Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 1 by Peng George Wang.
Dickkopf-related protein 1 (DKK1) is distinctly identified as an inhibitor of canonical Wnt/β-catenin signaling.
- miRNA
- AGA
- DKK1
- Wnt/β-catenin
- hair
1. Introduction
Hair growth is a relatively complex process. The human hair cycle comprises three main phases: anagen, catagen, and telogen [1]. Anagen is an active growing phase of hair and lasts for the longest period (up to eight years). During the catagen phase (2–3 weeks), hair follicles (HF) undergo regression. This is followed by the telogen, which is the resting phase (~3 months) [2]. Ultimately, hair shedding takes place, that may be considered as the fourth phase—exogen [3,4][3][4].
Hair growth is controlled and influenced by various endogenous factors, including intracellular and intercellular signaling molecules. Some of the influencing factors have inhibitory and some stimulatory effects on the hair cycle. The initiation of the anagen phase is inhibited by the activation of bone morphogenetic proteins (BMP) as well as transforming growth factor β (TGF-β) signaling [2]. On the other hand, the onset of Wnt/β-catenin [5,6,7][5][6][7] or sonic hedgehog (SHH) signaling pathways stimulate hair growth and anagen entry [2,8][2][8]. In case of the wingless and integrated-1 (Wnt)-mediated hair regrowth, hypo-phosphorylated β-catenin is stabilized, which triggers the interaction between β-catenin and T-cell factor/lymphoid enhancer factor (TCF/LEF) in the nucleus. As a result, growth-promoting genes are trans-activated and hair regrowth occurs [2]. Based on the available data, out of all the pathways implicated in the hair cycle, we assume that the Wnt/β-catenin signaling represents the key factor in hair growth regulation [9], as dihydrotestosterone (DHT)—a hormone that is upregulated in androgenetic alopecia (AGA) and causes the hair loss—impairs this signaling pathway [10]. The main identified inhibitors of this signaling pathway are dickkopf-related protein 1 (DKK1), secreted frizzled-related protein 2 (SFRP2), and sclerostin (SOST). As Wnt/β-catenin signaling is involved in numerous biological processes, each of the mentioned inhibitors features certain functions. Hence, the dysregulation of the levels of any of these inhibiting proteins affects the particular physiological pathogenesis, including AGA. DKK1 is a natural inhibitor of Wnt, and strongly suppresses the Wnt/β-catenin signaling pathway via disrupting the Wnt-induced frizzled-low-density lipoprotein receptor-related proteins (LRP) 5/6 complex formation. This adversely impacts HF morphogenesis and, thus, influences the hair cycle [11]. Indeed, the study has demonstrated that DKK1 triggers anagen-to-catagen transition when injected in the skin of C57BL/6 mice. Moreover, injection of the anti-DKK1 neutralizing antibody resulted in the delay of catagen progression [12]. Interestingly, the human study showed that a significantly high concentration of tissue DKK1 was present in patients with AGA compared to the healthy controls [11]. DHT is known to negatively interfere with the normal hair cycle via driving the HF in the balding scalp to catagen entry and inhibiting the growth of keratinocytes that takes place through DKK1 implication [13].
According to the aforementioned evidence, it is prudent to postulate that inhibition of DKK1—a canonical Wnt/β-catenin signaling inhibitor—is one of the key factors to promote hair growth and develop a sustainable remedy for AGA. Indeed, there are certain microRNAs (miRNAs) that target DKK1 and are involved in hair growth-related pathways.
2. DKK1 Implication in AGA
2.1. DHT-Induced DKK1-Mediated AGA
DKK genes encode secreted proteins that regulate the Wnt/β-catenin signaling pathway by antagonizing it [43][14]. Remarkably, DHT and DKK1 levels are positively correlated. In the body, 5 alpha-reductase (5αR) converts testosterone into DHT. It acts like an inhibitor for the growth of outer root sheath (ORS) cells that disrupts the normal hair growth process. Moreover, the anti-DKK1 neutralizing antibody substantially decreased the inhibition of ORS cells’ growth. As previously mentioned, DKK1 concentration is increased in the bald scalp of AGA patients compared with the haired scalp of AGA patients [10,40][10][15]. Besides, DHT-influenced DKK1 augmentation has been demonstrated to enhance the apoptosis of keratinocytes in vitro [10]. It can be presumed that DKK1 plays an important role in AGA development. Indeed, the study demonstrated that DKK1 is involved in AGA pathology. In the study, recombinant human DKK1 treatment triggered the hair cycle to enter the catagen phase earlier than normal in C57BL/6 mice, which resulted in the decrease of HF length. Contrarily, treatment with the neutralizing DKK1 antibody led to the increased HF length and delayed the shift of anagen to catagen. The recombinant DKK1 has inhibited the canonical Wnt/β-catenin signaling pathway that promotes hair growth in normal physiological condition. As a result of Wnt/β-catenin signaling suppression, the anagen phase is shortened and the apoptosis of follicular keratinocytes takes place [12].
