Arbuscular mycorrhizal (AM) fungi are biotrophic symbionts forming close relationships with an estimated 80% of terrestrial plants suitable as their host. Via an established AM fungal–host relationship, soil-bound nutrients are made available to the host plant through root cortical arbuscules as the site of exchange. At these sites, photosynthetic carbohydrates are provided to the AM fungus—carbohydrates that cannot be produced by the fungus. AM fungal–host symbiosis is very sensitive to soil disturbance, for example, agricultural tillage practices can damage and reduce AM fungal abilities to interact with a host and provide plant growth-promoting properties.
- arbuscular mycorrhizal fungi
- conservational
- glomalin
- soil quality
- sustainable
- symbiosis
- tillage
1. Introduction
Arbuscular mycorrhizal (AM) fungi are symbiotic biotrophs that form close relationships with a host plant via intracellular fungal structures, namely arbuscules, in root cortical cells [1]. It is currently estimated that AM fungi began their associations with host plants between 400 and 480 million years ago, contributing to the initial land colonisation by terrestrial plants. Approximately 80% of terrestrial plant species are in close symbiotic relations with AM fungi [2] for several plant-promoting properties, such as nutrient acquisition, increases in crop mass and yield, as well as reduced stress from abiotic pressures, e.g., soil salinity and drought [3].
AM fungi contribute to soil structure, stability and function through increased growth and mass from a host’s root system. One way in which they contribute to this is from the production of glomalin [4,5], a glycoprotein, with soil adhesive properties between microaggregates [4,6]. Additionally, branching mycelial networks produce exploratory hyphae through soil pores [7]. This exploration allows hyphae to wrap around glomalin-adhered microaggregates to form larger more stable macroaggregates, this also aids in the reduction of soil erosion by wind and water [8,9]. The management of soils, however, has the ability to preserve or damage AM fungal networks, their associations with a host plant, and their influences on soil properties [10].
Tillage, as a method of seedbed preparation, can be detrimental to AM fungal–host relations and wider mycelial network establishment [11,12]. This is indicative of conventional tillage (CT) which typically inverts soils to a maximum depth of 30 cm (20 cm within the UK [13]) forming the zone of tillage, with the formation of a densely compacted layer below (plough pan). Such a degree of soil inversion and disturbance breaks and homogenise AM fungal mycelia through the zone of tillage [13] and negatively impacts AM fungal life cycles by reducing the likelihood of AM fungi finding a host plant to form a symbiotic establishment, additionally seen in the use of excessive fertiliser use. This is a critical part of the AM fungal life cycle [14]. In contrast, zero tillage (ZT) does not invert or disturb soils. ZT practices utilise direct seed drilling. This method of land management is comparably more conservative to AM fungal abundance [9,15]. However, ZT management strategies are known for their employment of glyphosate as a means of weed removal as an alternate method to CT soil inversion [10]. Glyphosate was shown to be detrimental to AM fungi and their host crop interactions. Such disturbances to AM fungi, from both soil disturbance and chemical interaction, interfere with the AM fungal life cycle.
2. Life Cycle
The process by which AM fungi form symbiotic relationships with a host plant is a critical part of their life cycle. Figure 1 shows the process of pre-symbiosis and symbiosis between AM fungi and a host root system.
2.1. Pre-Symbiosis
The production of root hormones and organic acid exudates from the host root system begins during early seed germination and are the chemical signals deposited within the local soil environment [16]. Strigolactone, a root exudated plant hormone [17], is responsible for the recruitment of AM fungi (Figure 1a). Strigolactone stimulates AM fungal spores within the soil to begin germination, develop and grow their mycelia in the direction of the hormone-producing host plant (Figure 1b) [18]. Pre-symbiosis ends when AM fungal mycelia come into direct contact with the host’s cell surface of the root cortex and form an appressorium (Figure 1c) [19].
2.2. Symbiosis
The growth of hyphae through root structures, from the appressorium, marks the beginning of the symbiotic relationship between the host plant and fungus. To establish the relationship, intracellular arbuscules must be formed (Figure 1d), producing a large surface area for nutrient exchange from fungal acquired soil nutrients provided to the host [20]. Organelles of the plant cell must be modified in order to accommodate the arbuscule structure [21]. Hyphae begin to penetrate root cortical cells and the plant plasma membrane invaginates [22]. The plant cell-penetrating hyphae, the “trunk”, continues to branch whilst still enveloped by the plant cell membrane. After continued hyphal branching, the plant plasma membrane becomes the peri arbuscular membrane (Figure 2) and is the site of nutrient exchange [23]. The peri arbuscular membrane is continuous with the plasma membrane of the root cell; however, regulation of plant DNA has transcribed modified membrane proteins specialised for nutrient exchange [24]. Alteration of plant DNA regulation began once fungal hyphae penetrate the cortical cell [25] causing non transcribed regions of the plant chromosomes to decondense.
The mycelia of the main fungal body, branching into bulk soils will produce more spores (Figure 1e) for later germination from receiving exudated signals. Mycelia itself is able to continue growing towards plant hormone and exudate signals creating a further symbiosis with several other plants. When multi-plant symbiosis is created, it may involve several plant species rather than conforming to a single plant species. An example of this comes about from symbiosis with wheat crops and emerging weed plants. In cases of woodland soils and ectomycorrhizal fungi, multi-plant symbiosis was described as a “Wood Wide Web” [26].
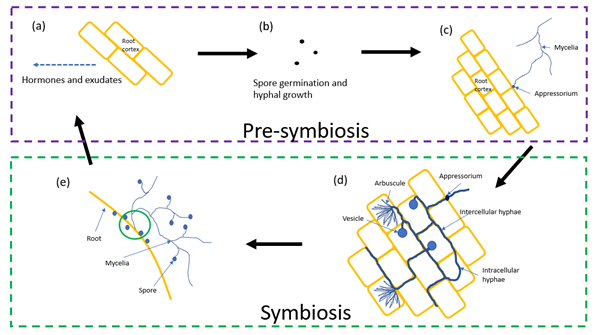
Figure 1. Diagrammatic representation, based on the model organism Triticum aestivum L., of the AM fungal life cycle through initial plant signalling (a), spore germination (b), and initial plant infection (c) in pre-symbiosis. Following infection, establishment of intracellular root cortical fungal structures (d) establishes plant–fungus symbiosis. Extra radiating mycelia are also available for growth and development from further plant exudated compounds to establish multi-plant symbiosis (e). Root arbuscules are not to scale with cortical cells.

Figure 2. ×1000 magnification of Vicker’s® microscope, from lab-controlled Zulu winter wheat growth showing AM fungal arbuscules (red), vesicle (green), peri arbuscular membrane (white arrow) and intracellular hyphae (yellow arrow). Imaged with a Bresser® HD microscope camera and stained with Sheaffer® blue [27].
