Subclinical inflammation in morbid obesity is associated with the activation of the immune system and the development of concomitant diseases. Impaired immune homeostasis and dysregulation of immune cells in adipose tissue are associated with phenotypic and functional changes in the pool of T-lymphocytes and the development of chronic vitamin D deficiency. Low vitamin D levels in obesity lead to activation, proliferation, and production of pro-inflammatory mediators T cells. Hypovitaminosis D is the reason for the decrease in the functional potential of regulatory and anti-inflammatory lymphocytes and the maintenance of the inflammatory response.Субклиническое воспаление при патологическом ожирении связано с активацией иммунной системы и развитием сопутствующих заболеваний. Нарушение иммунного гомеостаза и нарушение регуляции иммунных клеток в жировой ткани связаны с фенотипическими и функциональными изменениями пула Т-лимфоцитов и развитием хронического гиповитаминоза D. Низкий уровень витамина D при ожирении приводит к активации, пролиферации и продукции провоспалительных медиаторов. Т-клетками. Гиповитаминоз D является причиной снижения функционального потенциала регуляторных и противовоспалительных лимфоцитов и поддержания воспалительного ответа.
- vitamin D
- autoimmune disease
- Th17 cells
- inflammation
- obesity
1. IntroductionВведение
2. Vitamin D Metabolism
Vitamin D is a fat-soluble secosteroid, 70–80% of which is synthesized in a photolysis reaction in the skin [18][20]. Under the influence of ultraviolet radiation (UV) from the B spectrum of the sun, the process of photosynthesis in the epidermis is activated. In this process, 7-dehydrocholesterol (provitamin D) is converted into previtamin D3, which undergoes isomerization of three double bonds after a thermochemical reaction, forming the inactive vitamin D3 (cholecalciferol). The synthesis of vitamin D3 takes up to 3 days from the time the skin is exposed to UV rays. This process is the main source of vitamin D and it depends on the intensity of UV radiation (latitude, season, time of day), skin pigmentation and air pollution [18][19][20][20,21,22] (Figure 1).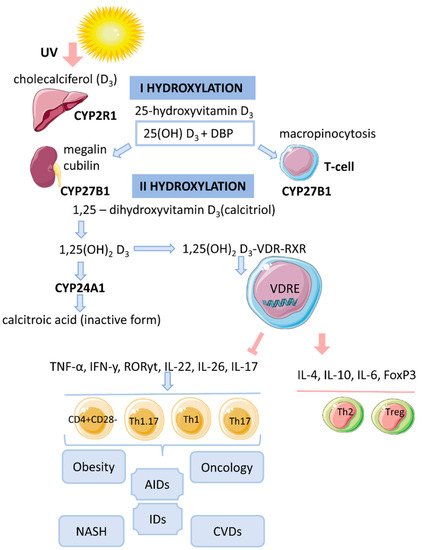
3. Cytosolic Complex 1,25(OH)2D3-VDR-RXRF
The biological effect of 1,25(OH)2D3 is related to the activation of the cytosolic VDR receptor, whose active ligand is DBP and an indirect change in the transcriptional activity of vitamin D-associated genes [14][17]. VDR is a transcription factor and belongs to the family of nuclear receptors for steroid hormones (receptors for retinoic acid, thyroid hormones, sex hormones, adrenal hormones). The human VDR gene consists of eight coding exons (two non-coding) and two promoters and is located on chromosome 12 [19][21]. The VDR protein contains 427 amino acid residues. It acts as an obligate heterodimer that interacts with the retinoid X receptor (RXR), which subsequently causes the translocation of the complex into the nucleus and binds in a ligand-dependent manner to the promoter regions of target genes that are sensitive to vitamin D [14][18][17,20]. The functional domains of VDRs are the highly conserved NH2-terminal DNA-binding domain (DBD) and the more variable COOH-terminal ligand-binding domain (LBD). The DBD is a region with two zinc fingers, each containing a zinc atom in a tetrahedral arrangement with four invariant cysteine residues [19][21][22][21,30,31]. The lipophilic molecule 1,25(OH)2D3 can pass through the cell membrane and interact with the VDR [18][20]. Binding of 1,25(OH)2D3 to the VDR leads to conformational changes in the structure of the VDR protein that facilitate interaction with RXR and coregulatory complexes involved in the transcription of target genes [19][21]. Subsequently, the cytosolic complex 1,25(OH)2D3-VDR-RXR migrates to the nucleus, where it interacts with VDRE [15][14] (Figure 1). In the nucleus, the 1,25(OH)2D3-VDR-RXR complex interacts with histone acetyltransferases, transcriptional coactivators, corepressors and chromatin-restructuring complexes to modulate the transcription of target genes [16][18].4. Genomic Mechanism of Action of Vitamin D (T-lymphocytes)
After the identification of VDR in activated T lymphocytes, vitamin D has been proposed as a regulator of the immune system [13]. The active form of vitamin D, 25(OH)2D3, has an immunomodulatory effect on many components of the innate and adaptive immune systems [14][17]. 1,25(OH)2D3 regulates differentiation and maturation of subpopulations of innate immunity cells, antigen presentation and the production cytokines and chemokines. 1,25(OH)2D3 inhibits the inflammatory response by suppressing the expression of Toll-like receptors 2 and 4 (TLR2/4) and the secretion of pro-inflammatory mediators (IL-1, IL-6, TNF-α). In addition, the active form of vitamin D negatively regulates the differentiation, maturation and immunomodulatory capacity of dendritic cells (DCs) by reducing the expression of MHCII, CD40, CD80, CD86 and the maturation proteins CD1a, CD83 [14][17]. 1,25(OH)2D3 also inhibits the DC-mediated activation process of T cells and decreases the expression of inflammatory mediators (IL-12, IFNy in DCs) [5][18][5,20]. 