2.2. Negative Influence of DKK1 on Hair Growth
DKK1 exhibits a dual role in the normal hair cycle. On the one hand, DKK1 induces attenuation of the hair growth process by inhibiting Wnt/β-catenin signaling via the LRP5/6 co-receptor and, on the other hand, it promotes apoptosis of keratinocytes, which are key cells involved in hair growth [10,40][10][15]. Hence, AGA, also known as male pattern baldness (MPB), is the consequence of the abovementioned dual mechanism of DKK1 in human HFs. Indeed, a case-control study that included 20 male AGA and 20 male alopecia areata (AA) patients has demonstrated that the immunohistochemical expression of DKK1 was remarkably increased in lesional scalp biopsies of both AGA and AA patients [40][15]. Additionally, DKK1 decreases the HF enlargement and width of the hairs [31][16]. Markedly, DKK1 levels are evidenced to be elevated along with age [44[17][18],45], whilst age is also related to the development of AGA [46][19] (Figure 2). Besides, AGA is manifested severely in the obese population [47,48][20][21]. Concomitantly, DKK1 is hypothesized as a potential biomarker in obesity [49,50][22][23]. Additionally, Kim et al. have demonstrated that treatment with minoxidil—a common drug for hair-loss—downregulated DKK1 and TGF-β in human keratinocyte cells [51][24].
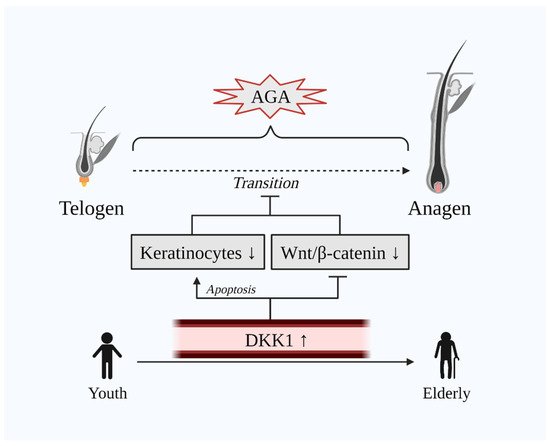
Figure 2. Dual unfavorable role of DKK1 on hair growth. The transition of telogen to anagen is delayed in AGA. DKK1 is upregulated in the scalp of AGA as well as in the serum of the elderly population where AGA is common. Elevated DKK1 levels induce the apoptosis of keratinocytes and inhibit Wnt/β-catenin signaling. Abbreviations: AGA, androgenetic alopecia; DKK1, dickkopf-related protein 1; Wnt, wingless and integrated-1.
2.3. Molecular Mechanism of Wnt/β-Catenin Signaling Inhibition by DKK1
DKK1 inhibits Wnt/β-catenin signaling via binding to LRP5 and LRP6, which prevents the interaction between Wnt and the other transmembrane receptor frizzled [52][25]. The inhibition is enhanced via the synergistic effect of DKK1 and its single transmembrane receptors kremens (KRM1 and KRM2) that promote the endocytosis of LRPs. Cselenyi and Lee have proposed that KRMs-dependent activation/inhibition of Wnt/β-catenin signaling depends on the presence of DKK1 [20,53][26][27]. In case of DKK1 presence, LRP5/6, DKK1, and KRM interact with each other and generate a complex that is endocytosed. Consequently, LRP5/6 is reduced in the plasma membrane; thus, Wnt/β-catenin signaling is suppressed, β-catenin is degraded, and the hair growth-related gene expression does not take place [54][28]. The inhibitive role of DHT-induced secreted DKK1 on the Wnt/β-catenin signaling pathway is shown in Figure 3.
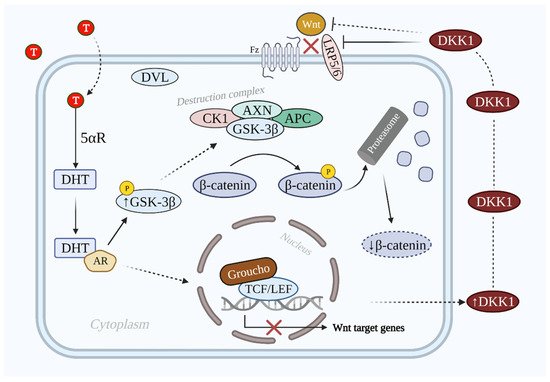
Figure 3. Illustration of Wnt/β-catenin signaling pathway inhibition by DKK1 in AGA. In the AGA population the T level is elevated. T is converted into DHT by 5αR, which binds to AR and prevents the dephosphorylation of GSK-3β. GSK-3β induces the phosphorylation of β-catenin via the destruction complex. Phosphorylated β-catenin is degraded by the proteasome. On the other hand, DKK1 is also secreted that antagonizes Wnt/β-catenin signaling via LRP5/6 inhibition. As a result, the Wnt/β-catenin signaling pathway is inactivated and target genes are not expressed. Abbreviations: T, testosterone; DHT, dihydrotestosterone; 5αR, 5 alpha-reductase; AR, androgen receptor; DKK1, dickkopf-related protein 1; Wnt, wingless and integrated-1; Fz, frizzled; LRP5/6, low-density lipoprotein receptor-related proteins 5/6; GSK-3β, glycogen synthase kinase 3β; CK1, casein kinase 1; DVL, dishevelled; APC, adenomatous polyposis coli; TCF/LEF, T-cell factor/lymphoid enhancer factor.
