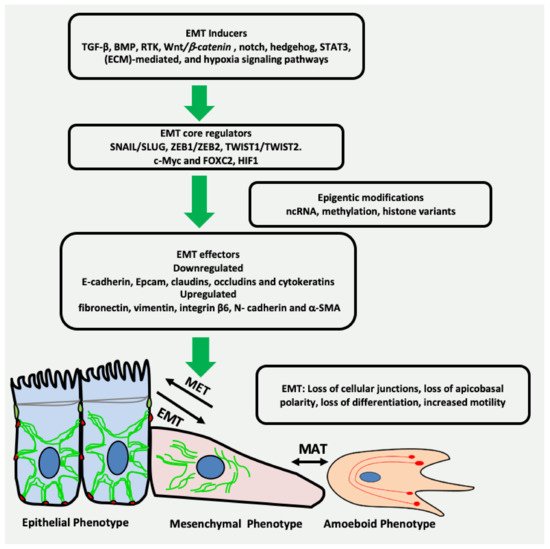
Cancer stem cells (CSC) are a small population of cells capable of self-renewal, which are known to resist therapeutic interventions and immune responses. Being pluripotent, these cells can provide cellular seeds to initiate new tumours at distant sites
[23]. It was proposed that EMT can transform non-CSCs into cancer stem cells, which are invariably vimentin-positive
[24]. In addition, it is believed that CSCs are generated as a result of adaptations and crosstalks with a tumour microenvironment, as well as in therapeutic interventions resulting in the generation of a heterogeneous subpopulation. Hypoxic conditions particularly contribute to the development of CSC characteristics including self-renewal, EMT, and drug resistance
[25]. Hypoxia-inducible factors (HIFs) are the primary mediators of cellular responses, such as proliferation, EMT and metastasis, to hypoxic conditions
[26]. Several other pathways implicated in the regulation of stemness phenotypes via HIFs include the TGF-β
[27], Wnt/β-catenin
[28], TNFα and NF-κB signalling
[29]. These signalling cascades are also implicated in the induction of EMT via the transcriptional control of EMT-associated transcription factors, such as SNAI1, TWIST, ZEB1, SLUG and TCF3 leading to vimentin expression, as described above (
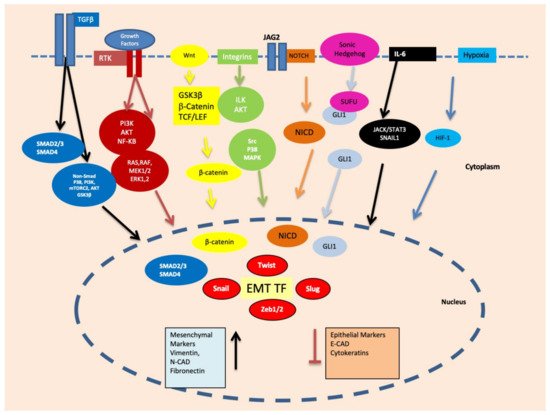
Figure 2).
Figure 2. Important signalling pathways, such as Smad, PI3K/mTORC2/AKT/GSK3β, RAS/RAF/MEK1/2/ERK1, PI3K/AKT/NF-κB, ILK/AKT, Src/P38/MAPK, NOTCH, GLI1/SNAI2, JACK/STAT3 and HIF-1 are activated during EMT via growth factors, hypoxia and other microenvironmental factors. These signalling pathways ultimately upregulate the transcription factors related to EMT such as Snail, Slug, Twist, ZEB1 and ZEB2.
Cancer stem cells can exist in epithelial, mesenchymal or hybrid (mixed) states. They have the ability to switch between these different cellular states to maintain their survival, escape the immunity and grow at secondary sites. This switching is conducted by the complex interactions of various transcription factors, signalling pathways and microenvironmental factors
[30]. The epithelial-like state of CSCs is characterized by the downregulation of mesenchymal markers, such as vimentin, and the upregulation of epithelial markers, such as CDH1. The expression of these markers is reversed in the mesenchymal state of CSCs. Both epithelial and mesenchymal markers are expressed in hybrid state
[31]. CSCs express different cell surface proteins, such as CD34, CD44, CD24 and CD133, transcriptional factors, such as SOX2, NANOG, OCT ¾, SALL4 and other proteins that are not characterised as cell surface proteins, or transcription factors such as, ALDH, BMI1, Nestin and CXCR. These diverse markers are used to distinguish CSCs from the rest of the tumour population in different cancers
[32]. These markers are expressed in a tissue-specific manner e.g., CD44, CD24 and ALDH are specific to breast cancer, CD34, CD8 to leukaemia, CD133 to colon cancer, CD44 to head neck cancer and CD90 to liver cancer
[33]. These markers can be variably expressed according to the state of the CSCs. For example, in breast cancer stem cells, the CD24-CD44+ signature is related to a mesenchymal state with higher vimentin expression while the ALDH+ signature corresponds to an epithelial state of the cancer stem cells
[31]. In normal cells, CD44 is a glycoprotein receptor for hyaluronic acid that is involved in cell adhesion, proliferation, differentiation, migration, angiogenesis and cell survival
[34]. It is overexpressed in a variety of cancers, such as breast, colon, bladder, gastric, glioma, head and neck, prostate and leukaemia
[32]. A soluble form of CD44 exists and is also overexpressed in certain cancers
[35]. This soluble CD44 can bind to the vimentin head domain on the surface of endothelial cells, which is consistent with the fact that both CD44 and vimentin are overexpressed in oral squamous cell carcinoma (OSCC) and prostate cancer; however, the molecular basis of this association has not been fully elucidated
[35]. CD24 is a cell adhesion sialoglycoprotein identified as a differentiation marker for hematopoietic and neuronal cells
[36]. A higher CD44/CD24 ratio is positively correlated with vimentin in the breast CSC population
[37]. ALDH1 (specifically isoform ALDH1A1) is another recently identified CSC marker in different tumours
[38][39][40][38,39,40] and regulates the oxidation of retinal substrates into retinoic acid
[40]. The increased expression of ALDH1 is related to the MET state of CSCS expressing lower levels of vimentin in breast cancer
[41]. The sex-determining region Y-box 2 (SOX2) is an important transcription factor essential for the potential of stem cell multi-lineage. It can reprogram primary cells into stem cells
[42], its levels are frequently upregulated in carcinomas such as HNSCC
[43] with an inverse correlation between SOX2 and vimentin expression
[44], and with a loss of SOX2 inducing tumour invasion through the upregulation of vimentin expression
[45]. The possible role of miR-378 in the SOX2/Vim inverse functional relationship was also reported
[31][46][31,46].