Arginine is one of the functional amino acids that enhances the growth of fetus and placenta development. Since the fetal growth and the nutrient requirement for fetuses are increased hugely during the late gestation period in high-prolific sows, supplementation of L-arginine could have a positive influence on the reproductive performance of sows and piglet uniformity.
- arginine
- late gestation
- sow
- reproductive performance
- piglet uniformity
1. Introduction
1. Definition
Arginine is one of the functional amino acids that signals embryonic and fetal development [11], and it was reported that arginine could partly be attributed to improving piglet birth weight and the uniformity of piglets at birth [12,13]. Dietary arginine intake was shown to increase the synthesis of nitric oxide (NO) and polyamines [14,15]. The nitric oxide thus produced resulted in increased blood flow to the placenta [16,17] and improved delivery of essential nutrients from maternal to fetal blood [18]. Polyamines also showed positive effects on embryogenesis and placental growth [19]. Thus, arginine was shown to influence placental growth and fetal development via the above mechanism [20,21].
2. Introduction
The high prolific sows are developed to improve the number of piglets and sow productivity in the swine industry [1]. However, as they have greater litter size, some problems were reported that the proportion of small piglets at birth, within-litter variation of piglet birth weight, and mortality of suckling piglets were also increased [2][3][4]. These problems are caused by several maternal factors, such as inadequate nutrient intake of late gestation, high number of fetus, insufficient cervical space, or insufficient reproductive tract for fetus development [5][6][7]. Therefore, various studies were conducted to minimize these problems and improve the piglet uniformity of high prolific sows with nutritional strategies including sow body condition, nutrients type or ratio, and functional amino acids [8][9][10].
Fetal growth is rapid during the late gestation period and nutrient requirement for the fetus also increases greatly [8]. The low birth weight of piglets was reported to increase the proportion of stillborn piglets considered to be at a greater risk of mortality and morbidity [9][10]. Since supplying adequate nutrient to sows during the late gestation period is important, an increased feeding method was suggested in order to meet their nutrient requirement. Although increased feeding during late gestation is likely to meet the nutrient requirement of sows and improve piglet birth weight [11][12], it also showed a negative effect on sow body condition and postpartum agalactia [13][14], resulting in poor milk production and lactation feed intake [15][16]. With the effects of arginine, if the nutrient delivery efficiency from the dam to the fetus in late gestation increased by additional arginine supplementation, it could partly be attributed to increase in the birth weight of small fetuses and piglet uniformity at birth.
Fetal growth is rapid during the late gestation period and nutrient requirement for the fetus also increases greatly [22]. The low birth weight of piglets was reported to increase the proportion of stillborn piglets considered to be at a greater risk of mortality and morbidity [23,24]. Since supplying adequate nutrient to sows during the late gestation period is important, an increased feeding method was suggested in order to meet their nutrient requirement. Although increased feeding during late gestation is likely to meet the nutrient requirement of sows and improve piglet birth weight [25,26], it also showed a negative effect on sow body condition and postpartum agalactia [27,28], resulting in poor milk production and lactation feed intake [29,30]. With the effects of arginine, if the nutrient delivery efficiency from the dam to the fetus in late gestation increased by additional arginine supplementation, it could partly be attributed to increase in the birth weight of small fetuses and piglet uniformity at birth. Most of the previous studies for evaluating the effect of arginine in gestating sows were investigated in the early-gestation period [31,32,33,34] or whole gestation period [12,13,35,36]. Additionally, most previous studies for the effect of arginine supplementation in sows, investigated the effect of 1% arginine supplementation in sow diet. Thus, there is a need to investigate the effects of dietary arginine below 1% of supplementation level keeping in mind the market cost of L-arginine, in order to apply this in the field.Most of the previous studies for evaluating the effect of arginine in gestating sows were investigated in the early-gestation period [17][18][19][20] or whole gestation period [21][22][23][24]. Additionally, most previous studies for the effect of arginine supplementation in sows, investigated the effect of 1% arginine supplementation in sow diet. Thus, there is a need to investigate the effects of dietary arginine below 1% of supplementation level keeping in mind the market cost of L-arginine, in order to apply this in the field.
3. Results
The arginine supplementation levels in the late-gestating sows’ diet did not affect body weight, backfat thickness, and the lactation feed intake of sows (Table 1).