2.4. Compounds Capable to Inhibit DKK1 Expression and Promote Hair Growth
According to the abovementioned information, the study of DKK1 inhibition strategies is an important tool for maintaining the Wnt/β-catenin signaling within the normal range. Indeed, some studies investigated the effect of certain natural compounds on the canonical Wnt/β-catenin signaling pathway for promoting hair growth. Zhou et al. have observed the impact of morroniside—a natural compound found in cornelian cherry—on the Wnt/β-catenin signaling pathway in cultured human ORS cells. In their results, morroniside indeed increased the proliferation of cells as well as the concentration of β-catenin. Concomitantly, the outcome of morroniside treatment was retrieved by the influence of DKK1 [52][25]. Another natural compound, vitexin, is also found to significantly increase the human dermal papilla cells’ (HDPCs) proliferation in a concentration-dependent fashion. Additionally, Luo et al. have demonstrated that in vitexin-treated HDPCs, the β-catenin level was upregulated while the DKK1 level was substantially lowered [55][29]. Aside from that, Panax ginseng extract exhibited proliferation of ORS keratinocytes, inhibition of apoptosis, and revealed the opposite effect of DKK1 in human hair organ culture [56][30]. One more natural compound costunolide activates Wnt/β-catenin and SHH, while it inhibits TGF-β/SMAD and BMP signaling pathways in HFs, and hence, stimulates hair growth [57][31]. The functional components of Ginkgo biloba—ginkgolide B and bilobalide—are known as agents that can aid in hair growth. Interestingly, these two compounds are evidenced to be associated with Wnt/β-catenin signaling activation via inhibition of DKK1 expression levels in the cytoplasm and, thus, upregulation of nuclear β-catenin and its messenger RNA (mRNA) concentration [58][32]. The antidepressant tianeptine is also evidenced to stimulate hair shaft elongation, and hence hair growth, via inhibiting DKK1 and delaying the premature transition from anagen to catagen in MPB [59][33]. All these studies claim that the abovementioned natural compounds promote hair growth via inhibiting DKK1.
3. miRNAs Targeting DKK1 Expression
miRNA was first identified in 1993 by Victor Ambros and colleagues, who found a short RNA molecule Lin-4 of 22–61 nucleotides in C. elegans that could downregulate the level of Lin-14 protein mRNA [60][34]. Seven years after the discovery, the first human-encoded miRNA Let-7 was described [61][35]. Currently, more than 3000 miRNAs are known to be present in the human genome and involved in numerous biological processes in almost all body fluids [62,63][36][37]. In 2013, the first miRNA mimic MRX34 (mimic of endogenous miR-34 that suppresses oncogenesis) entered into the clinical trials [64][38]. Currently, several miRNA drugs are in phase 1 or 2 of development [65,66][39][40]. However, up until now, there is no miRNA mimic approved by the Food and Drug Administration (FDA) [66][40]. On the other hand, there are already 3 small (short)-interfering RNA (siRNA)-based FDA-approved drugs, Patisiran [67][41], Givosiran [68][42], and Lumasiran [69][43], while the rest are in phase 3, 2, or 1 of clinical trials [66][40]. Given that it took 14 years since the initiation of the very first clinical trial (2004) [70][44] for siRNA to go to commercialization [71[45][46],72], it should not be long until the first miRNA mimic gains FDA approval. However, difficulties mainly regarding the delivery of miRNAs need to be overcome and the mechanisms of certain miRNAs should be fully understood.
Importantly, miRNAs play a crucial role in hair growth regulation [73][47]. They are involved in HF development as well as in DP cell proliferation. However, despite the increasing number of studies that demonstrate a critical role of miRNAs in skin regeneration, the molecular mechanisms are still not fully understood [74][48]. Some miRNAs that are implicated in hair morphogenesis have a positive while some have a negative impact on hair growth, e.g., one study showed that miR-214 targets β-catenin and modulates Wnt/β-catenin signaling, and hence inhibits hair growth development [75][49]. On the other hand, other miRNAs, such as miR-218-5p, are related to inhibition of SFRP2 that represents the antagonist of the Wnt/β-catenin signaling pathway and, thus, promotes hair growth [76,77][50][51]. As stated above, the level of DKK1 in the scalp of AGA patients is significantly higher [40][15]. Besides, DKK1 is already evidenced to inhibit the Wnt/β-catenin signaling, which results in hair miniaturization and growth suppression [78][52]. There are miRNAs that target DKK1, e.g., miR-335-5p is downregulated while DKK1 protein levels are increased in TNF-α-treated osteoblasts [79][53]. DKK1 is silenced by miRNAs including miR-335-5p at an early stage of osteogenic differentiation, and on the contrary, miRNA levels are decreased at a later stage of differentiation while DKK1 levels are upregulated. This points out the complex role of miRNAs in biological processes [80][54]. Besides, Michel et al. have demonstrated that the expression of Wnt antagonist SFRP2 mRNA was increased while the expression of another inhibitor DKK1 mRNA was not altered in scalp biopsies of AGA patients [81][55]. This fact might be rationalized with the involvement of certain miRNAs that target DKK1 mRNA, and the translation to protein is circumvented. Accordingly, developing the miRNA-based approach to inhibit DKK1 is certainly reasonable. The miRNAs that inhibit DKK1 expression in different health conditions/biological processes are presented in Table 1.
Table 1. Recent studies referring to miRNAs that directly target DKK1 and the association with certain disorders/biological processes.