3. Mycorrhizal–Host Nutrient Exchange
Through the establishment of a host–AM fungus relationship (Figure 1d), soil nutrients can be provided to the host via intracellular AM fungal arbuscules in exchange for photosynthetic carbohydrates. Current estimates suggest 20% of photosynthetically acquired hexose carbohydrates are provided to AM fungi [28] across peri arbuscular membranes.
Soil nutrient availability can alter the establishment of symbiosis. Soils deficient in nitrates produce plants with increased root exudation and encourage further AM fungal root colonisation and symbiosis [29]. The reverse is additionally true, soils high in nitrates reduce the overall AM fungal level of symbiosis. Further to this, studies were able to show that nutrient-deficient soils, producing higher levels of host–fungus interactions, result in more carbon exchange via arbuscular structures giving strong indications that a mutual relationship between plant and fungus exists through symbiosis [29,30]. However, nutrient-deficient soils may be managed in an inappropriate manner, resulting in deterioration of soils and lowering overall crop yields. An example of excessive fertiliser usage can be seen from the application of nitrogen fertilisers. The soil microbiome has a myriad of enzymatic pathways able to digest applied nitrogen-containing compounds. Nitrous oxide can be produced in large quantities, resulting in the loss of nitrogen desired for crop utilising as well as contributing to greenhouse gas emissions and algal blooms in proximity to waterways. Additionally, excessive applications of nitrogen fertilisers were shown to reduce AM fungal biomass and their degree of symbiosis. Nitrogen fertilisers are an essential part of crop maintenance and yield outputs and cannot be removed from arable management practices. However, assessing the quantity of applied nitrogen can reduce nitrogen loss to the atmosphere and increase the desired nutrient uptake via an established crop and their associated AM fungi.
3.1. Carbon
AM fungi require carbon compounds, in the form of carbohydrates, as part of their life cycle. They are, however, not able to produce the required carbohydrates alone. This brings about the need to establish close relationships with a host to provide the needed carbohydrates. Smith and Read [31] showed this can be achieved through indirect means from increases in extraradical mycelia after a root has been colonised and forming a symbiosis with several plants at any one time. Wang et al. [6] produced an in-depth review of direct carbohydrate exchange from host to fungus. The movement of carbohydrates is regulated by the plant and, in part, by the fungus. Sucrose, the photosynthetically produced carbohydrate, is transported to the root tissue via the phloem. Within these tissues, sucrose is cleaved into glucose and fructose [32] AM fungi do not prosses the ability to cleave sucrose to glucose and fructose, so must be performed via the host [6,33]. From carbon sink studies of Doidy et al. [34], it was shown that increases in root mycorrhizal colonisation cause the host plant to increase transported carbohydrates to root tissue. This increase in transported carbohydrates, however, still did not exceed 20% of the total produced photosynthetic carbon of the host. From this, it can be drawn that host biomass and carbon dioxide intake must increase to maintain the demands of both the plant and the symbiotic AM fungi.
Bonfante and Anca [24] showed changes in expressed membrane transporter proteins from transcribed regions of DNA stimulated from fungal colonisation and arbuscular formation. Sucrose transporter membrane proteins, SUT, and monosaccharide transporters (MST) were shown to increase in fungal colonised root cells. However, these transport structures were also seen to increase in the phloem, a region of root tissue that is not directly colonised by AM fungi [6]. This, therefore, reinforces the notion of plant responses to AM fungal interactions being whole plant rather than a localised phenomenon. From carbohydrate exchange in root tissues, carbohydrates must be transported from intraradical mycelia to extraradical mycelia for the continued growth and development of branching hyphae and spores in soils. To achieve this, plant-provided sugars are enzymatically converted into lipids, the main carbon storage molecule in AM fungi [35], stored within vesicles found within root tissues [27]. Bago et al. [32] found the conversion of carbohydrates to lipids, back to carbohydrates, was only 50% efficient.
3.2. Nitrogen
The supply of nitrogen in soils, without fertiliser application, is known to be a plant growth-limiting factor [36]. Many studies involving the provision of nitrogen favour methodologies of labelled N15. Johansen et al. [37] were one of many that followed labelled nitrogen from the soil, absorbed into extra radiating mycelia and incorporation into amino acids via the glutamine synthetase, glutamine oxoglyutarate aminotransferase (GS/GOGAT) cycle in the formation of arginine [6].
AM fungi are able to transfer organic nitrogen, NO3- and NH4+, to host plants and also increase soil organic nitrogen through organic matter decomposition [38]. From N15 labelled soil nitrate sources, it was shown that 21% of extra radiating mycelial sourced nitrogen was exchanged within root structures [39]. It was then later shown that between 30–50% of fungal acquired nitrogen was exchanged at root sites, with an estimated 75% reaching leaf tissue [40,41]. Govindarajulu et al. [41] were able to show the movement of arginine to intra-radiating mycelia and its breakdown into urea and ornithine. The utilisation of urea during breakdown produces ammonia gas and contributes to nitrogen loss from soils. However, most ammonia is released into the plant across arbuscular exchange sites, where ammonium transporters in AM fungi (GintAMT1, GintAMT2, and GintAMT3 in R.irregularis) are expressed in established symbiosis [42]. These transport structures are further expressed under low nitrogen conditions to facilitate further nitrogen transport, as well as AM fungal mediated acceleration of organic matter decomposition [43]. This strongly supports reductions in symbiosis from higher soil nitrogen levels. Branches of the arbuscular structure were found to express GmAMTH.1 and ATM2;3 on the peri arbuscular membrane rather than the “trunk” of the mycelia [44]. This indicates arbuscule branches are the site of nitrogen transfer.
3.3. Potassium
Potassium is the most abundant element within the soil’s composition [45], however, potassium has low availability towards plants and is a growth-limiting factor. Potassium also plays a crucial role in several plant functions, including membrane polarisation, plant growth, stomatal aperture functionality, as well as a plant’s environmental adaptation [46]. Via particle-induced X-ray emission studies [30,47], accumulation of potassium in AM fungi were seen in mycelia, vesicles and spores. Potassium transporter channels from AM fungi to the host plant are still under investigation with only four known transports; three SKC-type transport and one KT/KUP/HAK transporter [45,48]. Potassium, originating from AM fungi, was shown to increase plant stress tolerances, especially in response to salinity [49]. However, little is known regarding the mechanism of increased stress tolerance or the physiological pathway [45,50,51].
