1. Introduction
Kidney transplantation (KT) is the gold standard treatment of end-stage renal disease (ESRD). In the last two decades, the overwhelming discrepancy between available organs and patients in the transplant waiting list has led to a progressive expansion of the criteria adopted for donors’ acceptance and allocation. As such, an increasing number of so-called marginal kidneys are now being transplanted into high-risk recipients. The main consequence of this aggressive policy is a striking rise in the incidence of delayed graft function (DGF)
[1][2][3][1,2,3]. Clinically defined as the need for dialysis during the first post-transplant week, DGF is a complex entity that can negatively affect both patient and allograft survival
[4]. The most relevant risk factors for DGF are expanded criteria (ECD) or donation after circulatory death (DCD) donors, prolonged warm (WIT) or cold ischemia time (CIT), and recipient sensitization
[5]. Among the various peri-transplant events contributing to the development of DGF, ischemia–reperfusion injury (IRI) deserves special consideration as it is inevitable and represents the major determinant of acute tubular necrosis (ATN), the leading histological finding associated with DGF
[2]. IRI is accompanied by a massive pro-inflammatory response and can trigger multiple cell death programs, endothelial dysfunction, transcriptional reprogramming, and activation of both innate and adaptive immunity
[6]. Considering the strong association between IRI, DGF, acute rejection (AR), and progressive interstitial fibrosis with tubular atrophy (IF/TA) as well as their detrimental impact on allograft function and survival, the prevention and treatment of IRI have been recognized as primary issues for the transplant community
[7].
23. Mechanisms of Action
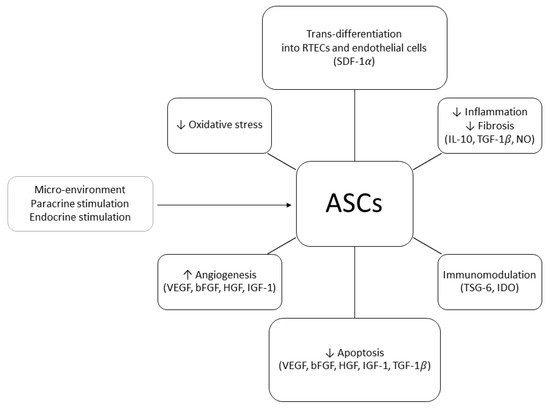
In order to explain the positive effects of ASCs on renal injury, two mechanisms of action have been proposed: differentiative and paracrine. In 2010, Li et al. demonstrated that, in a murine model of acute kidney injury (AKI), human ASCs (hASCs) injected in the tail vein 24 h after the induction of IRI could trans-differentiate into RTECs and repair the damage associated with ATN. At six months of follow-up, the animals treated with hASCs showed normal renal histology whilst the control group exhibited fibrosis and glomerular sclerosis [8][36]. ASCs’ ability to trans-differentiate into RTECs was confirmed by other studies in which mouse or rabbit kidney scaffolds were seeded with ASCs through the renal artery or ureter [9][65]. During these experiments, it was observed that ASCs were able to engraft the glomerular, tubular, and vascular areas of the scaffold, eventually trans-differentiating into RTECs and endothelial cells. The authors postulated that the attachment to the scaffold was mediated by the stromal cell-derived factor 1α (SDF-1α
)
[10][66]. It remains unclear whether ASCs could trans-differentiate into other renal cell types or build up a proper tridimensional structure. ASCs also exert complex paracrine actions
[11][45]. The main soluble molecules produced in response to IRI and mediating ASCs’ pro-angiogenic, proliferative, and anti-apoptotic effects are IGF-1, SDF-1α, and VEGF
[12][13][67,68]. HGF, IL-10, TGF-β1, and NO have been recognized as anti-inflammatory and anti-fibrosis agents
[14][69] whereas tumor necrosis factor-inducible gene 6 protein (TSG-6)
[15][70] and indoleamine 2,3-dioxygenase (IDO)
[16][17][71,72] seem to promote the development of regulatory T-cells (Tregs) and induce tolerance via up-regulation of the FOX-P3 pathway
[18][73].
