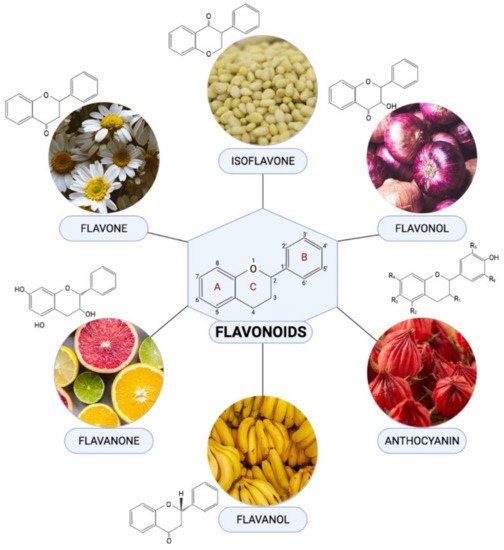
Flavonoids are one of the most diverse families of bioactive phytochemicals, with over 9000 different compounds. According to IUPAC Recommendations (2017), the term “flavonoid” refers to compounds that have the basic structure of phenyl-substituted propylbenzene derivatives with C15 skeleton, C16 skeleton, and flavonolignans with C6–C3 lignan precursors. Flavonoids are divided into six subclasses; isoflavones, flavones, flavanols, flavonols, flavanones, and anthocyanins, are abundant in plants and their metabolic routes have been thoroughly explored using biochemical and molecular approaches.
1. Introduction
Diabetes mellitus (DM) is one of the most deadly non-communicable diseases that leads to extensive impairments of organs and body functions. The increasing incidence of DM and its related complications have contributed to the surge of morbidity and mortality rate. DM affects about 463 million people aged between 20 to 79 years in 2019, and this figure is expected to climb up to 700 million by 2045 [1]. Moreover, DM is also one of the root causes for the development of cardiovascular diseases (CVD), which further exacerbate the mortality risk among patients with DM [2]. One of the complications resulting from chronic DM is diabetic cardiomyopathy (DCM). DCM is a cardiac pathological condition in patients with DM characterized by the appearance of aberrant myocardial morphology and cardiac functions in the truancy of other factors, such as coronary artery disease, hypertension, and prominent valvular disease [3]. Due to DCM, patients with DM are more likely to suffer from heart failure compared to their healthy counterparts [3]. This is why DCM is one of the most devastating consequences directly caused by DM.
The heart has a high energy consumption in order to efficiently pump and supply blood throughout the body. Hence, it has a high density of mitochondria population to fuel its activities. However, this dependence exposes the heart to deleterious consequences when mitochondrial malfunction occurs. Mitochondria serve a critical part in oxygen metabolism, hence it is crucial to understand the effects of their dysfunction in patients suffering from metabolic disorders, particularly diabetes [4]. In DCM, the minimal glucose utilization will shift to fatty acid, leading to energetic inefficiency [5]. Since the mitochondria lost its efficiency in energy production, mitochondrial dysfunction will then follow. The role of mitochondrial dysfunction in the progression of DCM has been well established in earlier studies [6][7][8][6,7,8]. As the heart contains a high amount of mitochondria, cardiac mitochondrial dysfunction can lead to cardiac oxidative stress which aggravates the development of DCM. Indeed, the diabetic patients heart mitochondria are typically found to have deteriorated in number and structure, exhibiting increased reactive oxygen species (ROS) emission, and compromised mitochondrial respiratory capacity in the mitochondria [9]. Thus, treatment targeting mitochondrial-induced oxidative stress is very crucial in suppressing DCM.
For more than 40 years, the pathogenesis and mechanisms involved in DCM’s development and progression has been well-studied and documented as well as of its preventive measures and potential therapeutic agents. Despite this, effective remedies for preventing and treating DCM remains unclear [10]. The need of having a treatment for DCM is of utmost importance considering that there is no specific treatment targeting DCM up to the moment [11][12][11,12]. Presently, diabetes management is only based on combination of lifestyle modification and therapeutic medications to regulate blood glucose level through glucose-lowering agents or insulin replacement therapy as well as with management of cardiovascular complications [12][13][12,13].