3.4. Phosphorus
Phosphorus is a major plant nutrient and is provided to a host by AM fungal branching mycelia, increasing soil uptake surface area of phosphorus, in the form of inorganic phosphate, in exchange for photosynthetic carbon across the peri arbuscular membrane (Figure 2). Phosphorus transport was shown to be induced from the establishment of symbiosis from manipulation of the Pht1 family of genes responsible for phosphate transport [52–54]. High levels of soil phosphate were shown to reduce gene transcription for strigolactone production [55] indicating reductions in symbiosis from excess soil nutrients. This was also identified from calcium spiking inhabiting pre symbiosis pathways [56]. Overall, this results in reduced translocation of other nutrients, via AM fungal mycelia, and potential reduction to soil quality [44,45,56]
3.5. Fatty Acids
Fatty acids and fatty-acid-derived lipids are crucial components of all living cells. Plant-derived fatty acids are comprised of carbon chains 16 or 18 carbon atoms long with 1 to 3 double bonds throughout the molecule [57]. These were shown to increase in AM fungi following the onset of established symbiosis [58], with 16 carbon chain fatty acids being detected and quantified, via labelled C13 methodologies [59], within intracellular arbuscules and vesicles and absent within extra-radiating mycelial and fungal spores [60]. Further studies were able to indicate the internal transport of acquired lipids via fungal hyphae and its utilisation as a carbon source for the development of further mycelia growth and spore development [60,61]. Subsequently, 95% of spore dry weight was determined to be fatty acids, along with 47% of hyphal volume in extra-radiating mycelia and 58% dry weight of intracellular vesicles in the host root cortex [62,63].
Feng et al. [59] describe lipid transfer from the host plant to AM fungus during abiotic stress and are able to show the reduction of lipid transfer under these conditions and the eventual collapse of arbuscular structures following reductions in lipid transfer resulting from the decreased regulation of lipid transfer genes for adenosine triphosphate (ATP) binding cassette (ABC) transporter stunted arbuscule (STR) and STR2. ABC, STR and STR2 are located within the peri-arbuscular membrane and are a critical component of early, and maintained, establishment of host–fungal symbiosis and have specialised functionality for the transport of host plant-derived lipids [64].
Genome and transcriptome analysis of AM fungi have revealed the absence of fatty acid synthesis genes in model AM fungi; including, Rhizophagus irregularis, Gigaspora margarita, and Gigaspora rosea [65,66]. However, AM fungi do possess enzymatic pathways to allow the elongation of plant-derived fatty acids [59]. This provides an example of the biotrophic nature of AM fungi and their dependence on a host.
3.6. Sulphur
Sulphur is an essential element required for plant growth and is becoming increasingly limited due to atmospheric sulphur rapidly reducing in concentration [67]. Soil sulphur is typically bound within soil organic matter in the form of sulphate esters and sulphonate. As with other forms of bound nutrients, sulphur is not directly available to a developing crop and requires constituents of the rhizosphere microbiome to enzymatically digest bound sulphur, making sulphur more available to the host crop [68,69]. Once plants are able to utilise sulphur, it enters several metabolic mechanisms producing metabolites such as cystine, methionine, glutathione, biotin, chlorophyll, and co-enzyme A, all of which are components of critical physiological pathways [70].
AM fungi, via their large branching mycelial network through soils and associated large surface area, were shown to take up soil sulphur and transport it to their host crop. However, studies have shown AM fungal-mediated crop growth benefits in maize (Zea mays), tomato (Solanum lycopersicum), and clover (Trifolium spp.), with other crop types still under investigation and expanding on current understanding [68,70]. Allen and Shachar-Hill [71] and Buchner et al. [72] studied Glomus intraradicies with carrot (Daucus carota subsp. sativus) root and were able to show the translocation of sulphate from AM fungi to host, concluding sulphur provisions to a host crop were limited and only occurred at the upregulation of the host. This further comes about from the AM fungal mediated upregulation of sulphur transporters within the peri arbuscular membrane in response to plant exudation as a communications pathway towards AM fungi.
Whilst AM fungal sulphur translocation provides a host crop, bacterial communities surrounding AM fungal extra-radiating mycelia contribute to the breakdown of soil-bound sulphur, making further sulphur accessible for AM fungi to absorb and transfer to a host [69,73,74]. This is an example of a tripartite relationship between AM fungi, bacteria, and crops.
3.7. Micronutrients
Micronutrients are essential elections required by any organism for growth, development, and reproduction. Examples of micronutrients in arable crops are copper, iron zinc, manganese, and cobalt [75]. Plants utilise copper and iron in redox reactive physiological mechanisms as co-enzymes and co-factors, whilst zinc has a structural support role in plant protein [76]. However, such micronutrients are also classed as heavy metals and, in high concentrations, lead to reactive oxygen species likely to damage plant cells [77]. Other metals, such as mercury, lead, and cadmium, and metalloids such as arsenic, are toxic to plants and inhibit growth, development, preproduction and crop yield [78].
AM fungi, whilst having been studied for nitrogen, phosphorus, and potassium nutrient exchange, also increase micronutrient uptake of their host [79]. In the absence of AM fungi, micronutrients are poorly mobile in soil. Schuβler and Walker [80] used compartmentalised soil pot systems to investigate the micronutrient mobility effects of extra-radiating mycelia of Funneliformis mossae, from cucumber (Cucumis sativus) roots, through soil pores. Schuβler and Walker [80] showed AM fungi contributed 75% of the total copper uptake in cucumber. Metanalysis of AM fungal copper provision has shown a significantly positive contribution to sopper host nutrition [81,82].
Zinc homeostasis is attributed to two transporter families, zinc-ion permease (ZIP) and cation diffusion facilitation (CDF). Currently, CDP has been reported within three species of AM fungi, Rhizophagus irregularis, Hebeloma cylindrosporum, and Oidiodendron maius [83]. However, AM fungal zinc acquisition from soils is dependant on soil phosphorus concentrations, as increases in zinc uptake are proportional to phosphorus acquisition [83] under low soil phosphorus. Jansa et al. [84] studied zinc and phosphorus translocation through AM fungal extra-radiating mycelia of R. irregularis from zinc-65 and phosphorus-33 isotopes, showing AM fungi can translocate zinc and phosphorus through 14 cm of mycelia networks to a host plant. However, increases in soil zinc reduce AM fungal mediated zinc acquisition whilst having no influence on crop zinc contents. In contrast, low soil zinc increased AM fungal zinc acquisition. This is an example of AM fungal–host communication and regulation of nutrient requirements.
4. Arbuscular Mycorrhizal Fungi and Soil Structure
AM fungal mycelia extend into bulk soils, primarily in the search of nutrient sources to acquire for plant growth in exchange for photosynthetic carbohydrates [32]. However, soil structure benefits from the extensions of mycelia from the formation of more stabilised soil macroaggregates from microaggregates formed from the presence of glomalin along the length of the fungal mycelia as an addition to structural support [85]. Through the improvement to soil stability, and reduction in wind and water erosion coming about from this, soil quality is also improved. A plot experiment by Li et al. [86] further explored this from soil inoculants of AM fungi and found increased aggregate stability and maintenance of a neutral soil pH. This is an example of how soil inoculants of AM fungi improve soil physical attributes, aggregation, through biological means, is key cooperation that defines soil quality [86,87]. The presence of glomalin is additionally a key component of AM fungal contribution to the improvement of soil quality.
