Parkinson’s disease (PD) is a slowly progressive multisystem disorder affecting dopaminergic neurons of the substantia nigra pars compacta (SNpc), which is characterized by a decrease of dopamine (DA) in their striatal terminals. A crucial unmet demand in the management of Parkinson’s disease is the discovery of new approaches that could slow down, stop, or reverse the process of neurodegeneration. Novel potential treatments involving natural substances with neuroprotective activities are being developed. Curcumin is a polyphenolic compound isolated from the rhizomes of Curcuma longa (turmeric), and is considered a promising therapeutic and nutraceutical agent for the treatment of PD. However, molecular and cellular mechanisms that mediate the pharmacological actions of curcumin remain largely unknown. Stimulation of nicotinic receptors and, more precisely, selective α7 nicotinic acetylcholine receptors (α7-nAChR), have been found to play a major modulatory role in the immune system via the “cholinergic anti-inflammatory pathway”. Recently, α7-nAChR has been proposed to be a potential therapeutic approach in PD. In this review, the detailed mechanisms of the neuroprotective activities of curcumin as a potential therapeutic agent to help Parkinson’s patients are being discussed and elaborated on in detail.
- curcumin
- Parkinson’s disease
- antioxidant properties
- pharmacokinetics
1. Curcumin as a Potential Neuroprotective Agent
2. Chemical and Physical Properties of Curcumin
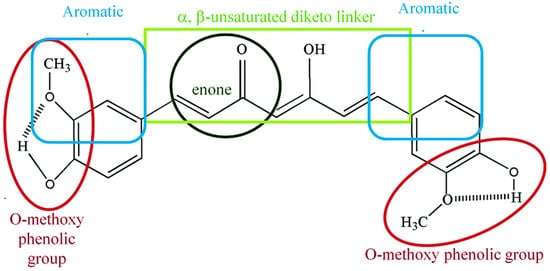
3. Pharmacokinetics and Pharmacodynamics of Curcumin
4. Biological Properties of Curcumin
5. Molecular and Cellular Neuroprotective Mechanisms of Curcumin in PD
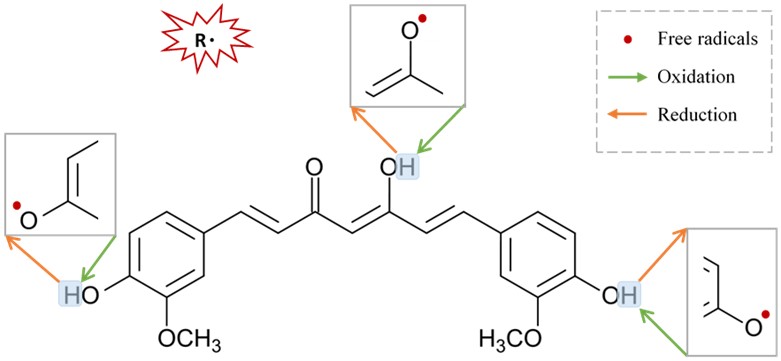 Figure 3. Suggested sites of exchange of phenol OH-group in curcumin structure with free radical oxidants, and its regeneration by a hydrogen donor antioxidant.
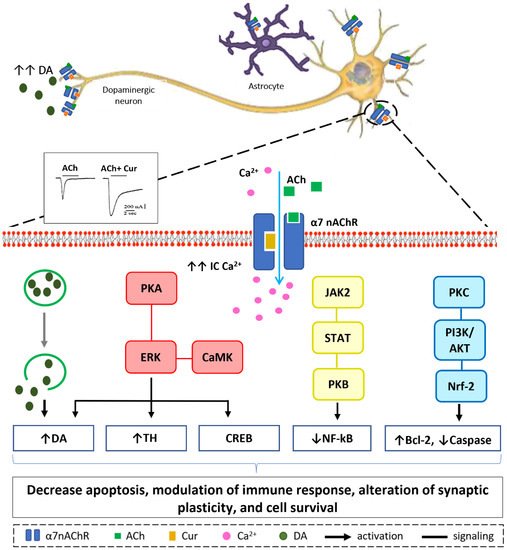
Figure 3. Suggested sites of exchange of phenol OH-group in curcumin structure with free radical oxidants, and its regeneration by a hydrogen donor antioxidant.67. Neuroprotective Mechanisms of Curcumin via Nicotinic Acetylcholine Receptors
Curcumin’s pharmacological actions are thought to be mediated by a variety of ligand-gated ion channels and receptors [66][128]. The recent study on the effects of the natural polyphenol compound provides evidence that curcumin possesses a potent neuroprotective effect as it preserves the integrity of the nigrostriatal dopaminergic system. This is distinctly manifested in the improved motor behavioral performance in the curcumin-treated animals through a α7-nAChRs-mediated mechanism [56][70]. This study adds to previous in vitro studies that show that curcumin enhances the effects of acetylcholine (ACh) through the function of α7-nAChRs in a concentration-dependent manner [67][129]. In addition, the results from another in vitro study highlight the significant role of curcumin in modulating the fluxes of calcium (Ca2+) ions via α7-nAChRs [68][130]. Based on the previous findings that curcumin acts as a type II PAM of α7-nAChRs and a potentiator of receptor function by significantly decreasing desensitization [67][129], it is reasonable to conclude that curcumin’s PAM action on α7-nAChRs has a beneficial effect in mediating neuroprotective effects [69][70][131,132]. Curcumin’s time-tested safety, neuroprotective efficacy, and preliminary clinical success of agents targeting nicotinic receptors in PD make it an appealing natural candidate for further investigation and development in the search for PD therapeutics. Our in vitro, in silico, and in vivo findings suggest that increasing Ca2+ influx may have a neuroprotective mechanism in neuronal and non-neuronal cells via various intracellular mechanisms, as shown in Figure 4 [56][67][68][70,129,130]. Stimulation of presynaptic α7-nAChR stimulates vesicular DA release via a Ca2+-dependent facilitation mechanism [71][72][73][133,134,135]. Extracellular signal-regulated mitogen-activated protein kinase (ERK/MAPK) activation can be triggered by protein kinase A (PKA) and/or calcium-calmodulin-dependent protein kinase (CaMK) [74][136]. A rise in intracellular Ca2+ levels is considered as a trigger factor of both signaling cascades. Activation of (ERK/MAPK) is a crucial signaling event in the cell survival pathway via upregulation of the cellular transcription factor; cAMP response element-binding (CREB), increasing gene expression of tyrosine hydroxylase and enhancing DA release [75][76][137,138]. α7-nAChR is also expressed on microglia and astrocytes and plays a major role in immune response via the “cholinergic anti-inflammatory pathway”. Activation of α7-nAChR results in an increase in intracellular Ca2+ concentration, and consequently modulates Janus kinase 2 (JAK2) and/or signal transducer and activator of transcription 3 (STAT3), ending up with an upregulation of protein kinase B (PKB), leading to inhibition of nuclear factor-kB (NFκB) [77][139]. The lipid signaling cascade that is started by protein kinase C (PKC), via phosphorylation of phosphatidylinositol 3-kinase (PI3K/Akt), is accredited with modulating the activities of neuroprotective and apoptotic factors, such as Bcl-2 and caspases, respectively [78][79][80][140,141,142]. Recent data demonstrate that the regulation of neuroinflammatory reactions by curcumin occurs through the modulation of the microglial JAK/STAT signaling pathway [81][143]. Collectively, all or some of these factors result in decreased apoptosis, enhance neuronal survival, modify immune responsiveness, and produce alteration in synaptic plasticity [82][144].