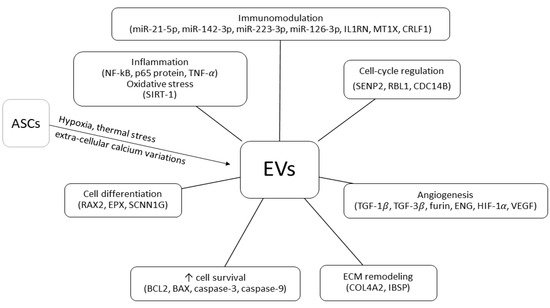
EVs represent further mediators of ASCs’ function
[19][74]. Classified as exosomes (30–100 nm) or micro-vesicles (100–1000 nm) depending on their maximal diameter, EVs can change their content and secretion pattern in response to different external stimulations
[20][21][75,76]. Hypoxia, extra-cellular calcium variations, and thermal stress are powerful triggers of EVs secretion
[22][23][77,78]. It has been hypothesized that ASC-induced renal tissue repair is mediated by specific mRNAs and miRNAs contained in the EVs
[24][79]. In particular, miR-24, miR-29, and several members of the let-7 miRNA family might be involved in the regenerative, proliferative, anti-apoptotic, and anti-inflammatory response to IRI
[25][80] whereas the immunomodulating effects on dendritic cells could be mediated by miR-21-5p, miR-142-3p, miR-223-3p, and miR-126-3p
[26][81]. EV-related mRNAs can contribute to cell differentiation (RAX2, EPX, SCNN1G), immunomodulation (IL1RN, MT1X, CRLF1), ECM remodeling (COL4A2, IBSP), cell-cycle progress (SENP2, RBL1, CDC14B), angiogenesis, and fibrosis (TGF-β1, TGF-β3, FURIN, ENG)
[27][82]. Investigating the role of EVs in AKI, Gao et al. observed that ASC-derived exosomes could significantly decrease the expression of important inflammatory mediators, such as NF-κB, p65 protein, and TNF-α
[28][83]. Exosomes can also up-regulate SIRT-1, thus protecting RTECs from oxidative stress
[29][84]. There is evidence that ASCs’ EVs are involved in cell survival regulation and tissue repair. As a matter of fact, exosomes release is associated with increased expression of the pro-survival protein BCL2 and decreased expression of the pro-apoptotic caspase-3, caspase-9, and BAX
[28][83]. Exosomes also favor angiogenesis induction through up-regulation of HIF-1α and VEGF
[28][83]. According to a recent meta-analysis of pre-clinical rodent models of AKI, ASC-derived EVs administration improves renal function, reducing apoptosis and inflammation
[30][85]. Remarkably, some reports suggest that EV-induced renal protection from IRI exhibits a concentration-dependent effect
[31][41].
Main ASCs and ASC-derived EVs mechanism of actions in IRI and KT models are summarized in Figure 1 and Figure 2.
Figure 1. Adipose-derived stem/stromal cells (ASCs) mechanisms of action in kidney transplant ischemia–reperfusion injury and tolerance induction models (↑, increased; ↓, decreased; RTEC, renal tubular epithelial cell; SDF-1, stromal cell-derived factor 1; IL-10, interleukin 10; TGF-1, transforming growth factor 1; NO, nitric oxide; TSG-6, tumor necrosis factor-inducible gene 6; IDO, indoleamine 2,3-dioxygenase; VEGF, vascular endothelial growth factor; bFGF, basic fibroblast growth factor; HGF, hepatocyte growth factor; IGF-1, insulin-like growth factor).
Figure 2. Adipose-derived stem/stromal cell extra-cellular vesicles (EVs) mechanisms of action in kidney transplant ischemia–reperfusion injury and tolerance-induction models (↑, increased; NF-kB, nuclear factor kappa-light-chain-enhancer of activated B cells; p65 protein, REL-associated protein; TNF, tumor necrosis factor; SIRT-1, sirtuin-1; RAX2, retina and anterior neural fold homeobox protein 2; EPX, eosinophil peroxidase; SCNN1G, sodium channel epithelial 1 subunit gamma; BCL2, B-cell lymphoma 2; BAX, bcl2-like protein 4; COL4A2, collagen type IV alpha 2 chain; IBSP, integrin binding sialoprotein; TGF-1, transforming growth factor 1; TGF-3, transforming growth factor 3; ENG, endoglin; HIF-1, hypoxia-inducible factor-1; VEGF, vascular endothelial growth factor; SENP2, SUMO specific peptidase 2; RBL1, retinoblastoma-like protein 1; CDC14B, dual specificity protein phosphatase CDC14B; miR, micro RNA; IL1RN, interleukin 1 receptor antagonist; MT1X, metallothionein 1X; CRLF1, cytokine receptor like factor 1).