Table 12.
| Treatment | 1 | SEM | 2 | p | -Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p | -Value | CON | ARG10 | ARG15 | Diet | Lin. | Quad. | ||||||||||
| CON | |||||||
|---|---|---|---|---|---|---|---|
| Body Weight, kg | |||||||
| Day 70 | 237.1 | 239.1 | 238.4 | 3.58 | 0.98 | 0.90 | 0.85 |
| Day 110 | |||||||
In reproductive performance, dietary arginine levels during late gestation had no influence on the number of piglets for total born, stillborn, mummy, and born alive (Table 2). However, piglet birth weight showed a quadratic response (
p
p < 0.10) the alive litter weight linearly. Increasing the dietary arginine levels in late gestation did not affect the farrowing time, helping frequency, and placenta weight.
Table 23.
| Treatment | |||||
|---|---|---|---|---|---|
| 1 | |||||
| SEM | |||||
| 2 | |||||
| p | |||||
| -Value | |||||
| CON | ARG10 | ARG15 | Diet | Lin. | Quad. |
Table 34.
| Treatment | 1 | SEM | 2 | p | -Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | ARG10 | ARG15 | Diet | Lin. | Quad. | |||||||||
| No. of Pigs |
| Total born |
| 13.7 |
| No. of Piglets | |||||||
| 15.0 | 14.8 | ||||||
| After-fostering | 11.5 | 11.6 | 11.6 | 0.10 | 0.90 | 0.77 | 0.72 |
| 3.98 | |||||||
| A | |||||||
| 0.093 | |||||||
| 0.08 | |||||||
| 0.03 | 0.66 | ||||||
The effect of dietary arginine levels in late gestation on the piglet uniformity is presented in Table 4. The SD for piglet birth weight was linearly increased (
p
p
p
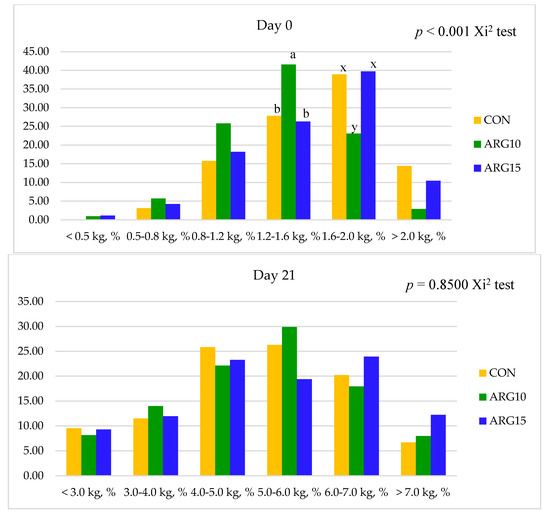
Figure 1. Effect of arginine supplementation level in late gestating sows on piglet distribution into body weight classes on day 0 and day 21 of lactation by the FREQ procedure, with Chi-square test. CON: corn-SBM-based diet with Arg 0.72%, Arg10: corn-SBM-based diet with Arg 1.0%, Arg15: corn-SBM-based diet with Arg1.5%.
Table 45.
| Treatment | 1 | SEM | 2 | p | -Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CON | ARG10 | |||||||||||||
Table 78.
| Treatment | 1 | SEM | 2 | p | -Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ARG15 | Diet | Lin. | Quad. | |||||||||||
| CON | ARG10 | ARG15 | Diet | Lin. | Quad. | Group | Date | Group × Date |
|---|---|---|---|---|---|---|---|---|
| CON | ARG10 | ARG15 | Diet | Lin. | Quad. | Group | Date | Group × Date |
| ARG10 | ARG15 | Diet | Lin. | Quad. |
|---|
| Piglet Uniformity at Birth | |||||||
| 0.50 | |||||||
| Avg. BW, kg | 0.53 | 1.57 | 1.38 | 1.52 | - | ||
| Alanine, μmol/L | ||||||||||||||||
| 0.46 | ||||||||||||||||
| Day 70 | 0.41 | 609.5 | - | - | - | 487.0 | 454.0 | 46.75 | 0.49 | 0.27 | 0.56 | 0.09 | <0.01 | 0.51 | 258.2 | 256.3 |
| Blood urea nitrogen, mg/dL | ||||||||||||||||
| Casein, % | |||||||||||||||||||||||||||||||||||||
| Day 70 | 12.5 | 11.7 | 11.9 | 0.45 | 0.85 | 0.72 | 0.63 | 0.99 | 0.01 | 0.93 | |||||||||||||||||||||||||||