4.1. Soil Glomalin
AM fungi are associated with the production of the soil glycoprotein glomalin [1,5,6,85,86–93]. Of the 250 species of known AM fungi, an estimated 90% of all species are capable of producing glomalin [94]. Glomalin produces improvements to soil microaggregates and leads to reductions in soil erosion [95]. Improvements to microaggregates can also be seen from the presence of fungal mycelia that physically bind soil aggregates together. Glomalin has adhesive properties between soil aggregates and particulates, whilst being comprised of an estimated 30–40% carbon, 3–5% nitrogen, 4–6% hydrogen, 30–50% oxygen, 0.03–0.1% phosphorus and 0.8–8.8% iron [96]. Following this, Glomalin Related Soil Protein (GRSP) comprises a large proportion of soil organic matter (SOC) [88], which is very stable [1]. GRSP is determined via the Bradford assay and is also referred to as Bradford reactive soil proteins (BRSP) [15,58,63]. Whilst this method is advantageous for large-scale field investigations into soil glomalin at a lower cost, antibody assays are able to more reliably determine the presence and abundance of soil glomalin. The drawback to an antibody assay is the cost and greater degree of required skill set to perform the assay. For large-scale field studies, a glomalin antibody assay is not always feasible and the Bradford assay is employed. It should be noted, however, the presence of humic acid in the extractant of the Bradford assay was shown to interfere with photospectrometry quantification of glomalin and can lead to results with reduced reliability, emphasising the need for a greater degree of sample replicates [4,85,89]. Some studies have estimated that glomalin contributes 5% of the total SOC measured within a soil sample [88], with a range of longevity in soils between 7 and 42 years [4]. Glomalin’s ability to remain in soils for several decades comes about from the water-insoluble characteristics and resistance to heat degradation from structural similarities to heat shock protein [5]. Further studies have additionally been able to show that increases in atmospheric carbon dioxide can produce increases in soil glomalin concentrations [6,51]. This is thought to occur from the photosynthetically fixed carbon, in the form of carbohydrates, being transported to root systems and provided to the present symbiotic fungi [97]. From the provided plant acquired carbohydrates, AM fungi utilise this as a source of carbon and produce glomalin [90,97]. Glomalin has the added function of structural support for fungal mycelia [85], but can be damaged and broken by the application of land management practices such as CT. This had negative connotations for soil quality from the reductions of soil aggregation leading to increased soil erosion from water and wind.
5. Tillage
Through the application of invasive conventional land management practices, such as CT, there is a building body of evidence to suggest the inability of AM fungi to survive the homogenisation and aeration of a soil profile [98]. Kabir [99] identified that the employment of intensive tillage practices is a major reducing factor of AM fungal abundance and diversity, with a later warning from Sosa-Hermandez et al. [98] to move away from such intensive practices. Sale et al. [100] investigated the diversity and abundance of AM fungi via soil spore analysis to a depth of 40 cm from reduced tillage (RT) and CT systems. Their findings supported the work of Kabir [99] showing a reduction in abundance and diversity from the application of an invasive soil management regime. However, Sale et al. [100] also were able to show that the diversity of abundance of AM fungi was greater in soils deeper than 40 cm. Within the UK, CT typically inverts soil to a maximum depth of 20 cm [13] but can invert soils to a maximum depth of 30 cm with a plough pan found between 30–40 cm deep. Findings of Kabir [99] and Sale et al. [100] support the notion of CT having negative implications for the damage and breaking of a soil AM fungal community, reducing the symbiotic nature of AM fungi. A potential solution for this is to manage soils through a zero tillage (ZT) practice, a practice that removed the mycelial damaging soil inversion. Compared with soil inversions of CT, ZT has very little to no soil disturbance. A possible drawback of a ZT system is an increase in soil bulk density, use of agrochemicals for weed control and the reduced mobility of nutrients and fertilisers through a soil profile [98].
Table 1 produces a comparison between tillage types and their impacts on soil. From the management of soils, resultant of CT, soil aggregation is reduced and leads to increases in soil erosion by means of wind and water [101]. Whereas in ZT soils, reduction of erosion and aggregation is seen. From improved aggregation, the decomposition of crop residue is protected [102], whilst having an additional benefit of improving soil aggregation [103]. Crittenden et al. [104] investigated the importance of tillage management on the stability of soil aggregates and found soil organic carbon (SOC) serves to increase aggregation, whilst Sheehy et al. [105] were able to show SOC has greater aggregate stabilising properties for macroaggregates and should be used as an indicator for carbon loss resultant of tillage management practices. Through a series of longer-term experiments, improvements of ZT were seen to increase microaggregates and improvements to total SOC levels [106]. This leads to reductions in soil erosion from wind and water, with reductions from carbon dioxide emissions additionally seen.
Table 1. Summary of select tillage types and their associated mechanical impact characteristics upon soils and arbuscular mycorrhizal (AM) fungi [107–112].
|
Tillage Type |
Equipment Employed |
Tillage Characteristics |
Impact on Soil and Arbuscular Mycorrhizal Fungi |
|
Conventional |
Mouldboard plough |
· Maximum 30 cm soil inversion · Loosens and aerates soil · Buries crop residue from previous crop |
· Reduces AM fungal abundance and diversity · Increase in soil erosion · Reduce soil carbon and increase carbon dioxide emissions · Increases in soil aeration · Reduction of soil bulk density in zone of tillage · Reduces weeds · Reduces select fungal pathogens |
|
Reduced |
Rotary disc |
· Maximum 15 cm soil disturbance |
· Reduced soil erosion compared to conventional tillage · Not always effective against weed reduction |
|
Strip |
Specialist equipment · Disk opener · Coulter |
· Maximum 15 cm soil disturbance · Only soils to be drilled are disturbed · Soil drying effect of conventional tillage |
· Weed reduction can be difficult · More passes of heavy equipment may be required · Removal of a cover crop may be difficult · Aerates top soil · Soil carbon, nitrogen and phosphorus are lower than zero tillage |
|
Zero |
Direct seed drill |
· Direct seed drilling into top soil · Little to no soil disturbance · Not suitable for soils with poor water drainage · Crop residue remains on surface of soil |
· Increases AM fungal abundance and diversity · Decreases in soil erosion · Increase soil carbon · Increase in soil bulk density · Increase in select fungal pathogens · Agrochemicals required to reduce weeds |
5.1. Tillage and AM Fungi
AM fungal mycelia act as the primary inoculum of host plant roots [113]. Applying tillage to soils breaks the delicate mycelial networks [114]. Applying CT levels of soil inversion homogenises a maximum depth of 30 cm, 20 cm typical in the UK, dramatically diluting broken mycelia across depths that AM fungi may struggle to survive due to reduced oxygen content of soils towards anaerobic conditions [115]. The dilution of mycelia causes delays in root colonisation [116] as the network of hyphae needs to redevelop. Additionally, reduced levels of resources will be available to the fragments of mycelia and can result in the death of many mycelial fragments further reducing the amount of AM fungi in CT soils [117]. Furthermore, this has negative detrimental implications for soil aggregation. AM fungi can be found in soils low in oxygen, however, are found in symbiosis with a host plant and able to utilise plant-derived resources [118].