2. Therapeutic Role of Flavonoid in Alleviating Mitochondrial Dysfunction-Induced Oxidative Stress in Diabetic Cardiomyopathy
Flavonoids are one of the most diverse families of bioactive phytochemicals, with over 9000 different compounds. According to IUPAC Recommendations (2017), the term “flavonoid” refers to compounds that have the basic structure of phenyl-substituted propylbenzene derivatives with C15 skeleton, C16 skeleton, and flavonolignans with C6–C3 lignan precursors
[14][88]. Flavonoids are divided into six subclasses; isoflavones, flavones, flavanols, flavonols, flavanones, and anthocyanins, are abundant in plants and their metabolic routes have been thoroughly explored using biochemical and molecular approaches
[15][16][89,90]. Many plants, including pomelos, blueberries, roselle, oranges, grapefruit, lemons, and limes, are all rich in flavonoids
[17][91].
Flavonoids have been demonstrated to alleviate pathological disorders, such as diabetes, cancer, obesity, and cardiovascular diseases. Flavonoids are abundant plant-based natural compounds with a good potential for medicinal and biological actions. These compounds showed the ability to exert anti-oxidative, anti-inflammatory, anti-fibrotic, and anti-apoptotic activities as reported previously
[18][19][20][92,93,94]. Given the role of mitochondrial-induced oxidative stress in the progression of DCM, this review aims to summarize mechanisms of action of flavonoids in alleviating DCM by targeting mitochondrial-induced oxidative stress.
Figure 14 demonstrated the chemical structure of flavonoid subclasses.
Figure 14. Chemical structures and example of sources where they are found abundant in for each flavonoid subclasses.
2.1. Flavones
Flavone is one of the significant flavonoid subclasses. They are found as glycosides in celery, parsley, red peppers, mint, and ginkgo biloba. This group of flavonoids includes luteolin, apigenin, and tangeritin
[21][95]. They have a double bond between positions 2 and 3 of the main C ring, as well as a ketone in position 4. The hydroxyl group at position 5 of the A ring is found in the majority of flavones from vegetables and fruits, but hydroxylation in other parts, most commonly in position 7 of the A ring or 3′ and 4′ of the B ring, varies depending on type of the vegetable or fruit
[22][96].
Luteolin is one of the most prevalent flavones that can be found in variety of vegetables, fruits and herbs such as apple, cabbage, carrot, tea, and celery. A previous study by Yang and colleagues
[23][97] which utilizes streptozotocin-induced diabetic rodents ischemia reperfusion model showed that luteolin treatment, at 100 mg/kg dose, was able to increase cardiac MnSOD and endothelium nitric oxide synthase (eNOS) expression as well as decrease Ca
2+ induced mPTP opening and ΔΨm.
In addition, myricitrin, a flavone that can be found in abundance in berries and teas, has been proven to suppress high glucose-induced superoxide production in mitochondria, depolarization of mitochondrial membrane potential and restored mPTP formation in diabetic cardiomyopathy via in vitro study
[24][98]. Another study has reported that flavonoid from
Abroma augusta L. (Malvaceae) leaf extract containing predominantly rutin improved co-enzyme Q
9 and Q
10 levels in the mitochondria by acting as antioxidants through scavenging ROS and thereby inhibit lipid peroxidation
[25][99].
2.2. Isoflavones
Isoflavones are phytocompounds with a chemical composition based on the 3-phenyl chromen-4-one backbone. Isoflavones are secondary plant metabolites extensively studied for its wide range of therapeutic effects, including antioxidant, chemopreventive, anti-inflammatory, anti-allergic, antibacterial, and cardioprotective effects
[26][27][100,101]. The highest content of isoflavones is identified to be in roots and seeds. Other medicinal plants with high isoflavones content include red clover, dyer’s broom, lucerne, and sohphlang flax. Beside soy, other legumes such as lupin beans, kudzu, barley, and fava beans are rich in isoflavones
[28][29][102,103]. The most important types of isoflavones are genistein, daidzein, glycitein, formononetin, biochanin A, and equol
[30][104].
Isoflavone has been reported to alleviate mitochondrial-induced oxidative stress on DCM. Recently, Upadhayay et al.
[31][24] reported that isoflavone was able to reduce mitochondrial-induced oxidative damage by reducing ROS generation in mitochondria and depolarization of mitochondrial membrane through silent information regulator 1 (SIRT-1) pathway or PPAR-α, which further attenuated mitochondrial dysfunction, thus conserving cardiomyocytes health. Besides, recent study conducted by Laddha and colleagues has confirmed that streptozotocin-induced type 1 diabetic rats were shown to maintain AMPK and SIRT-1 levels to normal levels whereby both activities are important in controlling free fatty acid uptake as well biogenesis in cardiac mitochondria
[32][105].