34. Harvesting Procedures
ASCs reside in peri-vascular niches within the adipose tissue, constituting about 20% of the cellular population of the SVF. Two main ASCs harvesting techniques are currently available: lipoaspirate and adipose tissue biopsy. Following specimen collection, the ECM of the adipose tissue undergoes enzymatic or mechanical disruption. The SVF released during the procedure is seeded and expanded in vitro to obtain ASCs [32][59]. Cultivation and expansion allow to produce a large number of ASCs from a relatively small sample of adipose tissue and permit to collect purified cellular products [33][61]. However, these methods are expensive, time-consuming, and require specific Good Manufacturing Practice (GMP) facilities. Therefore, it is unlikely that they can be used in the peri-operative setting of a deceased donor KT [34][43]. Allogenic non-donor derived ASCs represent an intriguing option as off-the-shelf cellular products. Nonetheless, some authors remain concerned that prolonged culture may cause cellular senescence and favor the acquisition of a pro-inflammatory phenotype [35][86].
The process required for SVF extraction is easy and expeditious enough that it can be performed in a timely manner in most hospitals. The SVF is a composite cellular mixture with wide intra- and inter-individual variability in cells quality and quantity, depending on patient’ characteristics, harvesting procedure, extraction technique, and isolation method
[36][87]. Such heterogeneity represents a remarkable issue as it negatively affects standardization and reproducibility of the vast majority of research protocols involving the SVF. Furthermore, the amount of adipose tissue required to obtain an adequate number of ASCs is significantly greater using the SVF extraction method than the ASCs expansion technique. Lastly, it is worth considering that the effects exerted on the transplanted kidney by the other cell types retrieved during the procedure (namely, pericytes, preadipocytes, endothelial cells, and fibroblasts) are still unclear
[34][43]. Immunomodulatory, anti-inflammatory, and pro-tolerogenic properties have been postulated for the non-staminal cells of the SVF such as adipose resident M2 macrophages, Tregs, and regulatory invariant natural killer (NK) cells, but they have not been confirmed.
[37][38][39][88,89,90].
The use of stem cell-based therapies in solid organ transplantation raises the question of whether autologous or allogenic cells should be preferred. Due to the lack of HLA class II molecules and the low expression of HLA class I and co-stimulatory signal molecules, ASCs are considered as immune-privileged cells
[40][91]. However, the actual risk of allograft rejection associated with the use of allogenic ASCs remains obscure
[41][42][43][92,93,94]. It has been demonstrated that donor-derived ASCs can promote the development of Tregs in vitro
[18][73]. A certain degree of tolerance induction has also been also observed in vivo
[44][95]. Nevertheless, a study comparing the efficacy of autologous or allogenic ASCs in a rat model of KT with total MHC mismatch, showed that donor-derived ASCs administration did not improve allograft survival and was associated with increased mortality through an immunologically-mediated mechanism
[45][96]. Considering the paucity of data, it sounds reasonable to assume that autologous ASCs represent a safer option compared to allogenic ASCs. Some authors argue that, due to the toxic effects of uremia, ASCs harvested from ESRD patients have lower regenerative potential than those retrieved from healthy subjects; thus limiting the use of autologous ASC-based therapies in KT setting. However, there is evidence that the beneficial properties of ASCs are not affected by uremia or dialysis. In fact, Roemeling-van Rhijn and colleagues demonstrated that ASCs harvested from patients with CKD exhibited normal characteristics and function. Remarkably, ASCs’ proliferative capacity and genetic stability remained consistent over time, also after repeated expansion and prolonged exposure to uremic serum. Furthermore, no differences could be detected between patients with normal or impaired renal function in ASCs’ response to immune activation and ASCs’ inhibitory effect on the proliferation of alloantigen-activated peripheral blood mononuclear cells
[46][97].
In theory, the use of ASC-derived EVs could help address many issues associated with traditional cell-based therapies such as the need for time-consuming and technically demanding preparation processes and the risk of HLA sensitization. A recent meta-analysis of EV-based therapies in pre-clinical rodent models of AKI concluded that MSC-derived EVs administration was able to improve renal function, modulate the inflammatory response
[47][98], and reduce cell apoptosis
[30][85]. Overall, available studies suggest that EVs could be beneficial for the prevention and treatment of renal IRI
[48][49][99,100]. On the contrary, current literature does not seem to support the utilization of EV-based therapies for KT rejection
[45][96]. The highly variable composition of different EVs mixtures should also be considered as a potential limitation
[22][77].