| Colostrum | 6.57 | 6.08 | 5.62 | 0.571 | 0.81 | 0.52 | 0.91 | 252.7 | 3.33 | 0.79 | Day 21 of lactation | 10.7 | 0.50 | 10.4 | |||||||||||||||||||||||
| SD | 274.0 | 10.9 | 0.16 | 0.52 | 0.99 | ||||||||||||||||||||||||||||||||
| 0.49 | 293.5 | 0.36 | |||||||||||||||||||||||||||||||||||
| 334.3 | 12.72 | 0.13 | 0.04 | 0.94 | |||||||||||||||||||||||||||||||||
| Day 90 | 709.1 | 710.5 | 652.0 | 37.24 | 0.79 | 0.53 | 0.79 | Day 90 | 11.3 | 12.0 | 12.5 | 0.57 | 0.69 | 0.41 | 0.83 | ||||||||||||||||||||||
| Milk (21 days) | 4.05 | 4.32 | 4.09 | ||||||||||||||||||||||||||||||||||
| 0.119 | 0.63 | 0.98 | BW gain (70–110 days) | 21.1 | 17.3 | Litter Weight, kg | |||||||||||||||||||||||||||||||
| 0.34 | CV | 18.4 | 21.8 | 14.3 | 2.46 | 0.53 | 0.27 | 22.9 | 0.79 | ||||||||||||||||||||||||||||
| 1.01 | 0.15 | 0.07 | 0.41 | ||||||||||||||||||||||||||||||||||
| Day 110 | 878.4 | A | 916.3 | A | 628.7 | B | 53.05 | 0.06 | 0.03 | Day 110 | 14.3 | 14.5 | 14.1 | 0.57 | 0.56 | 0.84 | 0.83 | ||||||||||||||||||||
| Fat, % | 0.26 | After-fostering | 17.67 | a | 16.01 | b | 17.73 | a | 0.326 | 0.04 | 0.63 | 0.01 | Creatinine, mg/dL | 224.9 | 218.5 | 3.97 | 0.53 | ||||||||||||||||||||
| Piglet Distribution into Birth Weight Classes | |||||||||||||||||||||||||||||||||||||
| Arginine, μmol/L | |||||||||||||||||||||||||||||||||||||
| Colostrum | 6.68 | 5.43 | 6.91 | 0.485 | 0.43 | 0.26 | 0.95 | ||||||||||||||||||||||||||||||
| 0.69 | 0.23 | Day 70 | 154.5 | 255.0 | 195.8 | 23.65 | Day 70 | 1.76 | 1.74 | ||||||||||||||||||||||||||||
| Milk (21 days) | 4.04 | 1.95 | 0.091 | 0.59 | 0.41 | 6.82 | 0.70 | 6.32 | 0.368 | 0.13 | 0.23 | 0.48 | BW loss (0–21 days) | −4.3 | −5.9 | −9.4 | 1.67 | 0.44 | 0.20 | 0.96 | |||||||||||||||||
| Backfat Thickness, mm | |||||||||||||||||||||||||||||||||||||
| Day 70 | 20.7 | 21.4 | 19.1 | 0.77 | 0.49 | 0.35 | 0.46 | ||||||||||||||||||||||||||||||
| Day 110 | 22.4 | 22.6 | 20.7 | 0.84 | 0.59 | 0.37 | 0.66 | ||||||||||||||||||||||||||||||
| BF gain (70–110 days) | 2.1 | 1.3 | 1.6 | 0.3 | |||||||||||||||||||||||||||||||||
| Stillborn | 0.7 | 0.7 | 0.8 | 0.12 | 0.93 | 0.73 | 0.89 | ||||||||||||||||||||||||||||||
| Mummy | 0.2 | 0.3 | 0.1 | 0.08 | 0.49 | 0.31 | 0.53 | 24 h postpartum | 233.7 | 230.8 | 227.9 | 3.73 | 0.82 | 0.54 | 0.92 | ||||||||||||||||||||||
| Born alive | 12.8 | 14.0 | 13.9 | 0.45 | 0.35 | 0.31 | 0.31 | Day 21 of lactation | |||||||||||||||||||||||||||||
| Total litter weight, kg | 229.4 | 19.61 | 19.91 | 21.49 | 0.521 | 0.30 | 0.13 | 0.73 | |||||||||||||||||||||||||||||
| Day 21 of lactation | 53.74 | B | 53.31 | B | 60.27 | A | |||||||||||||||||||||||||||||||
| <0.5 kg, % | 0 | 1.0 | 1.332 | 0.05 | 1.2 | 0.30 | 0.240.02 | 0.32 | 0.310.15 | 0.38 | 0.66 | 0.14 | <0.01 | <0.01 | 0.27 | <0.01 | 0.89 | Alive litter weight, kg | 18.47 | 19.01 | 20.69 | 0.506 | 0.19 | 0.07 | 0.81 | ||||||||||||