ZT practices do not damage the mycelial networks in top soils (<10 cm) allowing them to remain intact and able to begin root colonisation for host–fungal symbiosis [6]. Studies by Galvez et al. [119] and Sharma-Poudyal et al. [120] are able to show agricultural land management practices of a more conservation orientation, away from CT, have positive growth implications for soil fungi in both abundance and diversity. Sharma-Poudyal et al. [120], via molecular identification methods, specifically names Humicola, Cryptococcus, Cadophora and Hydondontaceae as being more abundant in ZT practices compared to CT sampled soils. Whilst these are part of a wider soil microbiome, they are not however classified as AM fungi.
Through the work of many others, the reduction of a diverse soil fungal community was reported [24,85,121]. This focuses primarily on fungal biomass within soils. An example of a fungal biomass marker, used for biomass estimations, comes about from ergosterol. Ergosterol acts as a cell membrane stabilising and support molecule in a similar way to the functions of cholesterol in mammalian cell membranes [122]. Biosynthesis of ergosterol has high demands on AM fungal metabolism but is offset by ergosterol’s ability to enable the fungus to withstand climatic conditions and variances [123].
5.2. Tillage, Glomalin and Soil Erosion
An increasing body of research exists for the effects of tillage on WSA via tillage implications to glomalin. With increases in soil glomalin, soil aeration and drainage are improved along with stable aggregates. Wilson et al. [124] noted high correlations between soil aggregation and carbon sequestration with increases in AM fungal abundance. Lovelock et al. [88] produced estimates of between 3% and 5% of soil carbon and nitrogen, respectively, being attributed to glomalin. Hontoria et al. [125] studied differences between CT and ZT systems in respect to WSA and extractable, Bradford reactive, glomalin. Hontoria et al. [125] were able to show ZT had nearly a third higher stabilised soil aggregates and three to five times more extractable glomalin.
Sharifi et al. [96] produce experimental rationale that glomalin had a greater influence on improved soil aggregation than organic matter contents. Additionally, Shafiri et al. [68] were able to show a positive correlation between glomalin and WSA. This was also shown earlier by Curaqueo et al. [126]. Whilst tillage does influence the amount of glomalin within soils, the plant species to which host AM fungi alter the quantity of glomalin deposited in soils. Therefore, crop type can improve or reduce initial glomalin levels, and relate to changes in WSA. However, the application of tillage can reduce glomalin through microbial metabolism. This not only reduces soil carbon but reduces aggregate adhesion leading to increases in soil erosion.
Nautiyal et al. [127] studied the influence of glomalin in regards to its contribution to soil carbon stores and was able to show a significant positive relationship with organic carbon contents of soils (p < 0.001) and aggregate stability. This conforms to the “glue” description of glomalin, by Wright and Upadhyaya [4], acting on soil aggregates aiding in the formation of macroaggregates from microaggregates. Whilst the reported findings of Nautiyal et al. [127] are important, Nautiyal et al. [127] are unable to consider the effects of tillage and land management practices on the quantities of glomalin in soils and alterations to glomalin’s adhesive properties. However, Sheehy et al. [105] produces direct comparisons between soil aggregates of three tillage practices (conventional, reduced and zero-till) and is able to give clear indications that zero-till practices maintain a greater proportion of macroaggregates, contribution to more stable soils and erosion resistant, than soil aggregates in conventional till. Sheehy et al. [105] additionally commented on the percentages of soil organic matter between the tillage practices and noted zero-till to have greater soil carbon when compared to conventional and reduced till with conventional till having the lowest quantifiable soil carbon. Ample literature is able to comment and explore the effects of tillage on soil aggregation or soil carbon stores between differing types of applied tillage, however, very few studies were able to comment and produce rationale regarding the direct influence of tillage against glomalin and how such an influence relates to soil aggregation with potential suggestions for which tillage practice produces the greatest benefits to overall soil quality. Shariffi et al. [96] explored the relationship between glomalin and tillage or fire land management in the Zagros forests of Iran. The tillage depth of Shariffi et al. [96] is equivalent to CT in European countries. However, Sariffi et al. [96] were unable to comment on the effects of such agricultural practices on soil aggregates but were able to conclude that applying tillage to soils did reduce the amount of quantifiable glomalin with it the system. Wright et al. [128] produced one of the few studies that make attempts at comparing tillage, glomalin and aggregate stability of soils. The results of Wright et al. [128] indicate ZT was constantly supported increased quantities of glomalin and soul aggregates compared with CT-treated soils. However, the presented data does not give clear indications that there is a largely significant difference between ZT and CT treatments through the data transformation of glomalin to total soil carbon ratios. This is largely contradictory to much of the other literature that has indicated greater differences between NT and CT soil aggregates and glomalin. An example of such is the study by Hontoria et al. [125]. The study area from Hontoria et al. [125] utilises olive groves in central Spain and is difficult to directly compare with the majority of literature that has focused upon arable cereal cultivation. Hontoria et al. [25], whilst concluding glomalin had a positive influence on soil aggregation, were not able to produce data with statistical support to justify such a conclusion, leaving the connection between glomalin, tillage and soil aggregates still under question and requiring further investigation
6. AM Fungi and Agrochemicals
Agrochemicals, such as herbicides, pesticides and fungicides, were used in arable agriculture for several decades with increasing intensity in aims to increase crop yields and productivity [11], as well as soil nutrients to sustain the increasing demand on food production [129]. The literature describes the influence of agrochemicals and fertilisers on human health and other mammals, birds and fish [129,130]. However, the soil microbiome is typically not explored and considered to the same degree in respect to agrochemical applications. Agrochemicals have specific effects upon their desired targets; for example, glyphosate herbicide reduces weed populations in ZT arable systems rather than inverted soils accomplishing a similar effect in CT management [131]. Glyphosate, however, additionally has unforeseen consequences upon the soil microbiome as some constituent species within the microbiome are non-target organisms (NTO) [132].
Glyphosate (phosphomethyl glycine) inhibits the enzyme 5-endopyruvylshikimate-3-phosphate (EPSP) synthase within growing weeds [11,133,134]. EPSP synthase is a component of the Shikimate pathway in the synthesis of tryptophan, phenylalanime, and tyrosine amino acids [11,134]. Tryptophan and phenylalanine amino acids are crucial components in the production of auxins and flavonoids, while tyrosine metabolites are critical for plant survival [133]. The Shikimate pathway and EPSP synthase are also found within some bacterial and fungal species as well as targeted plant species [135,136]. Glyphosate is typically short-lived in soils, approximately 24 days (Figure 3) [130] from adsorption onto soil particles [137], this increases the duration of glyphosate persistence in soil extending the period when the impact on NTO may be realised [138]. Aminomethylphosphonic acid (AMPA), a metabolite of glyphosate, persists within the environment for longer, shown in Figure 3, and has superior mobility within soils [130]. Giesy et al. [139], in an extension of their study of glyphosate ecotoxicity and NTOs to AMPA, found that AMPA is equally as toxic to NTOs as glyphosate. Evidently, the greater persistence, mobility and comparable ecotoxicity of AMPA is of greater concern to bacteria and fungi that share the Shikimate pathway and EPSP synthase [139,140], both of which are targeted by glyphosate when applied as a mode of weed control.
