Gluten proteins, major determinants of the bread-making quality of wheat, are related to several digestive disorders. Advances in plant genetic breeding have allowed the production of wheat lines with very low gliadin content through the use of RNAi and gene editing technologies.
- gluten
- coeliac disease
- NCWS
- transgenic wheat
- RNAi
- CRISPR/Cas9
1. Wheat and Wheat Proteins
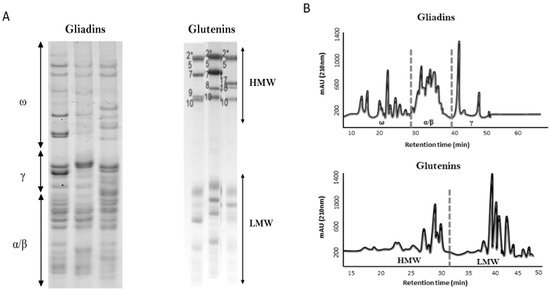
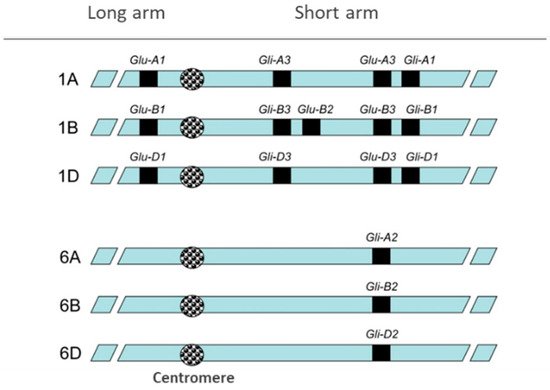
2. Wheat Pathologies and Gluten-Free Diet (GFD)
3. Towards Obtaining Wheat Lines with Low Immunogenic Peptides
4. Stability of the Gliadin Silencing
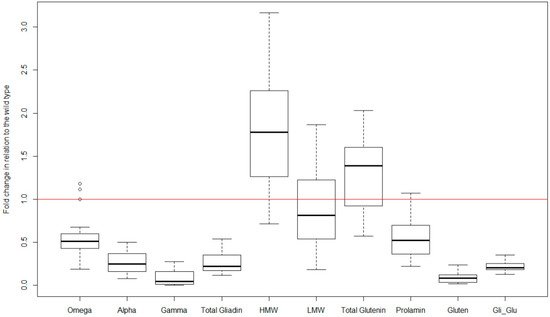
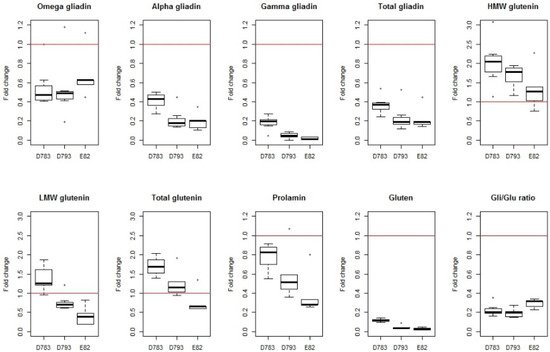
|
Variable |
Min. |
1st Qu. |
Median |
Mean |
3rd Qu. |
Max. |
|---|---|---|---|---|---|---|
|
Omega (mg/g) |
7.7 |
10.8 |
14.1 |
14.9 |
19.5 |
21.7 |
|
Alpha (mg/g) |
14.9 |
27.5 |
37.7 |
33.3 |
39.1 |
42.6 |
|
Gamma (mg/g) |
8.9 |
25.5 |
26.7 |
24.5 |
27.8 |
28.8 |
|
Total gliadin (mg/g) |
31.5 |
65.0 |
77.8 |
72.4 |
83.5 |
93.1 |
|
HMW (mg/g) |
6.3 |
7.1 |
12.4 |
12.3 |
17.0 |
19.4 |
|
LMW (mg/g) |
12.6 |
13.0 |
18.6 |
18.5 |
21.9 |
28.1 |
|
Total glutenin (mg/g) |
18.9 |
20.0 |
30.9 |
30.8 |
38.8 |
47.5 |
|
Prolamin (mg/g) |
51.9 |
84.0 |
108.7 |
103.1 |
121.8 |
140.6 |
|
Gluten (mg/kg) |
45,266 |
67,134 |
134,673 |
111,548 |
149,843 |
166,942 |
|
Gliadin to Glutenin ratio |
1.6 |
2.0 |
2.2 |
2.5 |
3.4 |
3.5 |
HMW, high molecular weight, LMW, low molecular weight.