Infectious bronchitis virus (IBV) is the causative agent of multi-systemic infection in the respiratory, reproductive and renal systems, which is similar to the symptoms of various viral and bacterial diseases reported in chickens. Currently, the live attenuated and killed vaccines are applied for the control of IBV infection; however, the continual emergence of IB variants with rapidly evolving genetic variants increases the risk of outbreaks in intensive poultry farms.
- infectious bronchitis virus
- vaccination
- immune system
1. Introduction
2. Vaccination
| Name of the Vaccine | Route of Delivery | Characteristics | |
|---|---|---|---|
| 1. Live attenuated IBV or | Aero nasal spray | Serial attenuation of virulent IB strain for weakened | |
| Live IBV vaccines | In Ovo route | virulence [16][17]. | virulence [32,33]. |
| Orally | |||
| Subcutaneous (S/C) | |||
| 2. Killed or inactivated | IM injection | Inactivated by chemical treatment or heat treat to kill the | |
| IB vaccines | S/C | virulence of strain [18]. | virulence of strain [34]. |
| 3. Viral Vector vaccine | In ovo route | Recombinant rNDV/APMV-2 expressing the S protein of | |
| IBV strain Mass-41 (rNDV/APMV-2/IBV-S) [19]. | IBV strain Mass-41 (rNDV/APMV-2/IBV-S) [35]. | ||
| 4. DNA vaccine | Mucosal/Orally | IBV-DNA vaccine carrying S1-protein and/or N-protein constructs | |
| IM injection | the respective vector [20][21][22][23]. | the respective vector [36,37,38,39]. | |
| Intranasal | |||
| In ovo route | |||
| 5. Recombinant protein (sub-unit) |
Intraocular-nasally IM injection |
Water-in-oil emulsified recombinant S-ectodomain protein [24]. | Water-in-oil emulsified recombinant S-ectodomain protein [40]. |
| Second heptad repeat (HR2) region of S protein were | |||
| repeatedly co-displayed in the Self-assembling | |||
| Protein Nanoparticle (SAPN) [25]. | Protein Nanoparticle (SAPN) [41]. | ||
| 6. Multi-epitope-based | Oral | Using attenuated S enterica serovar Typhimurium strain [26]. | Using attenuated S enterica serovar Typhimurium strain [42]. |
| peptide vaccine | Mucosal | Recombinant DNA: The EpiC gene was presented in | |
| (Lactococcus lactis bacterial system) |
Intranasal | Lactococcus lactis NZ3900 with 3 recombinant strains expressing EpiC gene [27]. | Lactococcus lactis NZ3900 with 3 recombinant strains expressing EpiC gene [43]. |
| 7. VLP-based IBV vaccine or | IM immunized | Efficient mucosal immune response [28] | Efficient mucosal immune response [44] |
| chimeric VLP vaccine |
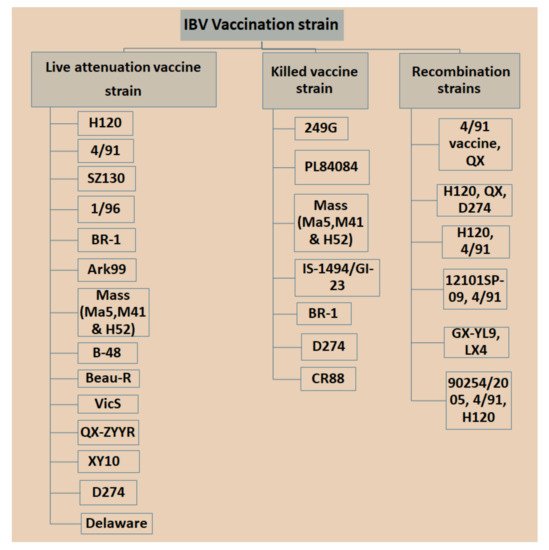
Currently, IBV vaccination programs have gained more attention with respect to the use of low-virulent, live or inactivated killed vaccines with the aim of booster shots at certain times to increase immunity and reduce the antagonistic response of epithelial cells in the respiratory region [29][89]. However, there is a significant limitation in applying live IBV vaccines because the attenuation of the vaccine is naturally deficient with respect to its capability in stimulating a mucosal immune response [30][31][90,91], which is a critical part of controlling IBV infection since killed vaccine can be an option [32][33][92,93]. Nonetheless, it was possible that inactivated killed vaccines can be applied to stimulate t mucosal immune responses once combined with several nanoparticles [32][92]. Different types of IBV vaccines are available in the market, which may vary in vaccine strains, and in nature based on local isolate and recombination in strains isolated from different countries with special legal legislation (Figure 13). Bijlenga et al. [34][49] described the earlier development of the H strain of IBV containing both the H52 and H120 due to its better capability of heterologous cross-protection against different serotypes of IBV. Further studies have revealed that heterologous IBV vaccines are also more effective for immunizing the 793B-type of variant that has been evidenced to be long lasting with live attenuated IB vaccines and are effectively applied against Italy 02 and QX stain [35][36][37][94,95,96]. The modified live vaccines and inactivated oil emulsions are available for a few serotypes such as Massachusetts, Connecticut and Arkansas in North America. The California strains and Georgia 98 vaccines are collected from the USA.