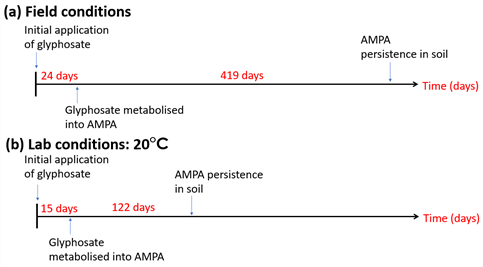
Figure 3. Timeline of glyphosate metabolism and Aminomethylphosphonic acid (AMPA) persistence in soils from both field soils (a) and laboratory-controlled soils (b), constructed from data provided by the Pesticide Properties DataBase (PPDB) [15,130].
The effect of glyphosate on AM fungi has significant implications toward the maintenance of a functional soil microbiome within the plant rhizosphere but literature on the topic remains sparse. The study by Wilkes et al. [15] determined that, under controlled conditions, glyphosate was able to inhibit AM fungal–host established relationships and reduce root cortical arbuscular structures by a factor of 41.6, equating to a reduction of 5057.14%. This supports in part the conclusions of Zaller et al. [12] who also found that the application of glyphosate reduced AM fungal colonisation and symbiosis. Furthermore, Wilkes et al. [15] were also able to quantify the overall growth implications towards AM fungi in soils supporting winter wheat between soils treated with glyphosate and soils receiving no chemical treatment. Results demonstrated that fungal biomass, as quantified via ergosterol, was inhibited at glyphosate concentrations above 75 gL−1 for 12 days. However, fungal biomass began to increase after 12 days post glyphosate application at concentrations of 75 gL−1, suggesting AM fungal growth recovers after glyphosate begins to break down. Growth inhibition and potential growth recovery were seen in both CT and ZT soils under controlled growth conditions reinforcing the direct effects of glyphosate upon AM fungal growth. Host wheat root cortical arbuscules were additionally seen to be inhibited at concentrations over 75 gL−1, beginning to increase marginally after 12 days at concentrations below 75 gL−1. This demonstrates the disruption to the AM fungal life cycle resultant of glyphosate [14,15,141].
In a similar case to glyphosate, other agrochemicals have NTO properties towards AM fungi. Whilst NTO influences of glyphosate will not be of direct relevance to all land managers, as the employment of glyphosate is specific to zero-till practices, influences of fungicides and pesticides are ubiquitous across many types of arable management. The method of agrochemical application, such as a fungicide seed treatment or direct application to a developing crop, was described by the literature to result in greater inhibitory effects to AM fungal–host colonisation as well as extra radial hyphae and spore production [142–144]. Select fungicides were studied to have neutral effects regarding AM fungal–host colonisation and development into bulk soils through hyphal growth. For example, Agrox™ and Thiram™ (Table 2) fungicides applied as a seed treatment were not seen to reduce AM fungal–host colonisation, sporulation, hyphal development, or glomalin soil concentrations in chickpeas (Cicer arietinum) [144].
Burrows and Ahmed [145] were able to show metalaxyl fungicides applied to maize (Zae mays L.) had varied influences on symbiotic AM fungal populations. Burrows and Ahmed [145] were able to show AM fungal–maize colonisation increased in the presence of metalaxyl alone and in combination with febuconazole. This is in contrast to the inhibitory effects of metalaxyl influences on AM fungal abundance and host colonisation of leeks (Allium ampeloprasum var. porrum), emphasising potential plant-derived AM fungal protection from metalaxyl.
Table 2. The effects of fungicide active components on arbsuclar mycorrhizal (AM) fungal abundance in soil and the rhizosphere, the influence on sporulation, and the concentration of soil glomalin between three different methods of fungicide application and the crop to which applied [130,143,144,146,147].
|
Method of Application |
Product |
Active Component |
Crop Type |
Effect on AM Fungal Abundance |
Effect on Sporulation |
Effect on Soil Glomalin Concentration |
|
Seed treatment |
Agrox™ |
Captan |
Pea (Pisum sativum), Chickpea (Cicer arietinum) |
Neutral |
No change |
No change |
|
Allegiance™ |
Metalaxyl |
Pea, Chickpea |
Negative |
No change |
No change |
|
|
Apron Maxx RTA™ |
Fludioxonil and metalaxyl |
Pea, Chickpea |
Negative |
No change |
No change |
|
|
Trilex AL™ |
Trifloxystrobin and metalaxyl |
Pea, Chickpea |
Negative |
No change |
No change |
|
|
Vitaflo 280™ |
Carbathiin and thiram |
Pea, Chickpea |
Negative |
Inhibited |
Reduced |
|
|
Crown™ |
Carbathiin and thiabendazole |
Pea, Chickpea |
Negative |
Inhibited |
Reduced |
|
|
Thiram 75wp™ |
Thiram |
Pea, Chickpea |
Neutral |
No change |
No change |
|
|
Plant application |
Benomyl |
1-[(butyamino)carboyl-1H-benzimidazole-2yl] carbonate |
Proso millet (Panicum miliaceum) |
Negative |
No change |
No change |
|
Bavistin |
Methylbenzimidazol-2-yl carbonate |
Proso millet |
Negative |
No change |
No change |
|
|
Agrox™ |
Captan |
Proso millet |
Positive |
No change |
No change |
|
|
Mancozeb |
Manganese ethylenebis (dithiocarbomate) (polymatrix) complex zinc salt |
Proso millet |
Negative |
No change |
No change |
|
|
Soil drench |
Benomyl |
1-[(butyamino)carboyl-1H-benzimidazole-2yl] carbonate |
Cucumber (Cucumis sativus) |
Negative |
Inhibited |
Reduced |
|
Fenpropimorph |
Rac-(2R,6S)-4-[(2E)-3-(-4-tert-butylphenyl)-2-methylpropyl]-2,6-dimethylmorpholine |
Cucumber |
Negative |
Inhibited |
Reduced |
|
|
Propiconazole |
1-((2-(2,4-dichlorophenyl)-4-propyl-1,3-dioxolan-20yl)methyl)-1H-1,2,4-triazole |
Cucumber |
Negative |
Inhibited |
Reduced |
|
|
Propiconazole and fenpropimorph |
As above |
Cucumber |
Negative |
Inhibited |
Reduced |
Fungicide type and method of application have a profound effect on AM fungal abundance, sporulation, and host colonisation [148]. Table 2 presents several fungicides, their active component, along with their influence on AM fungi and their host crop. Soil drench applications of fungicides, direct application of fungicide to soils, were studied by Kjoller and Rosendahl [147] and demonstrated the reduced abundance of AM fungi in bulk soils and rhizosphere soils from ITS sequencing. Benomyl was used in both the studies by Kjoller and Rosendahl [147] and Channabasava and Jorquera [146]. However, Channabasava and Jorquera [146] applied benomyl fungicide to a developing crop and were able to show a reduction to AM fungal biomass but to a lesser degree than the soil drench application utilised by Kjoller and Rosendahl [147]. This is due to the quantity and duration of exposure of benomyl to soil-dwelling AM fungi. A further difference between the two studies, which will have a large implication towards fungicide application methods, is the difference in studied crop: cucumber (Cucumis sativus) and proso millet (Panicum miliaceum). Both of these crops are from different plant families and will interact with AM fungi in marginally different mechanisms. Therefore, the interaction and influences of fungicides towards AM fungi are not clear cut and are also dependant on the host crop.