However, studies made on the effects of isoflavones on cardiac diabetes model are rather meagre in number. Yet, we can still refer to its therapeutic effect on other cardiac pathology models as well. There are several studies that reported favorable effects of isoflavones on cardiac mitochondria. Recently, isoflavones was shown to give positive effects on mitochondria by alleviating the excessive mitochondria Ca
2+ uptake in isolated heart
[33][106]. Apart from that, isoflavones was also capable in improving disturbance in ΔΨm as well as reduction of intracellular ROS release, thus proving that isoflavone was able limit oxidative stress induced by mitochondria
[34][35][107,108]. In addition, isoflavones also alleviate ΔΨm loss as well as curbing mPTP opening which exhibiting cardiac protective effect
[35][108].
2.3. Flavonol
Flavonoids with a ketone group are known as flavonols. Flavonols can be found plentiful fruits and vegetables. Kaempferol, quercetin, myricetin, and fisetin are among the most widely studied flavonols and they can be found in abundant in common daily diet including in onions, kale, lettuce, apples, and berries. Flavonol consumption has been proved to a variety of health advantages, including antioxidant potential and a lower risk of cardiovascular disease. Flavonols have a hydroxyl group in position 3 of the C ring, which can be glycosylated, unlike flavones. Flavonols have a vast spectrum of methylation and hydroxylation forms, and they are the most prevalent and largest subclass of flavonoids in fruits and vegetables based on their many glycosylation patterns
[36][109].
Flavonol has a vast potential in protecting heart mitochondria. Earlier study has revealed that flavonol was able to enhance mitochondrial biogenesis by increasing mitochondrial DNA content via upregulation of nuclear factor erythroid 2-related factor, Nrf-1, Nrf-2, and mitochondrial transcription factor A (TFAM) expression
[37][110]. Furthermore, flavonol also was capable to improve complexes I, III, and IV activities as well as upregulate expression of UCP-2 and UCP-3
[37][110]. These findings show that flavonols have promising capability in protecting against cardiovascular disease development.
Indeed, previous study reported that flavanols found in
Abroma augusta L. family of Malvaceae including rutin, myricetin, and quercetin have been proven to revive the ubiquinones (co-enzyme Q) function, which is important in electron carriers’ distribution within cell organelles chiefly and thus reduce ROS production in myocardial mitochondrial of T2DM
[25][99]. In another cardiac study model, quercetin also was found to control free FAO by modulating phosphorylation of AMPK via upregulation of AMPKα2, PPARα, and PCG-1α genes where these genes are crucial in altered energy metabolism mechanisms
[38][39][111,112]. The derivative of myricetin, dihydromyricetin, could boost mitochondrial performance in streptozotocin-induced diabetic rodents, thus reducing oxidative stress. In this study, the ATP levels and complexes Ι/ΙΙ/ΙΙΙ/ΙV maneuver in ETC as well as ΔΨm were enhanced when treated with dihydromyricetin in the cardiomyocytes
[40][113].
Recently, Ni and colleagues
[41][23] have demonstrated that flavonol icariin could upregulate Apelin, the gene in myocardium and the mitochondrial matrix gene Sirt3, hence elevates the ΔΨm and reduces mitochondria ROS production. Another study has shown that flavonol from quercetin could induce peroxiredoxin-3 (Prx-3) expression, a mitochondrial antioxidant, causing a significant decrease in myocardial biomarkers for mitochondrial uncoupling and redox stress, UCP3 protein expression, and reduction of cardiac thioredoxin-2 (Trx-2) expression as well as thioredoxin reductase-2 (TrxR2) activity. Therefore, it could upregulate the expression of Nrf2/Nrf1 and consequently elevate Prx-3 expression
[42][114]. An in vivo study conducted by using
Murraya koenigii (curry) and
Moringa oleifera leaf extract that contain quercetin and kaempferol has reported that these flavonols were able to enhance the expression SOD1 gene, PGC 1α gene, and ATPase and improve mitochondrial function in the diabetic heart
[43][115].
