In 1924, to understand the processes involved in developmental biology, Spemann and Mangold transplanted a blastopore lip between different ectodermal regions of amphibian embryos. The transplanted dorsal tissue differentiated mostly into a notochord, while the ectoderm of the host dorsal tissue that was sitting above the transplanted region (blastopore lip) was induced and differentiated to form a Siamese twin containing dorsal tissues such as somites and a neural plate, which would form the central nervous system, forming the bulk of a second axis. The major findings were that the transplant had altered the fate of the overlying cells and that the neural folds were built from recipient cells and not donor cells. Spemann and Mangold discovered the organizing center in the dorsal blastopore lip of amphibians. This center consists of a cluster of cells in the developing embryo that have the ability to interact and instruct morphogenesis in the surrounding cells during gastrulation. When transplanted to the ventral side of the embryo, the center will induce the formation of a secondary axis, promoting the development of the central nervous system, organs, and tissues, as well as the formation of the main body axis. Spemann and Mangold found the first evidence of the organizing center, thereafter called the “Spemann organizer”, and its major role in the development of vertebrates. This discovery also introduced the concept of induction in embryonic development, which refers to the method used by specific cells to affect the fate of other embryonic cells. A major milestone had been achieved for developmental biology.
1. Introduction
In 1924, to understand the processes involved in developmental biology, Spemann and Mangold transplanted a blastopore lip between different ectodermal regions of amphibian embryos
[1]. The transplanted dorsal tissue differentiated mostly into a notochord, while the ectoderm of the host dorsal tissue that was sitting above the transplanted region (blastopore lip) was induced and differentiated to form a Siamese twin containing dorsal tissues such as somites and a neural plate, which would form the central nervous system, forming the bulk of a second axis
[1][2][1,2]. The major findings were that the transplant had altered the fate of the overlying cells and that the neural folds were built from recipient cells and not donor cells. Spemann and Mangold discovered the organizing center in the dorsal blastopore lip of amphibians
[1]. This center consists of a cluster of cells in the developing embryo that have the ability to interact and instruct morphogenesis in the surrounding cells during gastrulation
[3]. When transplanted to the ventral side of the embryo, the center will induce the formation of a secondary axis, promoting the development of the central nervous system, organs, and tissues, as well as the formation of the main body axis
[3][4][5][3,4,5]. Spemann and Mangold found the first evidence of the organizing center, thereafter called the “Spemann organizer”, and its major role in the development of vertebrates. This discovery also introduced the concept of induction in embryonic development, which refers to the method used by specific cells to affect the fate of other embryonic cells
[1][2][3][1,2,3]. A major milestone had been achieved for developmental biology.
Years after the findings of Spemann and Mangold, Harland and Gerhart
[6], using the amphibian
Xenopus laevis, further studied the functions of the organizer and concluded the following. First, the organizer deactivates or activates the surrounding cells to differentiate and start morphogenesis. Second, the three germ layers (endoderm, mesoderm, and ectoderm) of the developing embryo are signaled and affected by the organizer. Third, the three germ layers will respond to the organizer’s signals
[6]. Since the initial research carried out mainly by Spemann and Harland, in recent decades many experiments have been carried out to provide more knowledge about the function of the Spemann–Mangold organizer during embryonic development
[3][5][7][8][9][10][3,5,7,8,9,10]. It has been found that the amphibious Spemann–Mangold organizer has developmental analogues in other vertebrates
[11]. In teleost fish this is an embryonic shield, in avian embryos it called Hensen’s node, and in mammals refer as node. However, the AVE (anterior visceral endoderm); an extra embryonic tissue also known as second organizer in mammals (
Figure 1)
[12][13][14][12,13,14]. This signifies that the results obtained by studying the Spemann–Mangold organizer can be extrapolated to all vertebrates
[5]. All vertebrates are thought to have an organizer that releases inductive signals, responsible for body plan arrangement
[5]. The organizer’s distinct cell populations differentiate into various tissues and can be subdivided into head, trunk, and tail organizers based on their different inducing abilities
[15].
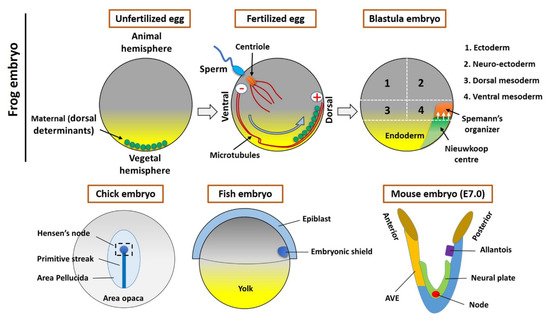
Figure 1. Organizer formation and fate map of Xenopus embryos (Upper panel). In an unfertilized egg, the maternal determinants (small vesicles) are located in the vegetal hemisphere. The sperm entry triggers the fertilization process and small vesicles associated with kinesin (motor protein) moved toward the opposite site (the plus end of microtubules) of the sperm entry point. These vesicles contain the components of Wnt signaling, which establishes the Nieuwkoop center and Spemann organizer during the cleavage stages of early embryogenesis. The lower panel shows the Hensen’s node (Chick embryo), embryonic shield (Fish embryo), and node/AVE (Mouse embryo).