45. Timing, Dose, and Route of Administration
The proper definition of the optimal timing, dose, and route of administration represents a key factor for the implementation of ASC-based therapies in KT. Possible routes of administration that can maximize ASCs homing and survival in the allograft include intra-venous, intra-arterial, and intra-parenchymal. Continuous infusion using a kidney perfusion machine has also been proposed. There is evidence that MSCs and ASCs homing to the wounded areas of the kidney is promoted by a specific chemoattractant. Among the pathways involved in MSCs migration during IRI, the SDF-1/CXCR4 axis plays a major role. Over-expression of the CXCR4 gene enhances MSCs migration and promotes neo-angiogenesis and vascular repair
[50][51][52][101,102,103]. These signals also regulate ASCs homing from the injection site to the transplanted organ
[53][104].
The easiest route of administration of ASCs is intra-venous injection. However, when injected into the venous circulation, ASCs undergo a first pass effect in “filter organs” such as the liver, spleen, or lung. This phenomenon reduces the number of cells reaching the target site and can cause engulfment of the pulmonary micro-circulation, eventually leading to pulmonary embolism and death
[54][105]. The results of clinical trials evaluating efficacy and safety of intravenously administered ASCs for the treatment of graft-versus-host disease demonstrate that the risk of micro-thrombosis and micro-embolism depends on the total number of cells infused. According to these studies, doses ranging from 0.4 to 9 × 10
6 MSCs or ASCs per kilogram represent a safe option
[54][55][56][105,106,107].
Intra-arterial administration can also be considered. In theory, ASCs injection into the renal artery could increase the number of cells engrafting the kidney. A meta-analysis of MSC-based therapies in small animal models of AKI concluded that the intra-arterial route was associated with better renal function preservation (as assessed by serum creatinine concentration, SCr) than the intra-venous route
[57][108]. It seems that, following injection into the renal artery, ASCs remain on site through passive mechanisms, mostly disappearing within 14 days
[58][109]. Sporadic episodes of mesangioproliferative glomerulonephritis with aberrant adipocytic differentiation have been reported
[59][110]. It can also be argued that, compared to intra-venous injection, intra-arterial administration is more invasive, entailing a greater risk of complications. Nevertheless, it should be recognized that a KT procedure offers wide kidney exposure and easy access to the renal artery.
ASCs intra-parenchymal injection represents another feasible option and it proved effective in several animal models of AKI
[12][67]. This route of administration allows to combine ASCs with special biomaterials so as to extend their permanence and survival in the target site. An elegant study by Gao et al. showed that the use of thermosensitive chitosan chloride hydrogel (CCH) could improve the efficacy of ASC-based therapy in IRI. More in details, CCH was able to increase intra-renal ASCs retention and survival as well as to promote host kidney cells proliferation and resistance to apoptosis. At four weeks, the groups treated with CCH exhibited higher renal micro-vessels density and RTECs proliferation rates than controls, with better renal function. A possible explanation is that CCH favors a more suitable micro-environment for ASCs survival through enhanced ROS scavenging
[60][111]. Encouraging results were also obtained by Zhou and colleagues using renal ECM hydrogel as an injectable scaffold to deliver ASCs into ischemic kidneys. In their experience, ECM hydrogel reduced ASCs dispersion and increased the number of viable ASCs retained in the target site. Furthermore, the authors observed that ECM hydrogel could inhibit ASCs apoptosis in vitro and promote ASCs proliferation, trans-differentiation into RTECs, and secretion
[61][112]. Overall, available data suggest that ASCs cultivation in three-dimensional aggregates or spheroids combined with direct injection into the renal cortex can increase ASCs retention in the kidney and improve the efficacy of ASC-based therapies. Compared to bi-dimensional monolayers, three-dimensional configurations provide better resistance to oxidative stress and ischemia. They also facilitate cell migration and secretion
[62][63][113,114].
ASCs can be delivered to the renal allograft using specific perfusion machines
[64][115]. Several randomized clinical trials and meta-analysis have demonstrated that HMP reduces the incidence of DGF compared to SCS, particularly in case of ECD or DCD transplants
[65][116]. Therefore, combining the benefits of continuous or pulsatile kidney perfusion with the regenerative potential of ASCs-based therapy represents a promising strategy. Preliminary data obtained from animal models of KT or acute kidney IRI showed that adding ASCs, BM-MSCs, or BM-MSC-derived EVs to the machine perfusion solution could improve renal function and allograft survival
[66][67][68][117,118,119]. Current experience with NMP is more limited. Indeed, higher costs and logistic difficulties prevent widespread utilization of this preservation modality. Pool and coworkers investigated the effects of NMP plus hASCs in a model of porcine DCD KT. The experimental treatment was administered following kidney preservation with SCS
[69][120] or HMP
[13][68]. Interestingly, positive results were observed only in the HMP group. Compared to SCS, HMP ensured better ASCs homing and viability as well as higher resistance to oxidative stress and improved tubular regeneration capacity.