Interestingly, the crop application of Agrox™ was quantified to increase AM fungal abundance in bulk soils and the rhizosphere, from ITS sequencing methodologies, however, had no influence on sporulation and soil glomalin concentrations [144,146]. Channabasava and Jorquera [147] did not investigate the degree of established symbiosis with host crop root cortical cells and were not able to comment whether the quantified increase to AM fungi from Agrox™ also increased symbiotic structures, i.e., arbuscules, or was resultant of an increase in intra-radiating hyphal networks. This is an effect on AM fungi that requires further validation.
AM fungi are of agronomic importance, however, are impacted by the application of pesticides, with previous studies showing a range of influences resultant of pesticide application. Pesticides with a short half-life (7 to 21 days) were shown to have a reduced impact on AM fungal–host colonisation, abundance, and sporulation due to reduced interaction with AM fungi as an NTO [149]. Ipsilants et al. [150] comment on the low soil persistence of azadirachtin, a pesticide from neem trees (Azadirachta indica Juss). The half-life of azadirachtin was reported to be within 14 days [151]. This is primarily due to microbial degradation of azadirachtin. However, azadirachtin has been shown to have fungicidal properties, with studies showing increased application concentrations up to ten-fold have bacterial and wider fungal genera inhibition properties [152].
Wang et al. [153] produced a study investigating phoxim pesticide with application concentrations ranging from 0 to 400 mgL−1 and applied directly to soils supporting the development of carrots (Daucus carota) and spring onions (Allium wakegi). Wang et al. [153] were able to show all phoxim applications reduced AM fungal colonisation of carrots but had no impact on AM fungal colonisation of spring onions. In a similar case to applications of fungicides, the host crop plays an invaluable role in the mitigation of agrochemical effects on the preservation of AM fungi communities. Furthermore, application time throughout the cropping year is also highly variable. Ipsilants et al. [150] applied a range of pesticides: azadirachtin, carbendazim, pyrethrum, spinosad, and terpens, across a range of days (20 to 90 days) post planting for a pepper (Piper nigrum) crop. Chemical applications at 20 days had the greatest degree of influence on AM fungi-mediated changes to root length. Spinosad and pyrethrum increased AM fungal colonisation and reduced root length by an additional 7% compared with control samples receiving no pesticide treatment. Such an increase in AM fungi colonisation may be the result of reduced community competition allowing Glomus mosseae, as identified by Ipsilants et al. [150], to increase host colonisation. All pesticide applications at day 90 produced increases in the degree of G. mosseae root colonisation, further suggesting reduced community competition within the rhizosphere has allowed G. mossaea to increase abundance and host interactions. Several studies have alluded to the reduced community of fungivorous nematodes from pesticide applications. Fungivorous nematodes were also studied to reduce the overall abundance of AM fungi. Pesticide-mediated reduction of fungivorous nematodes may be the causation for the increase in G. mossaea reported by Ipsilants et al. [150].
7. Abiotic Management of AM Fungi
Over the past several decades, literature has described the fragile nature of AM fungi along with the degrees of destruction and inhibition resultant of agricultural practices, including soil disturbance and chemical applications [130,154,155]. However, the physical and chemical properties of arable soils can limit the initial population of AM fungi. Such properties include soil salinity, drought, and heat stress.
Soil salinity and salt-stressed crops are rapidly expanding issues facing modern agriculture. Sodium chloride is the most dominant form of salt found to increase soil salinity and derived from primary sources such as weathering of parent rock, as well as from sea water spray carried further in-land and deposited over arable soils [155]. Secondary processes that increase soil salinity can be seen from poor irrigation and drainage, improper management of water, and groundwater. The Food and Agriculture Organisation (FAO) [156] estimated a total area of 1 billion hectares, across 100 countries, were suffering from saline soils, with 0.3 to 1.5 million hectares added to that estimates every year reducing the overall area for adequate crop production. Several mechanical methodologies could be employed to reduce soil salinity, however, the application of AM fungi as a means of bio-amelioration [155]. Studies have shown the increased maintenance of ionic homeostasis, osmotic equilibrium, induce antioxidant synthesis, enhance photosynthetic efficiency, and regulate phytohormone production to mitigate the influence of soil salt growth inhibition and nutrient acquisition leading to reduced crop yields [157,158]. Some sources advise the addition of sulfuric acid to soils to reduce salinity along with gypsum salts or increased irrigation [159]. However, these will have connotations for AM fungi with the potential to reduce their interaction with a host crop to a greater degree than the salinity of the soil alone. Several studies have begun to produce a series of AM fungal species-specific applications to a developing crop (Table 3) to biologically mitigate the abiotic stressor [154,160–168]. Santander et al. [169] investigated the use of AM fungi as a biofertiliser for the mitigation of salt stress and yield increase in lettuce (Lactuca sativa), finding Claroideoglomus claroideum inoculations increased crop biomass and nitrogen uptake. C. claroideum was concluded to have mitigated salt stress from high salinity soils. However, inoculating L. sativa with AM fungal spores isolated from saline soils had a significantly reduced effect on overall crop biomass [169], potentially indicating a change in AM fungal diversity.
