origin, species, growth, harvesting, and processing conditions, thus altering the biological activities.
Several studies show that antibacterial activity can be attributed to the occurrence of some specific components such as total polyphenol contents
[42][39], namely gallic acid, rutin
[43][40], and epicatechin
[44][41]. The resistance of Gram-positive bacteria towards plant extracts has been previously reported
[44,45,46][41][42][43]. The results obtained encourage the use of species as a food preservative and for pharmaceutical purposes.
5.2. Antioxidant Activity
Scientific research is interested in quantifying and using antioxidants mainly due to their potent biological activity.
[47][44]. Although antioxidant compounds may be synthesized, there is a growing interest in the natural compounds from different plants
[48,49][45][46]. Taking this into account, Luis et al.
[50][47] performed a research to determine the phenolic, flavonoid, and total alkaloid content on four different extracts (methanolic, ethanolic, hydroalcoholic, and acetone) of
A. altissima (steams, stalks, and leaves) to establish a correlation to the antioxidant activity of these extracts. To assess their antioxidant activity, DPPH (2,2-diphenyl-1-picryl-hydrazyl-hydrate) assay and the ϐ-carotene bleaching test were used.
It was possible to quantify the total amount of phenolic compounds, 268.15 mg/g of dry extract in all extracts. The most important finding of this research is the potential antioxidant properties of
A. altissima, since it is abundant in phenolic, flavonoid, and alkaloid compounds. Lungu et al.
[51][48] have identified the presence of flavonoids (ranging around 5.34 to 5.41 g of rutin equivalents per 100 g of dry weight) in leaf extracts of
A. altissima. The extraction method by reflux or ultrasound did not have a significant effect on the final concentration.
Moreover, the antioxidant activity was performed by the DPPH assay and the 2,2-AzinoBis (3-ethylbenzthiazoline-6-Sulphonic acid) (ABTS) radical scavenging assay. Once again, the extraction method did not affect the obtained concentrations, meaning that values are approximately 0.16 mM Trolox equivalent of extract for DPPH and around 9 mM Trolox equivalent when ABTS was used. In summary, results showed that flavonoids were in higher amounts than other compounds and are responsible for the antioxidant properties of the ethanolic leaf extracts
[51][48]. Another study with leaves from different localities in Tunisia found that the main phenolic compounds in methanolic extracts were gallic acid, chlorogenic acid, HHDP-galloylglucose, epicatechin, rutin, hyperoside, and quercetin-3-galloyl hexoside
[40][37]. Once again, the typical antioxidant activity assays (DPPH and ABTS) were used, as well as the ferric reducing antioxidant power (FRAP) assay and the 2-deoxyribose method, which was used for the determination of the scavenging effects of the methanolic extracts on hydroxyl (OH) radicals. Results showed good antioxidant activity of the methanolic extracts.
Altogether, leaves from the Bousalem region showed better antioxidant activity than those from other regions, with EC
50 values of 14.78 μg/mL, 8.64 μg/mL, and 4.42 μg/mL when using the DPPH assay, ABTS assay, and the OH scavenging test, respectively. Regarding the leaf hydrodistilled residues, results were similar in efficacy to the DPPH and ABTS test. However, when using the OH radical test, the hydrodistilled residues showed a better antioxidant activity for the species that came from Bab Saâdoun
[40][37]. The DPPH test to assess the antioxidant activity of
A. altissima leaves was also used by Rahman et al.
[39][36]. Methanolic extracts presented an IC
50 value of 35.46 μg/mL and its ethyl acetate fraction possessed an even lower IC
50 value (16.45 μg/mL). Recently, research was conducted to evaluate the physicochemical parameters and pharmacological bioactivity of
A. altissima seed oil. El Ayeb-Zakhama et al.
[52][49] were able to show that the seed oil presents antioxidant activity. The DPPH assay determined that the IC
50 was 24.57 μg/mL, although the concentration of polyphenols was considered low (1.067 mg gallic acid equivalent/100 g oil). In addition, the study also showed that this seed oil presents moderate antimicrobial activity against Gram-positive bacteria.