3. Immune Response against IBV
3.1. Local Immune System
A vaccine requires a certain period of time in order to elicit a protective immune response in avian hosts. Moreover, passive immunization can induce immunity from maternal derivative’s antibody (MDA), which is particularly supportive during the early stages of life [43][106]. The structure and function of birds’ immune systems are distinctly different from human immune systems due to their virtue of possessing extra lymphatic organs such as the bursa of Fabricius and the thymus responsible for humoral and cellular immunity, respectively. Furthermore, the birds have carried the secondary peripheral organs of the lymphatic system, for example, the Harderian gland (HG), conjunctiva associated lymphoid tissue (CALT), head associated lymphoid tissue (HALT), gut-associated lymphoid tissue (GALT) and bronchus associated lymphoid tissue (BALT), spleen and cecum tonsils, respectively [44][107], showed in Figure 24. These assemblies are regularly enmeshed in a chicken’s immune response, especially in the respiratory mucosal system during IBV infection.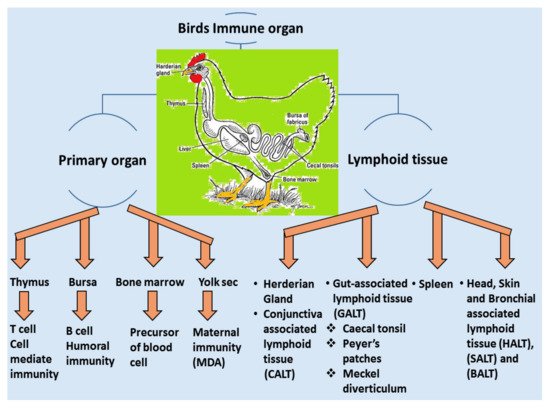
3.2. Adaptive Immune System
3.2.1. Humoral Immunity
3.2.2. Cell-Mediated Immunity (CMI)
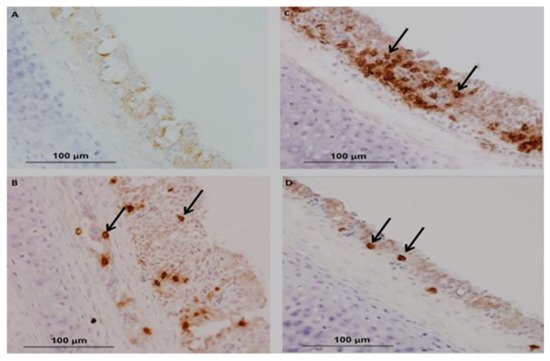
In chickens, CMI is one of the essential immunoregulatory weapons during IBV infection, especially for aiding the clearance of viruses, decline of infection and reducing virus shedding and vaccine development [49][146]. The evaluation of cellular arms is performed by lymphocytic transformation assays, cytotoxic lymphocyte activity [50][147], delayed-type hypersensitivity [51][148] and natural killer cell activity [52][149]. Histological lesions of CMI responses are performed by T-cell infiltration in the respiratory and renal tissues of IBV-infected chickens [53][150]. The experimental studies have shown a positive relationship between lymphoproliferative responses and resistance to challenge infection [23][39]. Alternative studies have been published on mouse monoclonal antibodies (mAbs) that differentiate between T-lymphocytes and are used to evaluate the role of T-cells in viral clearance [54][151]. N and S genes have a specific protein response associated with the stimulated virus-specific protective immunity of CTLs, which is characterized by the reduction in viral load and clearance of the virus from circulation [55][56][152,153]. A marked increase in CD4+ and CD8+ T-cells has been described as the recombinant S1-gene associated with the induction of cellular immunity of specific IBV vaccines [57][154]. Guo et al. [58][155] reported that IB vaccination with N gene-based DNA vaccine significantly increased the number of CD4+ and CD8 + T cells in peripheral blood mononuclear cells (PBMCs). The existence of CD8+ cytotoxic T lymphocytes (CTL) signifies an essential relationship for reducing infection and resembles a decrease in clinical signs by the action of major histocompatibility complex (MHC), and lysis is facilitated by CD8+CD4 cells [59][156]. Consequently, the major histocompatibility complex organized cytotoxic T lymphocytes (CTL), and the cytokine activities of chickens participated during the early stages of IBV infections [60][157]. Several studies have been conducted on tracheal immunity induced by live vaccines, and they found that all vaccines induced significantly higher expression of CD4+ and CD8+ compared with unvaccinated birds using a nephron-pathogenic IBV strain [28][44]. In the following year, other studies have reported that CD4+ cells are recruited into the trachea earlier than CD8+ on 5 dpi (days of post-infection) [61][158], which agrees with the findings by Kotani et al. [62][159] who recognized that the frequency of CD4+ and CD8+ cell numbers significantly peaked at 5 dpi when using a virulent IBV strain. In contrast, studies reported that CD8+ cells were recruited into the trachea earlier than CD4+ cells after infection with virulent 793B [63][160] or live attenuated IBV vaccine [28][44] or a combination of live attenuated vaccine with a booster dose of an inactivated vaccine [64][161]. CD8+ memory T cells have greatly protected the newborn chicks from acute IBV infection at 4 dpi and mild clinical symptoms show at 5 dpi [59][156]. Even though the adoptive transfer of CD4 + T cells could not be significantly protected in the early stage of IBV infection, primed αβ T cells carrying CD8+ T cells are critical in protecting chicks from IBV infection [55][152]. Chhabra et al. [28][44] reported that the protection against Q1-IBV strain changes the quantity of CD4+ and CD8+ cells in the trachea using immunohistochemistry. The results showed that the overall patterns of CD8+ cells are dominant compared to those of CD4+ cells in the two vaccinated groups. The kinetics of CD4+, CD8+ and the IgA-carrying B lymphocytes in the trachea are shown in Figure 35B–D) compared with control Figure 35A in vaccinated groups as differences may have a close relationship with the IBV-specific strains.