The disruption of mitochondrial transmembrane potential is one of the causes that lead to mitochondrial induce oxidative stress. Taxifolin (dihydroquercetin), a subclass of flavonol could restore mitochondrial transmembrane potential in H9c2 cell lines (Sun et al. 2014). Wu and the team
[40][113] have reported that the dihydromyricetin could enhance the ATP content levels, citrate synthase activity and complex Ι/ΙΙ/ΙΙΙ/ΙV and ATP synthase activities as well as increase in ΔΨm.
2.4. Flavanol
Flavanols are the 3-hydroxy derivatives of flavanones commonly known as dihydroflavonols or catechins. They are a multi-substituted and highly diverse subclass of flavonoids.
[22][96]. Due to the hydroxyl group attached to position 3 of the C rings, flavanols are also known as flavan-3-ols. There is no double bond between positions 2 and 3, unlike many flavonoids. Fruits such as bananas, pears, apples, blueberries, and peaches are rich in flavanols.
In earlier study, flavanol has appear to protect heart mitochondria via various mechanisms. This includes protection effect of flavanol via meddling with ETC complexes activities through deprivation of complex I activity, consequently mitochondrial membrane depolarization which then of ROS production (NO and H
2O
2)
[44][116]. This has been corroborated by previous study on T2DM model where epigallocatechin-3-gallate (EGCG), a flavanol, attenuated myocardial deterioration and showed beneficial effects on myocardial mitochondrial components. EGCG has been demonstrated to revive complex I, III, and IV, as well as voltage-dependent anion-selective channel 1 (VDAC1) activities that produce major ROS. Mitochondrial DNA (mtDNA) copies and the mitochondrial dehydrogenase activity were significantly revived in treatment model
[45][117]. This evidence indicated that EGCG could be an effective substances to protect mitochondria-induced oxidative stress in cardiomyocytes of T2DM.
Moreover, epicatechin is one of the flavanols reported to attenuate DCM through modulation of mitochondrial-induced oxidative stress. Ramírez-Sánchez and colleagues
[46][118] have demonstrated that epicatechin could block the suppressive effect of high glucose on heart mitochondrial biogenesis involving mitofilin, SIRT1, PGC-1α, and TFAM levels. Treatment with epicatechin also has reversed the high level of eNOS-O-GlcNAc in the diabetic heart.
2.5. Anthocyanins
In terrestrial plants, anthocyanins are one of the most common and abundantly distributed secondary metabolites. Anthocyanins are responsible for red, purple, and blue colors in the flowers, seeds, and fruits of numerous plant species.
[47][48][119,120]. Anthocyanins are natural antioxidants because they are electron deficient, making them highly reactive to ROS
[49][121]. More than 600 anthocyanins have been extracted from a variety of plant species. They are based on the flavylium ion, which has a single fundamental core structure. As a result, the C15 skeleton is formed with a chromane ring with a second aromatic ring B in position 2 (C6-C3-C6) containing single or more sugar molecules attached at various hydroxylated sites on the basic structure. The C3 hydroxyl in the C ring commonly conjugates sugar molecules to the anthocyanidin structure
[49][121]. Anthocyanins and anthocyanin-rich foods have been found to exhibit a variety of biological functions mostly as an antioxidant that may benefit for human wellbeing
[50][122]. The role of anthocyanins has been proven to improve DCM through modulation of mitochondrial-induced oxidative stress. Anthocyanins mainly can be found in plant such as roselle, blackberries and blackcurrants
[51][123]. However, the study that was conducted by using anthocyanins in targeting mitochondrial-induced oxidative stress in DCM is still very limited.
Mitochondria damage is a key factor leading to cardiomyocytes impairment and cell death as well as other cardiac diseases and making mitochondria an attractive target for pharmacological interventions. As a matter of fact, protocatechuic acid (PCA), a primary metabolite of anthocyanins that found in roselle, has been shown to possess as an antioxidant. In an in vitro study by Semaming et al.
[50][122], PCA significantly reduced mitochondrial ROS level and attenuated mitochondrial membrane depolarization. They also found that PCA treatment attenuated mitochondrial swelling as the ROS level decrease. Not only that, PCA treatment alone was able to reduce blood glucose level via enhancing GLUT4 translocation and adiponectin secretion caused by elevated PPARɣ activity in adipocytes. This shows that mediating this mechanism is crucial in alleviating increase of FAO in the mitochondria
[52][124].