2. The Spemann’s Organizer and Homologous Tissue in Vertebrates
In all vertebrates, the organizer (or equivalent) is functionally conserved. The transplanted Hensen’s node can induce neural tissue in fish, mammalian, and amphibian embryos
[3][12][16][3,12,18], which indicates that homologous tissue might be functionally conserved among vertebrate species. In this section, we mainly focused on embryonic processes involved in forming amphibian organizer. Additionally, a brief discussion regarding the equivalent tissue in vertebrates is provided.
2.1. The Maternal Determinants Establish the Organizer
The formation of Spemann’s organizer involves a series of complex intercellular events that begin right after fertilization. The overall process of organizer formation can be described by some sequential/parallel embryonic events that can be described as two steps. Cortical rotation is the first intracellular event in the fertilized egg that allows for the shifting of maternal determinants from the vegetal to the animal region of the embryo (
Figure 1), which establishes the dorsal side (the opposite to sperm entry)
[6][10][6,10]. The dorsal determinants located in membrane vesicles in the vegetal pole of the embryo are transported to the dorsal side by cortical microtubules and kinesins (
Figure 1)
[10]. These vesicles are associated with Dishevelled (Dvl/Dsh), a component of the Wnt signal transduction pathway. Therefore, the Wnt pathway will be activated, which will lead to the accumulation and stabilization of β-catenin on the dorsal side (dorsal blastomeres) (reviewed in
[10]). This is tightly regulated by the Adenomatous Polyposis Coli (APC), Glycogen synthase kinase 3 (GSK3), and Axin to form a destruction complex and induces phosphorylation-mediated ubiquitination and proteasomal degradation
[16][17][18,19]. Activation and binding of Wnt ligands to Frizzled (FZD) receptors and co-receptors of the LDL Receptor-Related Protein (LRP) inhibit GSK3 and the destruction complex; hence, β-catenin can accumulate, translocate, and stabilize in the nucleus of the dorsal cells of a blastula stage embryo
[18][20]. β-catenin is required for the activation of initial organizer marker genes like
goosecoid (
gsc),
chordin (
chrd),
noggin (
nog), and
Xnrs [19][20][16,21]. The second event includes the signal generated by maternal factors like β-catenin; these maternal factors activate dorsal determinants. Furthermore, the maternal T-box transcription factor VegT and the maternal TGF-β-family growth factor Vg1 are located on the vegetal hemisphere
[21][22]. At the midblastula stage, mesoderm induction begins, which activates many mesoderm- and endoderm-specific genes in the dorsal and ventral domains of the marginal zone
[6]. On the ventral side of the embryo, VegT and Vg1 downregulate the levels of nodal-related genes to originate the ventral mesoderm
[3][6][22][23][3,6,23,24]. Conversely, on the dorsal side, the Wnt/β-Catenin levels coupled with VegT and Veg1 induce the homeobox transcriptional activator Siamois (Sia)/Twin (Xtwn) in the blastula Chrd and Nog-expressing (BCNE) center and regulate the expression of nodal-related genes including
Xnr1,
Xnr2,
Xnr4,
Xnr5, and
Xnr6 [11][24][11,25]. In particular,
Xnr5 and
Xnr6, acting in combination with Wnt/β-catenin signaling, form the Nieuwkoop center (NC) in the dorsal–vegetal region (
Figure 1) of the blastula
[25][26]. Consequently, the NC with Sia and Xtwn will induce the expression of pre-organizer genes such as
gsc,
Frzb-1, and
dickkopf-1 (
dkk-1)
[26][27]. Finally, the Cerberus (
cer) gene will be upregulated, promoting the formation of overlying mesoderm and the establishment of the Spemann’s organizer, during gastrulation, at the dorsal mesoderm of the embryo
[11][24][11,25].
2.2. The Homologous Structure of Organizer in Other Vertebrates
All vertebrates have a position-specific group of cells (collectively refer as organizers) that are absolutely essential in embryonic development. During embryonic development of the chick, after the mid-streak stage, the top region of the streak thickens and forms the Hensen’s node. Later on, these cells assembled to the midline of the mesoderm to form a notochord
[27][28]. Whereas the embryonic shield forms at the blastoderm margin region (future dorsal side) within the fish embryo, it resembles the dorsal lip of the amphibian embryo
[28][29]. A streak is located on the posterior side of the mouse embryo, wherein the AVE is located at the anterior side. The node located on the ventral surface (during embryonic day 7 (E7)) (
Figure 1)
[29][30][30,31]. This species-specific organizer (s) secrete molecules are most common across the vertebrates, including BMP, Wnt, and Nodal inhibitors
[6], and forms prechordal plate and notochord
[12][31][12,32]. Several studies reported that the organizer’s cells mostly differentiate into axial mesoderm and notochord (or chordamesoderm)
[6][8][14][30][32][6,8,14,31,33]. However, the organizer influences fate of surrounding cells (or tissues) by extracellular secretion of active molecules
[3][6][14][3,6,14]. During development, the organizer (or homologous) tissue transforms into notochord and prechordal mesendoderm (or derivate) that involves several signaling pathways to acquire final fate
[6][33][34][35][6,34,35,36]. However, in the present study, we focused on organizer-induced or mediated embryonic development.