Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Camila Xu and Version 4 by Camila Xu.
Programmed cell death 1 (PD-1) and its ligands PD-L1 and PD-L2 are receptors that act in co-stimulatory and coinhibitory immune responses. Signaling the PD-1/PD-L1 or PD-L2 pathway is essential to regulate the inflammatory responses to infections, autoimmunity, and allergies, and it has been extensively studied in cancer.
- PD-1
- PD-L1
- PD-L2
- asthma
- BTLA
1. Introduction
Allergic diseases are characterized by hyperreactive immune responses to otherwise innocuous antigens in the general population, the incidence and prevalence of which are increasing worldwide, particularly in the developed world. In some western countries, every third child needs medical attention from a physician caused by allergic diseases [1][2][3][1,2,3].
The allergic inflammatory process involves different cell types that release a range of inflammatory mediators and cytokines. Th2 cytokines, including IL-4, IL-5, and IL-13, induce IgE production, while IL-5 promotes eosinophil infiltration [4][5][4,5]. Other factors, including Th1 cytokines, may sometimes affect the clinical manifestation creating a potential tolerance induction or hyperactivity [6]. Moreover, the progression of allergy disease is mediated by the innate immune system on the respiratory or gastrointestinal tract mucosa. Indeed, antigen-presenting cells (APCs) become activated, and epithelial alarmins (IL-25, IL-31, IL-33, and TSLP) are released due to activation of the airway epithelial cells in response to allergens [7], leading to activation of Th2/mast cell/eosinophil/eosinophil-mediated allergic pathology, including IL-4 and IL-5. Another pathway is the Th1/Th17/neutrophil-mediated response to chronic innate immune activation and irreversible airway obstruction [8][9][8,9]. In addition, the high cellular plasticity in the context of type-2-driven inflammation refers to the capacity of T-helper cell subpopulations to respond depending on environmental signals differentiating into other effector cell types. For example, in chronic asthma, Th1 and Th2 cytokines contribute to airway wall remodeling. Another group of cells that respond to environmental signals are group 2 innate lymphoid cells that exhibit a dynamic phenotype in allergic airway inflammation [10]. Allergic diseases such as asthma are heterogeneous with multiple conditions. The molecular phenotypes defined based on the predominant inflammatory cell are endotypes. In this context, the type 2 immune response can be categorized into different subgroups, such as IL-5-high, IL-13-high, or IgE-high, and others [11].
In contrast, IL-10 downregulates the inflammatory process. IL-10 acts directly on CD4+ T cells, primarily by down-regulating IL-2 and other cytokines produced by Th1 cells. It also inhibits Th2 cell production of IL-4 and IL-5. On dendritic cells (DCs), monocytes and macrophages, and APCs, the effects of IL-10 include inhibition of the production of free mediator molecules in the medium, such as receptor decreases, or abolished antigen presentation and phagocytosis [12]. Thus, IL-10 plays an essential role as a regulatory molecule in both innate and adaptive immune responses, leading to immune tolerance or dampening tissue inflammation in humans. Similarly, the programmed cell death 1 (PD-1) and its ligands PD-L1 and PD-L2 play an essential role in regulating T cells activation. It controls the immune balance preventing the accumulation of self-reactive T cells [13]. PD-1 has been characterized as a negative regulator of conventional CD4+ T cells. In addition, PD-L1 and PD-L2 have an essential but oppositive role in modulating and polarizing functions of T-cells in hyperreactivity [14].
2.
2. PD-1 and Its Ligands PD-L1 and PD-L2
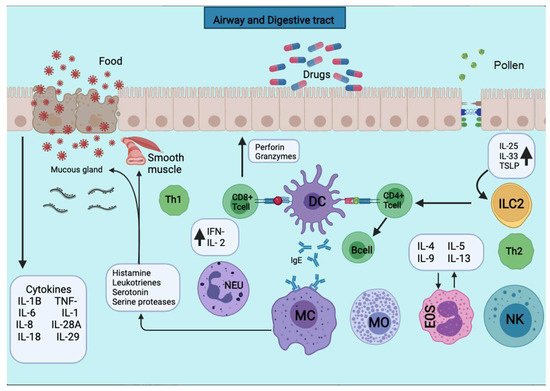
Figure 12. APCs present the antigens to the CD4+ T cells subpopulation for activation. APCs take up the first sensitizing contact with the antigen, bind it to MHC class II molecules, and expose it to the membrane. APCs take up the first sensitizing contact with the antigen, which processes and binds it to MHC class II molecules and exposes it to the membrane. Several cytokines such as TNF-a, IL-1, IL-4, IL-5, IL-6, IL-13, MIP-1a, and various colony-stimulating factors such as GM-CSF and IL-3 are released. The triggering of the allergic reaction is stimulated by the production of IgE by B lymphocytes. The production of IgE stimulates the triggering of the allergic reaction by B lymphocytes. The IgE will bind to mast cells and basophils for degranulation and the release of several mediators.
3.
3. PD-1 and Its Ligands in Allergy Diseases
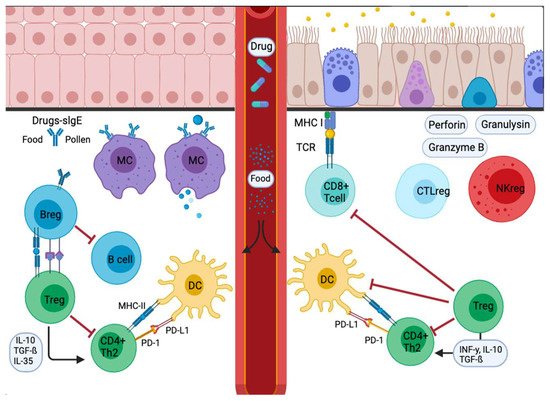
Figure 23. Signal inflammatory cascade allergy response in mucosas. Incoming allergens are taken up and processed by APCs or antibodies. Allergens cross the mucosa, then move to the lung pleura or intestine. In the sensitization stage, APCs present the antigen and give an immediate response. Others migrate to the draining lymph nodes, where the processed allergen is presented to naïve T and B cells, responses characterized and influenced by the cytokines secreted and the presence or absence of cell-bound co-stimulatory molecules. Decreased stimulus and/or the presence of IL-10, IL-35, or TGF-β triggers Treg cell transformation. In addition, APCs lead to the production of cytokines that regulate the change and expression of coinhibitory receptors such as PD1/PD-L1.