| miRNA | Disorder/Biological Process | miR Level | DKK1 Relative Expression | miR Predicted Binding Site in 3′UTR of DKK1 | Study Type | Ref. | |
|---|---|---|---|---|---|---|---|
| miR-1-3p | Oral squamous cell carcinoma | ↓ | ↑ |  |
in vitro | [82] | [56] |
| miR-9-5p | Dopaminergic neuron differentiation | N/A | ↓ |  |
in vitro | [83] | [57] |
| miR-29a | Bone metabolism disorder | ↓ | ↑ |  |
in vitro | [84] | [58] |
| miR-31 | Breast cancer | ↓ | ↑ | 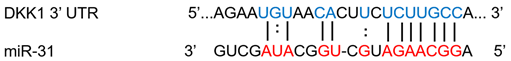 |
in vitro | [85] | [59] |
| miR-33a-5p | Esophageal cancer | ↓ | ↑ |  |
in vivo in vitro |
[86] | [60] |
| miR-34a | Cardiac hypertrophy | ↑ | ↓ |  |
in vivo | [87] | [61] |
| miR-101-3p | Osteogenic differentiation | ↑ | ↓ |  |
in vitro | [88] | [62] |
| miR-103a-3p | Osteogenic differentiation | ↑ | ↓ |  |
in vitro | [89] | [63] |
| miR-107 | Osteosarcoma | ↓ | ↑ |  |
in vitro | [90] | [64] |
| miR-130b-3p | Melanoma | ↑ | ↓ | 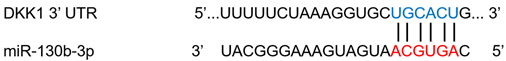 |
in vitro | [91] | [65] |
| miR-146a | Ankylosing spondylitis | ↑ | ↓ |  |
ex vivo | [92] | [66] |
| miR-152 | Multiple myeloma | ↓ | ↑ |  |
in vitro in vivo |
[93] | [67] |
| *miR-186-5p | Idiopathic pulmonary fibrosis | ↓ | ↑ |  |
in vitro, ex vivo, in vivo |
[94] | [68] |
| miR-203 | Lung adenocarcinoma | ↓ | ↑ |  |
ex vivo, in vitro |
[95] | [69] |
| Osteoporosis | ↑ |  |
in vitro | [96] | [70] | ||
| miR-217 | Colon cancer | ↑ | ↓ | 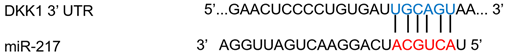 |
in vitro | [97] | [71] |
| Hepatocellular carcinoma | ↑ | ↓ |  |
in vitro/ex vivo in vivo |
[98] | [72] | |
 |
|||||||
| Osteonecrosis | ↓ | ↑ |  |
in vitro, ex vivo |
[99] | [73] | |
| miR-291a-3p | Osteoporosis | ↓ | ↑ |  |
in vitro, in vivo |
[100] | [74] |
| miR-302b-3p | Multiple myeloma | ↓ | ↑ |  |
in vitro, in vivo |
[101] | [75] |
| miR-302e | Cardiac hypertrophy | ↑ | N/A |  |
in vitro | [102] | [76] |
| Cervical cancer | ↓ | ↑ |  |
in vitro, in vivo |
[103] | [77] | |
| miR-335-5p | Diabetic osteoporosis | ↓ | ↑ |  |
in vitro | [104] | [78] |
| miR-371/372/373 | Stem cell tumorigenesis | ↑ | ↓ |  |
in vitro | [105] | [79] |
| miR-373-3p | Tongue squamous cell carcinoma | ↑ | ↓ |  |
in vitro | [106] | [80] |
| miR-410 | Colorectal cancer | ↑ | ↓ |  |
in vitro | [107] | [81] |
| miR-433-3p | Osteogenic differentiation | ↑ | ↓ | 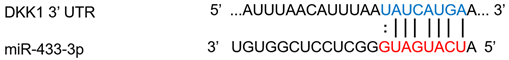 |
in vitro | [108] | [82] |
| miR-488 | Fracture | ↓ | ↑ |  |
in vitro | [109] | [83] |
| miR-493-3p | Gastric cancer | ↑ | ↓ |  |
in vitro | [110] | [84] |
| miR-522 | Hepatocellular carcinoma | ↑ | ↓ |  |
in vitro | [111] | [85] |
| miR-523-3p | Retinoblastoma | ↑ | ↓ |  |
in vitro, ex vivo |
[112] | [86] |
| miR-613 | Rheumatoid arthritis | ↓ | ↑ | 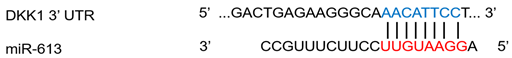 |
in vitro | [113] | [87] |
| miR-3064-3p | Cementoblast differentiation | ↓ | ↑ |  |
in vitro | [114] | [88] |
| miR-6783-3p | Lung adenocarcinoma | ↑ | ↓ |  |
in vitro, in vivo |
[115] | [89] |
| miR-6807-3p | Lung adenocarcinoma | ↑ | ↓ |  |
in vitro, in vivo |
[116] | [90] |
| miR-BART10-3p | Gastric carcinoma | ↑ | ↓ |  |
in vitro | [117] | [91] |
| miR-BART22 | Gastric carcinoma | ↑ | ↓ |  |
in vitro | [118] | [92] |
Notes: Dots between miRNA and DKK1 sequence indicate wobble pairing. N/A, not applicable. *, These binding sites are cited from TargetScan.org.
Notes: Dots between miRNA and DKK1 sequence indicate wobble pairing. N/A, not applicable. *, These binding sites are cited from TargetScan.org.
References
- Chen, C.L.; Huang, W.Y.; Wang, E.H.C.; Tai, K.Y.; Lin, S.J. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed. Sci. 2020, 27, 1–11.
- Choi, B.Y. Targeting wnt/β-catenin pathway for developing therapies for hair loss. Int. J. Mol. Sci. 2020, 21, 4915.
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal E ff ects on Hair Follicles. Int. J. Mol. Sci. 2020, 21, 5342.
- Houschyar, K.S.; Borrelli, M.R.; Tapking, C.; Popp, D.; Puladi, B.; Ooms, M.; Chelliah, M.P.; Rein, S.; Pförringer, D.; Thor, D.; et al. Molecular mechanisms of hair growth and regeneration: Current understanding and novel paradigms. Dermatology 2020, 236, 271–280.