5.3. Anti-Inflammatory Effects
Inflammation can lead to detrimental effects of different pathologies such as neurodegenerative and cardiovascular diseases, cancer, diabetes, and others
[21][18]. It is also essential to find compounds that can tackle this problem by inhibiting the pro-inflammatory molecules and associated pathways
[53][50].
El Ayeb-Zakhama et al.
[52][49] reported that
A. altissima seed oil could induce anti-inflammatory effects on edema (in vivo experiment), by reducing it more than 60% after 3 h, via the administration of doses of 0.2 and 1 g/kg. The same study also demonstrates the analgesic capacity of the oil due to its complex composition. The acute toxicity study and the analgesic effect of the seed oil was compared with a widely marketed drug, acetylsalicylic acid (ASL), in which the average lethal dose of the oil was estimated to be more than 2 g/kg and the analgesic effect was almost as potent as ASL at a dose of 1 g/kg. Other research
[54][51] has suggested that the ethanolic extract of
A. altissima (leaves and branch) can inhibit inflammatory mediators, both in vitro and in vivo, mainly by the inhibition of cyclooxygenase-2 (COX-2). By extracting canthin-6-one, an alkaloid, from the stem barks of
A. altissima, Cho et al.
[55][52] has proven this compound’s potential effect in exerting anti-inflammatory effects of macrophages dysregulating pro-inflammatory cytokines and pathways. This same family of compounds was the target of the research conducted by Kim et al.
[56][53], amongst other compounds. The authors reached the same conclusion regarding the anti-inflammatory properties. Kang et al.
[57][54] have shown the potent inhibitory effects of
A. altissima decoction on the decrease of cytokine levels, such as tumour necrosis factor (TNF) and interleukin (IL)-6 and IL-8 as well as on the reduction of histamine levels, by using both in vitro and in vivo models.
Moreover, the nuclear factor kappaB (NF-κB) pathway can also be inhibited by
A. altissima compounds. Kim et al.
[58][55] used the ethanolic extract of
A. altissima leaves to assess the anti-inflammatory properties in astrocytes, where inflammation was induced by lipopolysaccharide. Results showed that the anti-inflammatory mechanism functions through the inhibition of COX-2 and other cytokines and the inhibition of the NF-κB pathway. These findings may lead to the use of
A. altissima as a new therapeutic approach to neuroinflammatory diseases.
Overall, studies about the anti-inflammatory effects of the
A. altissima compounds, whether they come from the leaves, bark, or seed oil, all conclusions favour the beneficial effects against pro-inflammatory molecules.
5.4. Phytotoxic Effects/Phytochemical Activity
Recent studies have demonstrated the potential of plant phytotoxins, such as ailanthone from
A. altissima, as bioherbicides.
A. altissima has produced phytotoxic compounds and the main toxin identified is the ailanthone quassinoid, first isolated by Heisey and Heisey
[30][27]. Both tested the herbicidal effect of the methanol extracts from the stem and bark of
A. altissima on 17 weed and crop species and found that their application provided a strong herbicidal effect on all tested species. Numerous studies confirmed that quassinoids have a wide range of biological activities, including antileukemic and anticancer, antiamoebal, antimalarial, insecticidal and antifeedant, antiviral, antifungal, antitubercular, and herbicidal activities
[59][56]. It is considered that the bioactivity of quassinoids is based on the plasma membrane NADH oxidase inhibition
[60][57]. The phytotoxic effect seemed to correlate better with the extracts of higher phenolic contents; such results have previously been reported in
Psychotria leiocarpa [61][58]. Phytochemical studies have shown that the main allelochemicals are ailanthone, ailantinone, chaparrin, and ailantinol with the first component being the most potent allelochemical. Moreover, both the methanolic extract and the hydrodistilled residue presented a significant phytotoxic activity
[40][37]. Lungu et al.
[51][48] extracts showed an excellent inhibitory action on the germination of seeds and growth of lettuce seedlings.
Regarding inhibiting progestin, Ahmed et al.