1,25(OH)2D3 inhibits the proliferation of T lymphocytes and the production of IFNy and IL-17 and increases the secretion of IL-4 and IL-10. Thus, 1,25(OH)2D3 enhances the regulatory Th2 immune response and induces the differentiation of Treg cells, thereby reducing the pro-inflammatory potential of Th1 and Th17 cells. The complex 1,25(OH)2D3-VDR-RXR blocks the formation of NFAT/AP-1. It is known that the formation of the NFAT/AP-1 complex is necessary for the activation of the IL-2 promoter. The repressive effect of 1,25(OH)2D3 on IFNy gene transcription is due to the direct interaction of VDR-RXR with the silencer regions on the gene promoter. 1,25(OH)2D3 enhances the production of IL-4 by Th2 cells and potentiates the regulatory properties of Treg cells by activating the expression of the transcription factor FoxP3 [5][20][5,22]. The immunomodulatory effect of 1,25(OH)2D3 is also associated with an increase in IL-6 secretion, which may lead to a shift in the balance towards the Treg cell response [18][23][20,32]. 1,25(OH)2D3 prevents IL-17 production by suppressing inflammation and Th17-mediated autoimmunity [24][25][33,34]. The mechanism of IL-17 suppression by 1,25(OH)2D3 is based on blocking NFAT binding to the IL-17 gene promoter, sequestration of Runx1 factor by the VDR, recruitment of histone deacetylase (HDAC) and induction of FoxP3 expression [20][22]. In addition, the VDR interacts with the P105/P50, P100/P52 and P65 proteins of the NF-kB factor.5. Vitamin D and T-lymphocytes
5.1. Th1 and Th2 Cells
5.2. Non-Pathogenic Th17 Cells and Treg
5.3. Th1.17-Cells
5.4. Memory T Cells
6. Hypovitaminosis D in Obesity
Based on previously obtained data, theories have been formulated that partially explain the relationship between hypovitaminosis D and obesity [44][45][46][55,56,57].- (1)
-
Volume dilution. Serum vitamin D levels decrease with increasing body size [47][48][58,59].
- (2)
-
Sequestration of vitamin D in adipose tissue. Vitamin D (synthesized by skin, supplements/medications) becomes tightly bound in fat stores and does not enter the bloodstream in sufficient quantity to maintain serum levels of 25(OH)D3 [44][46][55,57].
- (3)
-
Different ability to activate vitamin D in adipose tissue of lean and obese individuals [47][58]. In adipocytes, high expression of the enzymes 1α-hydroxylase (mitochondrial CYP27B1) and 25-hydroxylase CYP2J2 was found [44][55]. However, their expression is lower in obese people compared to lean people [49][60].
7. Molecular Mechanism of Action of Vitamin D on T Cells under Hypoxic Conditions Associated with Obesity
In obesity, the formation of local foci of hypoxia is observed. At the same time, hypoxia activates the transcription of the factor HIF-1α, which, together with mTORC1, promotes pTh17 cell differentiation [32][41]. Vitamin D has been shown to suppress LPS-induced expression of HIF-1α, thereby reducing the extent of hypoxia [50][62]. The mTORC1 pathway is believed to play an important role in the induction of plastic properties and the formation of Th1.17 cells (with a pathogenic phenotype) [51][63]. The PI3K (phosphatidylinositol 3-kinase)-AKT-mTORC1-S6K axis has been described as a positive regulator of Th17 cell differentiation that induces nuclear translocation of the RORyt factor [52][64]. InКроме addition, hypoxia and other cellular disturbances (DNA damage, endoplasmic reticulum stress, energy stress) cause transcription of the того, гипоксия и другие клеточные нарушения (повреждение ДНК, стресс эндоплазматического ретикулума, энергетический стресс) вызывают транскрипцию гена DDIT4 gene (DNA damage-induced transcript 4, or(транскрипт 4, индуцируемый повреждением ДНК, или REDD1). RecentНедавние studies have shown that binding of исследования показали, что связывание 1,25 (OH)2 2 D3 3 toс VDR can increaseможет увеличивать экспрессию гена DDIT4 gene expression. ConsideringУчитывая, that activation of что активация DDIT4 leads to inhibition of приводит к ингибированию mTORC1, their interaction can be regulated by их взаимодействие может регулироваться 1,25 (OH) 2 D 3 [ 65 [53]]. AtВ the same time,то же время DDIT4 suppresses the activity of подавляет активность mTORC1, inducing theиндуцируя комплекс TSC1 / 2 complex, and regulates the production of , и регулирует продукцию IL-17 in patients with multipleу пациентов с рассеянным склерозом [ sclerosis72 [54]]. UnderВ pathological conditions, adenosine triphosphate (ATP) is released from intracellular stores into the extracellular space, where it acts as a stress signal (alarm, warningпатологических условиях аденозинтрифосфат (АТФ) высвобождается из внутриклеточных запасов во внеклеточное пространство, где он действует как сигнал стресса (тревога, предупреждение, DAMP) by binding to purinergic receptors, связываясь с пуринергическими рецепторами. TheПуринергический purinergic receptor рецептор P2X7 (P2X7R), an extracellular ATP-dependent channel, is involved in the secretion of proinflammatory cytokines (which trigger theвнеклеточный АТФ-зависимый канал, участвует в секреции провоспалительных цитокинов (которые запускают воспалительный ответ), гибели клеток и аутофагии [ inflammatory38 response), cell73 death, and74 autophagy [29][55][56]].