Table 3. Arbuscular mycorrhizal (AM) inoculum to established crop to mitigate abiotic plant stressors (salinity, heat and drought), with measured resultant effects. An update to [154] [157–168].
|
Plant Stressor |
Crop |
AM Fungal Inoculum |
Crop response |
|
Salinity |
Cucmis sativus L. |
Glomus intraradices, Glomus mossaea |
Increased chlorophyll content in leaves and overall biomass |
|
Solanum lycopersicum L. |
Glomus intraradices |
Increased ion absorption and leaf chlorophyll |
|
|
Leymus chinensis |
Glomus mosseae |
Increased AM fungal colonisation, water content, and phosphorus and nitrogen uptake |
|
|
Triticum aestivum L. |
Rhizophagus intraradices |
Maintianed overall biomass, increased water uptake |
|
|
Heat |
Triticum aestivum L. |
Rhizophagus irregularis Rhizophagus intraradices |
Increased nutrient content and uptake, increase to overall biomass and water content |
|
Zea mays L. |
Rhizophagus intradices |
Increased crop biomass and leaf chlorophyll |
|
|
Drought |
Triticum aestivum L. |
Glomus mosseae Glomus fasciculatum Rhizophagus irregularis Rhizophagus intraradices |
Increased crop biomass, ascorbic acid content, and leaf chlorophyll |
|
Triticum aestivum |
Glomus masseae |
Increased crop biomass, ascorbic acid content, nitrogen and phosphorus metabolism, and leaf chlorophyll |
|
|
Triticum durum |
Rhizophagus intraradices |
Increased metal ions (copper, zinc, manganese) |
|
|
Zea mays |
Rhizophagus intraradices |
Increased absorption of phosphorus, potassium, nitrogen and magnesium |
Global climate change has driven an increased persistence in drought across many regions of the world. Drought drastically suppresses plant growth and reduces overall crop yields [169]. Many soil microbes, AM fungi, in particular, aid plant responses to drought. However, the interactions between plant and microbe in response to drought conditions, are not fully understood and are still an area requiring further investigation. Literature has demonstrated the increased water uptake from AM fungal branching mycelial networks through soil pores, not typically explored by host root systems, and the transport of water to the AM fungal host [159,170–172]. Duc et al. [170] and Auge et al. [158] reported the increased stomatal conducting leading to a higher transpiration rate, drawing up more AM fungal acquired water through the plant and mitigating drought conditions further for the host crop.
Studies and arable advice to reduce drought stress aim at increasing soil organic matter and moving soil management regimes towards conservational practices such as reducing till and ZT [155]. As shown by Wilkes et al. [173], the implementation of ZT practices conserves AM fungal populations and increases their influence on soil quality. Kozjek et al. [174] investigated the influence of drought on winter wheat via the construction of drought shelters over selected areas of developing crops and effectively reducing rainfall irrigation by 65%. Via molecular sequencing methodologies, Kozjek et al. [174] were able to show the predominate AM fungal genera, Acaulospora, Paraglomus and Funneliformis, associated with winter wheat both under drought shelters and in open field conditions. Furthermore, Kozjek et al. [174] demonstrated the adaptability of established AM fungal symbiosis under temporary drought between organic and CT farming systems. Reports of the adaptability of AM fungi in CT-managed soils are advantageous as CT-managed soils have the greatest degree of negative impacts and implications to AM fungal populations, leading to reductions in soil quality [173].
One of the larger issues facing the implementation of field scale inoculations with AM fungi under drought conditions is producing the required inoculum. AM fungi are biotrophic organisms and cannot be cultured under laboratory conditions to the required quantities for field applications over large areas. Due to this, several studies have attempted to increase AM fungal abundance via indirect means. One of these is through the soil inoculation of mycorrhizal helper bacteria (MHB) [175]. MHB can have multiple interactions and form a series of interactions with both a host crop and present AM fungal populations, increasing both AM fungi abundance and acting as a plant growth-promoting rhizobacterium (PGPR) [173]. MHB are readily culturable under laboratory conditions and studies have shown the feasibility of field scale inoculations with MHB. Under drought stress conditions, inoculating soils with MHB may be more advantageous [176,177]. However, it is worth noting that, such as interactions with a host crop, AM fungal species should be identified before MHB are applied to soils as a species-specific interaction was shown in the literature [154,173,175,177].
Plants often face multiple stressors simultaneously rather than individually. Global temperatures are currently increasing in all regions, with profound drawbacks on the production of select crops [163,178,179]. An increase of 10–15°C is adequate to endure heat stress, a plant stressor that was reported with increasing frequency in the last decade and attributed to global warming [170,180]. AM fungi were studied to aid in the mitigation of heat stress in a similar manner to that of increasing drought tolerance in crops, i.e., through the provision of water, as well as regulating root hydraulic pressure, aquaporin gene expression, and phytohormone exudation [181]. Duc et al. [170] highlight the need for continued investigations into AM fungal mitigations of heat stress as the body of literature addressing such a problem is greatly limited, however, is going to become a more pressing issue with further increases to global temperatures as global warming continues.
8. Summary
In summary, estimates were able to identify that AM fungi have been historically monumental in the development of plant evolution and are a critically important symbiotic organism for arable agriculture with crop growth promotion, soil stability and soil quality increases.
AM fungi can ascertain, utilise, and transport soil-bound nutrients to a host crop. This is of great advantage to a developing crop as the plant can use its resources for continued growth and development of above-ground biomass rather than exploratory root systems. However, the application of arable fertilisers can disrupt the degree of established symbiosis between AM fungi and host. Through the application of inappropriate quantities of fertiliser, AM fungal abundance, biomass, and symbiosis are negatively influenced. This has profound connotations for the reduction in soil quality and support of an arable crop. This suggests further work is required to understand and tailor fertiliser applications for the desired crop and the soils in which support them.
The manner in which arable soil is managed has a profound influence on the abundance and symbiosis of AM fungi. Soil managed via CT is the most invasive and AM fungal disruptive, whilst ZT managed soil as a stark contrast to CT management, is the least soil invasive and AM fungal disruptive, leading to increased AM fungal contributions to soil stability and soil quality. However, agrochemicals, such as glyphosate employed in ZT management strategies, additionally is detrimental to AM fungal communities and their associations with a host crop. Regardless, sustainable, conservative soil management utilising glyphosate maintains a greater abundance of AM fungi within arable soils when compared with disruptive CT management.
The use of other agrochemicals, such as fungicides and pesticides, have a varied effect on AM fungi populations within the rhizosphere. Many of these chemical applications have reducing and/or inhibitory properties towards AM fungal–host colonisation. However, several agrochemicals have neutral effects towards AM fungi, not influencing the degree of crop root-associated AM fungi, maintaining nutrient transfer and soil quality increasing effects from glomalin. A select number of fungicides and pesticides were shown to increase the abundance and symbiotic root structures of AM fungi. Current understanding potentially suggests a reduction in community competition within the rhizosphere allowing AM fungi to increase in abundance and become more dominant within this region of soil.
Soil properties also cannot be ignored in respect to arable management and crop production, such that drought susceptible regions require a modified soil management strategy that could include a focused application of select AM fungal species dependant on the crop under development. This is also true for many abiotic plant stressors. In order to overcome this, further work on species-specific interactions between AM fungi and host crops in a variety of soil textures, agrochemical types and applications, as well as potential plant stressors, should be studied further. Such a tailored approach is a monumental undertaking, however, the benefits of such information and modifications to arable practices could lead to increased soil quality and crop production in soils and environments that may otherwise struggle to support a crop.