[62][59] analysed 13 Chinese plant species, including
A. altissima, in which they used the extract of the stem part. With the treatment of the T47D human breast cancer cell line with 314.46 ng/mL of progesterone and with three different levels of concentration of 10 μg/mL, 20 μg/mL and 40 μg/mL of ethanolic extracts, it was possible to verify that these concentrations could significantly prevent the action of the hormone in a dose-dependent manner.
Despite all the beneficial effects, some reports depict the contrary.
A. altissima has been reported to negatively affect human health, with effects such as allergies, dermatitis and myocarditis. In addition, this invasive tree can also produce toxic and noxious environments for other species in the neighbourhood from the soil since the main compound responsible for its allelopathy (ailanthone) is present in high concentrations in the root bark and when released into the soil
[6,7,63][6][7][60].
5.5. Anti-Tumour and Anti-Viral Activity
Anti-tumour activity is one of the fields that has contributed to more research on the effects that the bioactive compounds of
A. altissima exert. Several studies with the dried bark, show that its active compounds exert antitumor effects in several cell lines, for example in A549, MDA-MB-231, LAPC4, A375, B16, and SGC-7901 among others. These in vitro studies lead us to state that ailanthone specifically may be a good inhibitor for cancers such as melanoma, acute myeloid leukaemia, lung, liver, breast, bladder, osteosarcoma, and prostate cancer
[22,64,65][19][61][62]. In the work of Ding et al.
[65][62] the different mechanisms of the action of ailanthone that justify its inhibitory effects were described and explained. The main basis of these effects is the mechanism of apoptosis. Apoptosis can be triggered by regulation of Bcl-2 family proteins, transcription factors such as β-catenin, tumour suppressor genes, and by the PI3K/AKT/mTOR and JAK/STAT3 pathways. Arylanthone induces apoptosis by the downregulation of Bcl-2 and the downregulation of the Bax proteins. It also induces this cell death through the downregulation of p53 (tumour suppressor protein) and miR-195, which consequently leads to inhibition of the JAK/STAT3 signalling pathway. On the other hand, the signalling pathway which is responsible for cell proliferation, differentiation, and metabolism that can lead to anti-apoptosis is PI3K/AKT/mTOR. In this pathway ailanthone has a suppressive effect, inducing apoptosis through the phosphorylation of PI3K and AKT. The review work of these authors is very interesting in that they warn of the problem that there are few studies evaluating the cytotoxicity of ailanthone in normal cells. Importantly, a preclinical study by Tang et al.
[66][63] assessed the safety of ailanthone in Kumming rats and classified the mean lethal dose of ailanthone (27.3 mg/kg) as level 2 (severe) from the Globally Harmonised System of Classification and Labelling of Chemicals.
Furthermore, they warn that research into this active compound from
A. altissima is still very early, as there are no studies on the bioavailability and side effects of ailanthone. Researchers are advised to focus on comparing the efficacy of ailanthone with other existing chemotherapy drugs and also to conduct in vivo and clinical trials
[65][62].
Regarding the antiviral activity, some studies demonstrated that the main compound responsible for this effect is the main compound of
A. altissima: ailanthone. In the review by Li et al.
[22][19] the authors summarize all the existing pharmacology considering the dried bark of this species. They state that the active compound shows moderate inhibitory effects against the tobacco mosaic virus (TMV). In a study conducted by Tan et al.
[67][64] these effects reached IC
50 values of 0.30 mmol/L. However, they mention that the mechanism of action leading to this activity is not yet well elucidated.
5.6. Potential and New Applications
Desami et al., 2019
[68][65], and Caser et al., 2020
[69][66], provide new insights into
A. altissima extracts and their phytotoxicity as potential natural herbicide use as a sustainable solution for weed management in horticultural crops.
The methanolic extracts of
A. altissima showed antibacterial activity against different food pathogens; simultaneously, antioxidant properties indicated a potential use of extracts from this plant as food preservatives
[39][36]. However, toxicity studies must be developed concerning the uses in the food industry and given the presence of alkaloids. The papermaking potential of this species, and its use as a fiber alternative for pulp production for the paper, was also evaluated
[70,71][67][68]. The results of the paper are close to those of the reference ones. Regarding the growth and longevity of the
A. altissima tree, other wood applications could be studied for this species.