- Li, Y.H.; Zhang, K.; Ye, J.X.; Lian, X.H.; Yang, T. Wnt10b promotes growth of hair follicles via a canonical Wnt signalling pathway. Clin. Exp. Dermatol. 2011, 36, 534–540.
- Li, Y.H.; Zhang, K.; Yang, K.; Ye, J.X.; Xing, Y.Z.; Guo, H.Y.; Deng, F.; Lian, X.H.; Yang, T. Adenovirus-mediated wnt10b overexpression induces hair follicle regeneration. J. Investig. Dermatol. 2013, 133, 42–48.
- Xiong, Y.; Liu, Y.; Song, Z.; Hao, F.; Yang, X. Identification of Wnt/β-catenin signaling pathway in dermal papilla cells of human scalp hair follicles: TCF4 regulates the proliferation and secretory activity of dermal papilla cell. J. Dermatol. 2014, 41, 84–91.
- Lin, W.; Xiang, L.; Shi, H.; Zhang, J.; Jiang, L.; Cai, P.; Lin, Z.; Lin, B.; Huang, Y.; Zhang, H.; et al. Fibroblast Growth Factors Stimulate Hair Growth through β-Catenin and Shh Expression in C57BL/6 Mice. Biomed Res. Int. 2015, 2015, 730139.
- Gentile, P.; Garcovich, S. Advances in Regenerative Stem Cell Therapy in Androgenic Alopecia and Hair Loss: Wnt Pathway, Growth-Factor, and Mesenchymal Stem Cell Signaling Impact Analysis on Cell Growth and Hair Follicle Development. Cells 2019, 8, 466.
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Investig. Dermatol. 2008, 128, 262–269.
- Fawzi, M.M.T.; Mahmoud, S.B.; Shaker, O.G.; Saleh, M.A. Assessment of tissue levels of dickkopf-1 in androgenetic alopecia and alopecia areata. J. Cosmet. Dermatol. 2016, 15, 10–15.
- Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dickkopf 1 promotes regression of hair follicles. J. Investig. Dermatol. 2012, 132, 1554–1560.
- Premanand, A.; Rajkumari, B.R. Androgen modulation of Wnt/β-catenin signaling in androgenetic alopecia. Arch. Dermatol. Res. 2018, 310, 391–399.
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481.
- Mahmoud, E.A.; Elgarhy, L.H.; Hasby, E.A.; Mohammad, L. Dickkopf-1 Expression in Androgenetic Alopecia and Alopecia Areata in Male Patients. Am. J. Dermatopathol. 2019, 41, 122–127.
- Lei, M.; Guo, H.; Qiu, W.; Lai, X.; Yang, T.; Randall, B.; Chuong, C.; Lian, X.; Yang, L. Modulating hair follicle size with Wnt10b-DKK1 pair during hair regeneration. Exp. Dermatol. 2015, 23, 407–413.
- Seib, D.R.M.; Corsini, N.S.; Ellwanger, K.; Plaas, C.; Mateos, A.; Pitzer, C.; Niehrs, C.; Celikel, T.; Martin-Villalba, A. Loss of dickkopf-1 restores neurogenesis in old age and counteracts cognitive decline. Cell Stem Cell 2013, 12, 204–214.
- Dovjak, P.; Dorfer, S.; Föger-Samwald, U.; Kudlacek, S.; Marculescu, R.; Pietschmann, P. Serum levels of sclerostin and dickkopf-1: Effects of age, gender and fracture status. Gerontology 2014, 60, 493–501.
- Ho, C.H.; Sood, T.; Zito, P.M. Androgenetic Alopecia; StatPearls Publishing: Treasure Island, FL, USA, 2021.
- Yang, C.C.; Hsieh, F.N.; Lin, L.Y.; Hsu, C.K.; Sheu, H.M.; Chen, W. Higher body mass index is associated with greater severity of alopecia in men with male-pattern androgenetic alopecia in Taiwan: A cross-sectional study. J. Am. Acad. Dermatol. 2014, 70, 297–302.e1.
- Morinaga, H.; Mohri, Y.; Grachtchouk, M.; Asakawa, K.; Matsumura, H.; Oshima, M.; Takayama, N.; Kato, T.; Nishimori, Y.; Sorimachi, Y.; et al. Obesity accelerates hair thinning by stem cell-centric converging mechanisms. Nature 2021, 595, 266–271.
- Ali, H.; Zmuda, J.M.; Cvejkus, R.K.; Kershaw, E.E.; Kuipers, A.L.; Oczypok, E.A.; Wheeler, V.; Bunker, C.H.; Miljkovic, I. Wnt pathway inhibitor DKK1: A potential novel biomarker for adiposity. J. Endocr. Soc. 2019, 3, 488–495.
- Colditz, J.; Picke, A.; Hofbauer, L.C.; Rauner, M. Contributions of Dickkopf-1 to Obesity-Induced Bone. J. Bone Miner. Res. 2020, 4, 1–14.
- Kim, M.J.; Lim, C.; Lee, J.Y.; Im, K.R.; Yoon, K.S.; Song, J.M. Visible-to-near IR quantum dot-based hypermulticolor high-content screening of herbal medicines for the efficacy monitoring of hair growth promotion and hair loss inhibition. J. Biomol. Screen. 2013, 18, 462–473.
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Morroniside regulates hair growth and cycle transition via activation of the Wnt/β -catenin signaling pathway. Sci. Rep. 2018, 8, 13785.