Although the use of anthocyanins on diabetic cardiomyopathy research has not yet been extensively investigated, we can presume the result of its interventions by looking at findings of its impact on different cardiac disease models. Previously, anthocyanins has proven to attenuate oxidative stress by scavenge ROS via various mechanisms including direct scavenge ROS, induction of enzymes (superoxide dismutase, catalase) responsible for ROS removal or modulation of ROS forming enzymes (NADPH oxidase) in mitochondria
[53][54][125,126]. This has been confirmed by another study reporting that anthocyanins was able to quench ROS and thus preserve mitochondrial complex activities in heart
[55][127].
2.6. Flavanones
Flavanones are another important compound found in citrus fruits, including oranges, lemons, and grapes. This group of flavonoids includes hesperidin, naringenin, and eriodyctiol. Because of their free radical-scavenging characteristics, flavanones have been linked to various health advantages
[22][96]. Citrus fruit juice and peel contain these substances, which give them a bitter taste. Citrus flavonoids have pharmacological actions that include antioxidant, anti-inflammatory, anti-hyperglycemia, and anti-hypercholestrolemia. The C ring is saturated in flavanones (saturated double bond between positions 2 and 3), giving them the alternative name of dihydroflavonols, and distinguishes them from flavones
[36][109].
Naringin, a major flavanone glycoside found mostly in citrus fruits, has been reported to alleviate mitochondrial-induce oxidative stress by preventing the high glucose-induced loss in mitochondrial membrane potential
[56][128]. In another study conducted by You and colleagues
[57][129], naringin also reduced the downregulation of mitochondrial ATP-sensitive potassium channels, which is important in sensing the metabolic changes in pancreatic beta cells and thus protecting the cardiomyocytes against the hyperglycemic condition.
Similar to anthocyanins, the extensive study on the effect of flavanone in mitochondrial-induced oxidative stress in DCM is still limited. However, in other cardiac models, flavanone has been demonstrated to modulate mitochondrial function in cardiomyocytes. Previously, flavanone was found to ameliorate mitochondrial disruption in cardiomyocytes by reducing impaired mitochondrial membrane potential and suppressing mitochondrial ROS levels and increase mitochondrial antioxidant via regulation of AMPK-mTOR signaling pathways
[58][59][130,131]. Moreover, flavanone was able to alleviate mitochondrial membrane potential collapse and preserve mitochondrial complex II activity on isolated heart mitochondria
[59][131]. Aside from that, Ca
2+ overload was reduced significantly with the treatment of flavanone and hence reviving mitochondrial function in the heart
[60][132].
Table 1 shows an overview of the role of flavonoids in alleviating mitochondrial-induced oxidative stress in DCM.
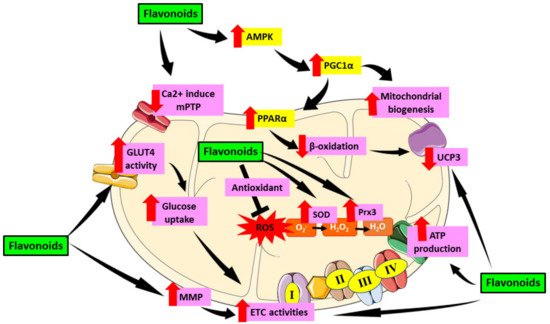
Figure 25 demonstrated the role of flavonoids in targeting mitochondrial induce oxidative stress in DCM.
Figure 25. Role of flavonoids in targeting mitochondrial induce oxidative stress in DCM. Flavonoids have been proven to alleviate mitochondrial dysfunction by targeting mechanisms involving oxidative stress in mitochondria including activation of AMPK which then activate PGC1α. PGC1α enhances mitochondrial biogenesis as well as PPARα expression and cause reduction of β-oxidation in mitochondria which then downregulate UCP3. Flavonoids also reduce the formation of Ca2+ induced mPTP which preserve mitochondrial number and prevent apoptosis. Furthermore, flavonoids were proven to enhance GLUT4 activity which then led to increase glucose uptake as well as enhance MMP that cause increase in ETC activities which further elevate ATP production. Moreover, flavonoids can act as antioxidant and scavenge ROS as well as increase SOD and Prx3 enzyme which later attenuate oxidative stress. This figure is illustrated based on the review of the previous research. Black arrow indicates flow of mechanisms; red arrow indicates increase/decrease of level/activity.