- Choi, Y.S.; Zhang, Y.; Xu, M.; Yang, Y.; Ito, M.; Peng, T.; Cui, Z.; Nagy, A.; Hadjantonakis, A.K.; Lang, R.A.; et al. Distinct functions for Wnt/β-Catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell 2013, 13, 720–733.
- Cselenyi, C.S.; Lee, E. Context-dependent activation or Inhibition of Wnt-beta-catenin signaling by kremen. Sci. Signal. 2008, 1, pe10.
- Rothbächer, U.; Lemaire, P. Crème de la Kremen of Wnt signalling inhibition. Nat. Cell Biol. 2002, 4, E172–E173.
- Luo, J.; Chen, M.; Liu, Y.; Xie, H.; Yuan, J.; Zhou, Y.; Ding, J.; Deng, Z.; Li, J. Nature-derived lignan compound VB-1 exerts hair growth-promoting effects by augmenting Wnt/β-catenin signaling in human dermal papilla cells. PeerJ 2018, 6, e4737.
- Lee, Y.; Kim, S.U.N.A.; Hong, Y.D.; Park, B.C.; Na, Y. Panax ginseng extract antagonizes the effect of DKK-1-induced catagen-like changes of hair follicles. Int. J. Mol. Med. 2017, 40, 1194–1200.
- Kim, Y.E.; Choi, H.C.; Nam, G.; Choi, B.Y. Costunolide promotes the proliferation of human hair follicle dermal papilla cells and induces hair growth in C57BL/6 mice. J. Cosmet. Dermatol. 2019, 18, 414–421.
- Zhang, H.; Shi, Q.; Nan, W.; Wang, Y.; Wang, S.; Yang, F.; Li, G. Ginkgolide B and bilobalide promote the growth and increase β-catenin expression in hair follicle dermal papilla cells of American minks. BioFactors 2019, 45, 950–958.
- Jeong, H.S.; Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Attenuation of Dickkopf 1-Induced Hair Growth Inhibition in Cultured Human Hair Follicles by Tianeptine. Ann. Dermatol. 2017, 29, 102–105.
- Feinbaum, R.; Ambros, V.; Lee, R. The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to lin-14. Cell 1993, 75, 843–854.
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Mu, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89.
- Gazerani, P. Current Evidence on Potential Uses of MicroRNA Biomarkers for Migraine: From Diagnosis to Treatment. Mol. Diagn. Ther. 2019, 23, 681–694.
- Ahmed, S.P.; Castresana, J.S.; Shahi, M.H. Glioblastoma and MiRNAs. Cancers 2021, 13, 1581.
- Bouchie, A. First microRNA mimic enters clinic. Nat. Biotechnol. 2013, 31, 577.
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138.
- Zhang, S.; Cheng, Z.; Wang, Y.; Han, T. The Risks of miRNA Therapeutics: In a Drug Target Perspective. Drug Des. Dev. Ther. 2021, 15, 721–733.
- Mullard, A. 2018 FDA drug approvals. Nat. Rev. Drug Discov. 2019, 18, 85–89.
- Mullard, A. 2019 FDA drug approvals. Nat. Rev. Drug Discov. 2020, 19, 79–84.
- Zhang, M.M.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X.B. The growth of siRNA-based therapeutics: Updated clinical studies. Biochem. Pharmacol. 2021, 189, 114432.
- Ozcan, G.; Ozpolat, B.; Coleman, R.L.; Sood, A.K.; Lopez-Berestein, G. Preclinical and clinical development of siRNA-based therapeutics. Adv. Drug Deliv. Rev. 2015, 87, 108–119.
- Hoy, S.M. Patisiran: First Global Approval. Drugs 2018, 78, 1625–1631.
- Ledford, H. Gene-silencing drug approved Outrage over changes to EPA chemical assessments. Nature 2018, 560, 291–292.
- Ning, M.S.; Andl, T. Control by a hair’s breadth: The role of microRNAs in the skin. Cell. Mol. Life Sci. 2013, 70, 1149–1169.
- Zhang, Y.; Xia, S.; Wang, T.; Wang, S.; Yuan, D.; Li, F.; Wang, X. Chi-miR-30b-5p inhibits dermal papilla cells proliferation by targeting CaMKIIδ gene in cashmere goat. BMC Genom. 2020, 21, 430.
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.N.; Botchkareva, N. V MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway. J. Cell Biol. 2014, 207, 549–567.
- Zhao, B.; Chen, Y.; Yang, N.; Chen, Q.; Wu, X. miR-218-5p regulates skin and hair follicle development through Wnt/β-catenin signaling pathway by targeting SFRP2. J. Cell. Physiol. 2019, 234, 1–13.
- Hu, S.; Li, Z.; Lutz, H.; Huang, K.; Su, T.; Cores, J.; Dinh, P.U.C.; Cheng, K. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci. Adv. 2020, 6, eaba1685.
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653.
- Li, S.; Yin, Y.; Yao, L.; Lin, Z.; Sun, S.; Zhang, J.; Li, X. TNF-α treatment increases DKK1 protein levels in primary osteoblasts via upregulation of DKK1 mRNA levels and downregulation of miR-335-5p. Mol. Med. Rep. 2020, 22, 1017–1025.
- Zhang, J.; Tu, Q.; Bonewald, L.F.; He, X.; Stein, G.; Lian, J.; Chen, J. Effects of miR-335-5p in modulating osteogenic differentiation by specifically downregulating Wnt antagonist DKK1. J. Bone Miner. Res. 2011, 26, 1953–1963.
- Michel, L.D.; Reygagne, P.; Benech, P.; Scalvino, S.; So, S.L.K.; Hamidou, Z.; Bianovici, S.; Pouch, J.; Ducos, B.; Bonnet, M.; et al. Study of gene expression alteration in male androgenetic alopecia: Evidence of predominant molecular signalling pathways. Br. J. Dermatol. 2017, 177, 1322–1336.
- Wang, Z.; Wang, J.; Chen, Z.; Wang, K.; Shi, L. MicroRNA-1-3p inhibits proliferation and migration of oral squamous cell carcinoma cells by targeting DKK1. Biochem. Cell Biol. 2018, 96, 355–364.
- Mirfazeli, E.S.; Arefian, E.; Nadri, S.; Rezazadeh Valojerdi, R.; Kehtari, M.; Zeynali, B. DKK1 expression is suppressed by miR-9 during induced dopaminergic differentiation of human trabecular meshwork mesenchymal stem cells. Neurosci. Lett. 2019, 707, 134250.
- Li, C.; Zhang, P.; Gu, J. miR-29a modulates tumor necrosis factor-α-induced osteogenic inhibition by targeting Wnt antagonists. Dev. Growth Differ. 2015, 57, 264–273.
- Lv, C.; Li, F.; Li, X.; Tian, Y.; Zhang, Y.; Sheng, X.; Song, Y.; Meng, Q.; Yuan, S.; Luan, L.; et al. MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists. Nat. Commun. 2017, 8, 1036.
- Song, Q.; Liu, H.; Li, C.; Liang, H. miR-33a-5p inhibits the progression of esophageal cancer through the DKK1-mediated Wnt/β-catenin pathway. Aging 2021, 13, 1–14.
- Fang, Q.; Liu, T.; Yu, C.; Yang, X.; Shao, Y.; Shi, J.; Ye, X.; Zheng, X.; Yan, J.; Xu, D.; et al. LncRNA TUG1 alleviates cardiac hypertrophy by targeting miR-34a/DKK1/Wnt-β-catenin signalling. J. Cell. Mol. Med. 2020, 24, 3678–3691.
- Xiang, J.; Fu, H.Q.; Xu, Z.; Fan, W.; Liu, F.; Chen, B. lncRNA SNHG1 attenuates osteogenic differentiation via the miR-101/DKK1 axis in bone marrow mesenchymal stem cells. Mol. Med. Rep. 2020, 22, 3715–3722.
- Liu, J.U.N.; Wu, M.; Feng, G.; Li, R.U.I.; Wang, Y.; Jiao, J. Downregulation of LINC00707 promotes osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by regulating DKK1 via targeting miR-103a-3p. Int. J. Mol. Med. 2020, 46, 1029–1038.
- Zhang, Z.; Liu, J.; Shao, Z.; Pu, F.; Wang, B.; Wu, Q.; Zhang, Y.; Zeng, X.; Guo, X.; Yang, S.; et al. In vitro effect of microRNA-107 targeting Dkk-1 by regulation of Wnt/β-catenin signaling pathway in osteosarcoma. Medicine 2017, 96, 27.
- Liao, Y.; Jia, X.; Ren, Y.; Deji, Z.; Gesang, Y.; Ning, N.; Feng, H.; Yu, H.; Wei, A. Suppressive role of microRNA-130b-3p in ferroptosis in melanoma cells correlates with DKK1 inhibition and Nrf2-HO-1 pathway activation. Hum. Cell 2021.
- Di, G.; Kong, L.; Zhao, Q.; Ding, T. MicroRNA-146a knockdown suppresses the progression of ankylosing spondylitis by targeting dickkopf 1. Biomed. Pharmacother. 2018, 97, 1243–1249.
- Xu, Y.; Chen, B.; George, S.K.; Liu, B. Downregulation of MicroRNA-152 contributes to high expression of DKK1 in multiple myeloma. RNA Biol. 2015, 12, 1314–1322.
- Zhou, J.; Lin, Y.; Kang, X.; Liu, Z.; Zhang, W.; Xu, F. microRNA-186 in extracellular vesicles from bone marrow mesenchymal stem cells alleviates idiopathic pulmonary fibrosis via interaction with SOX4 and DKK1. Stem Cell Res. Ther. 2021, 12, 96.
- Cheng, R.; Lu, C.; Zhang, G.; Zhang, G.; Zhao, G. Overexpression of miR-203 increases the sensitivity of NSCLC A549/H460 cell lines to cisplatin by targeting Dickkopf-1. Oncol. Rep. 2017, 37, 2129–2136.
- Xia, Z.; Wang, Y.; Sun, Q.; Du, X. MiR-203 is involved in osteoporosis by regulating DKK1 and inhibiting osteogenic differentiation of MSCs. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5098–5105.
- Jia, Y.; Chen, L.; Guo, S.; Li, Y. Baicalin induced colon cancer cells apoptosis through miR-217/DKK1-mediated inhibition of Wnt signaling pathway. Mol. Biol. Rep. 2019, 46, 1693–1700.
- Jiang, C.; Yu, M.; Xie, X.; Huang, G.; Peng, Y.; Ren, D. miR-217 targeting DKK1 promotes cancer stem cell properties via activation of the Wnt signaling pathway in hepatocellular carcinoma. Oncol. Rep. 2017, 38, 2351–2359.
- Dai, Z.; Jin, Y.; Zheng, J.; Liu, K.; Zhao, J.; Zhang, S.; Wu, F.; Sun, Z. MiR-217 promotes cell proliferation and osteogenic differentiation of BMSCs by targeting DKK1 in steroid-associated osteonecrosis. Biomed. Pharmacother. 2019, 109, 1112–1119.