Table 1. Summary of flavonoid and its subclasses in targeting mitochondrial-induce oxidative stress in DCM.
| Flavonoid Subclass |
Type |
Study Design |
Dose |
Results |
Reference |
| Anthocyanin |
Protocatechuic acid |
In vivo; T1DM Sprague-Dawley rats |
50 and 100 mg/kg/day |
Reduce mitochondrial ROS levels, attenuated mitochondrial depolarization and decreased mitochondrial swelling in cardiomyocytes. |
[50] |
| Flavones |
Vitexin |
In vitro; H9C2 cells |
1, 3, 10, and 30 µM |
Improve mitochondrial ATP production Revive mitochondrial respiratory function by increasing expression of levels of COX IV and SDHB in H9c2 cells. |
[61] |
| Rutin |
In vivo; T2DM Wistar rats |
100 and 200 mg/kg/day |
Improve co-enzyme Q9 and Q10 in the mitochondria. |
[25] |
| Luteolin |
In vivo; T1DM Sprague-Dawley rats |
100 mg/kg/day |
Increase MnSOD and eNOS expression and decrease Ca2+ induced mPTP opening and mitochondrial inner membrane in cardiomyocytes. |
[23] |
| Isoflavones |
|
In vitro; H9C2 cells |
20–200 μg/mL |
Reduce mitochondrial-induce oxidative by lowering mitochondrial ROS generation, depolarization of ΔΨm through SIRT-1 pathway or PPAR-α which further attenuate mitochondrial dysfunction and thus conserve cardiomyocytes health. |
[31] |
| In vivo; T1DM Sprague-Dawley rats |
25, 50, and 100
mg/kg orally |
Maintained the AMPK and SIRT1 levels. |
[32] |
| Flavonol |
Icariin |
In vivo and in vitro; db/db, db/+ mice and C57 mice cardiomyocytes |
7.5, 15, and 30 µM |
Upregulate myocardium gene apelin and the cardiac mitochondrial matrix gene Sirt3.
Increase the mitochondrial membrane potential.
Reduce mitochondria ROS production. |
[41] |
| Flavonol |
Quercetin |
In vivo and in vitro; T1DM Wistar rats and H9C2 cells |
50 mg/kg and 1 and 10 μM |
Induce Prx-3 expression, causing downregulation in myocardial UCP3 protein.
Reduce cardiac Trx-2 expression and TrxR2 activity.
Induce the expression of transcription factor Nrf2/Nrf1. |
[42] |
| Quercetin and Kaempferol |
In vivo; T1DM albino rats |
200 mg/kg/twice daily |
Enhance the expression SOD1 gene, PGC 1α gene and ATpase and improve mitochondrial function. |
[43] |
| Taxifolin/dihydroquercetin |
In vivo and in vitro; T1DM C57BL/6 mice and H9C2 cells |
10, 20, and 40 µg/mL and 25, 50, and 100mg/kg/day |
Restore mitochondrial transmembrane potential in H9c2 cell lines. |
[62] |
| Dihydromyricetin |
In vivo; T1DM C57BL/6 mice |
100 mg/kg/day |
Enhance ATP levels, CS activity, and complex Ι/ΙΙ/ΙΙΙ/ΙV activities, increase ΔΨm. |
[40] |
| Flavanol |
Epigallocatechin-3-gallate |
In vivo; T2DM Goto–Kakizaki rats |
100 mg/kg/day |
Revive Complex I, III, IV, and VDAC1 activities as well as recover mtDNA copies and the mitochondrial dehydrogenase activities. |
[45] |
| |
Epicatechin |
In vivo and in vitro; T2DM C57BL/6 mice and HCAEC cells |
100 nM and 1 mg/kg/day |
Blocked the suppressive effect of high glucose on heart mitochondrial biogenesis involving mitofilin, SIRT1, PGC-1α, TFAM protein levels and reversed the high level of eNOS-O-GlcNAc of diabetic heart. |
[46] |
| Flavanone |
Naringin |
In vitro; H9C2 cells |
5 μM |
Prevent the HG-induced loss in ΔΨm. |
[56] |
| |
Naringin |
In vivo and in vitro; T1DM Sprague-Dawley rats and H9C2 cells |
80 μM and 25, 50, and 100 mg/kg/day |
Reduce the downregulation of KATP channels. |
[57] |
Summary of flavonoid and its subclasses in targeting mitochondrial-induce oxidative stress in DCM.