- Li, Z.; Hu, H.; Zhang, X.; Liu, G.; Ran, B.; Zhang, P.; Liao, M.; Wu, Y. MiR-291a-3p regulates the BMSCs differentiation via targeting DKK1 in dexamethasone-induced osteoporosis. Kaohsiung J. Med. Sci. 2019, 36, 35–42.
- Wu, Z.; Zhang, Y.; Yang, Z.; Zhu, Y.; Xie, Y.; Zhou, F.; Cai, L. Elevation of miR-302b prevents multiple myeloma cell growth and bone destruction by blocking DKK1 secretion. Cancer Cell Int. 2021, 21, 187.
- Zhang, Q.; Wang, F.; Wang, F.; Wu, N. Long noncoding RNA MAGI1-IT1 regulates cardiac hypertrophy by modulating miR-302e/DKK1/Wnt/beta-catenin signaling pathway. J. Cell. Physiol. 2019, 235, 245–253.
- Luan, Y.; Xie, B.; Wei, W. REST-repressed lncRNA NPPA-AS1 regulates cervical cancer progression by modulating miR-302e/DKK1/Wnt/β-catenin signaling pathway. J. Cell. Biochem. 2021, 122, 16–28.
- Li, J.; Feng, Z.; Chen, L.; Wang, X.; Deng, H. MicroRNA-335-5p inhibits osteoblast apoptosis induced by high glucose. Mol. Med. Rep. 2016, 13, 4108–4112.
- Lin, S.C.; Wu, H.L.; Yeh, L.Y.; Yang, C.C.; Kao, S.Y.; Chang, K.W. Activation of the mir-371/372/373 mirna cluster enhances oncogenicity and drug resistance in oral carcinoma cells. Int. J. Mol. Sci. 2020, 21, 9442.
- Weng, J.; Zhang, H.; Wang, C.; Liang, J.; Chen, G.; Li, W.; Tang, H.; Hou, J. MIR-373-3p Targets DKK1 to Promote EMT-Induced Metastasis via the Wnt/β-Catenin Pathway in Tongue Squamous Cell Carcinoma. Biomed. Res. Int. 2017, 2017, 6010926.
- Wang, W.; He, Y.; Rui, J.; Xu, M.-Q. miR-410 acts as an oncogene in colorectal cancer cells by targeting dickkopf-related protein 1 via the Wnt/β-catenin signaling pathway. Oncol. Lett. 2019, 17, 807–814.
- Tang, X.; Lin, J.; Wang, G.; Lu, J. MicroRNA-433-3p promotes osteoblast differentiation through targeting DKK1 expression. PLoS ONE 2017, 12, e0179860.
- Wang, F.; Hu, X.-Y.; Cao, C.; Zhao, Y.-W.; He, S.-H. MiR-488 promotes fracture healing by targeting DKK1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8965–8972.
- Jia, X.; Li, N.; Peng, C.; Deng, Y.; Wang, J.; Deng, M.; Lu, M.; Yin, J.; Zheng, G.; Liu, H.; et al. miR-493 mediated DKK1 down-regulation confers proliferation, invasion and chemo-resistance in gastric cancer cells. Oncotarget 2016, 7, 7044–7054.
- Zhang, H.; Yu, C.; Chen, M.; Li, Z.; Tian, S.; Jiang, J. miR-522 contributes to cell proliferation of hepatocellular carcinoma by targeting DKK1 and SFRP2. Tumor Biol. 2016, 37, 11321–11329.
- Zhou, X.; Wang, Y.; Li, Q.; Ma, D.; Nie, A.; Shen, X. Biochemical and Biophysical Research Communications LncRNA Linc-PINT inhibits miR-523-3p to hamper retinoblastoma progression by upregulating Dickkopf-1 (DKK1). Biochem. Biophys. Res. Commun. 2020, 530, 47–53.
- Liu, L.; Zuo, Y.; Xu, Y.; Zhang, Z.; Li, Y.; Pang, J. MiR-613 inhibits proliferation and invasion and induces apoptosis of rheumatoid arthritis synovial fibroblasts by direct downregulation of DKK1. Cell. Mol. Biol. Lett. 2019, 24, 8.
- Wang, C.; Liao, H.; Zhang, Y.; Cao, Z. MicroRNA-3064-3p regulates the differentiation of cementoblasts through targeting DKK1. J. Periodontal Res. 2018, 53, 705–713.
- Yao, Y.; Hua, Q.; Zhou, Y. CircRNA has_circ_0006427 suppresses the progression of lung adenocarcinoma by regulating miR-6783-3p/DKK1 axis and inactivating Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2018, 508, 37–45.
- Yao, Y.; Zhou, Y.; Hua, Q. circRNA hsa_circ_0018414 inhibits the progression of LUAD by sponging miR-6807-3p and upregulating DKK1. Mol. Ther. Nucleic Acid 2021, 23, 783–796.
- Min, K.; Lee, S.K. EBV miR-BART10-3p Promotes Cell Proliferation and Migration by Targeting DKK1. Int. J. Biol. Sci. 2019, 15, 657–667.
- Dong, M.; Gong, L.; Chen, J.; Zhang, X.; Zhang, Y.; Hui, D. EBV-miR-BART10-3p and EBV-miR-BART22 promote metastasis of EBV-associated gastric carcinoma by activating the canonical Wnt signaling pathway. Cell. Oncol. 2020, 43, 901–913.
More