| Weight gain (0–21 days) | 36.06 | B | 37.17 | AB | 42.58 | A | 1.262 | 0.07 | 0.02 | ||||||||||||||||||||||||||||
| 0.10 | 0.64 | ||||||||||||||||||||||||||||||||||||
| 0.5–0.8 kg, % | 3.1 | 5.6 | 4.2 | 0.79 | 0.44 | 0.59 | 0.25 | ||||||||||||||||||||||||||||||
| Day 90 | 262.9 | Day 90 | b | 349.1 | b | 482.7 | a | 29.44 | <0.01 | <0.01 | 0.88 | 2.23 | 2.43 | Piglet birth weight, kg | 1.57 | ||||||||||||||||||||||
| A | |||||||||||||||||||||||||||||||||||||
| Protein | 2.39 | 0.104 | 0.74 | 0.63 | 0.55 | , % | |||||||||||||||||||||||||||||||
| 1.38 | |||||||||||||||||||||||||||||||||||||
| B | |||||||||||||||||||||||||||||||||||||
| 1.52 | |||||||||||||||||||||||||||||||||||||
| A | |||||||||||||||||||||||||||||||||||||
| 0.036 | 0.08 | 0.85 | 0.03 | ||||||||||||||||||||||||||||||||||
| Farrowing time, min | |||||||||||||||||||||||||||||||||||||
| Piglet Weight, kg | |||||||||||||||||||||||||||||||||||||
| 0.8–1.2 kg, % | 15.8 | 25.8 | 18.2 | Day 110 | 196.4 | 2.36 | b | 301.2 | 0.21 | a | 366.9 | 0.87 | a | 24.34 | 0.07 | Day 110 | 2.97 | 2.90<0.01 | <0.01 | 0.34 | 3.18 | 0.097 | 0.48 | 0.32 | 0.50 | ||||||||||||
| Colostrum | 8.28 | 7.91 | 7.29 | 0.804 | 0.89 | 0.63 | 0.99 | 182.4 | 186.2 | 205.7 | |||||||||||||||||||||||||||
| 1.2–1.6 kg, % | 27.9 | b | 41.5 | a | 26.3 9.33 | 0.58 | 0.31 | 0.82 | |||||||||||||||||||||||||||||
| After-fostering | 1.55 | a | 1.38 | b | 1.53 | a | 0.031 | 0.04 | b | 0.82 | 0.01 | 4.39 | 0.03 | 0.49 | 0.01 | ||||||||||||||||||||||
| Aspartic Acid, μmol/L | Total protein, g/dL | ||||||||||||||||||||||||||||||||||||
| Milk (21 days) | 4.72 | 4.77 | 4.61 | 0.115 | 0.86 | 0.68 | 0.72 | Helping frequency | Day 70 | 0.80 | 0.81 | 1.00 | 0.141 | 0.82 | 0.55 | 0.85 | |||||||||||||||||||||
| Day 21 of lactation | 5.03 | 5.14 | 5.51 | 0.103 | 0.13 | 0.04 | 0.78 | 0.51 | |||||||||||||||||||||||||||||
| 1.6–2.0 kg, % | 39.0 | ab | |||||||||||||||||||||||||||||||||||
| Day 70 | 23.0 | b | 14.039.7 | a | 6.8514.35.55 | 0.08 | 0.64 | 13.5 | 1.330.03 | 0.97 | 0.88 | 0.89 | 0.69 | 0.15 | 0.74 | 6.67 | 7.15 | 0.256 | 0.76 | 0.63 | 0.67 | 0.95 | 0.98 | 0.65 | |||||||||||||
| Lactose, % | Placenta weight, kg | 3.74 | 3.74 | 3.900.57 | 0.30 | ||||||||||||||||||||||||||||||||
| 0.224 | 0.95 | 0.76 | 0.90 | ||||||||||||||||||||||||||||||||||
| Weight gain (0–21 days) | 24 h postpartum | 21.0 | 21.1 | Placenta efficiency | 5.80 | 20.3 | 0.85 | 0.91 | 0.72 | 5.89 | 0.84 | ||||||||||||||||||||||||||
| 6.02 | 0.246 | 0.94 | 0.73 | 0.98 | |||||||||||||||||||||||||||||||||
p < 0.05) the litter weight at day 21, and the litter weight gain, such that litter weight and litter weight gain for ARG15 tended to be greater than those for CON (Table 3). The piglet weight at day 21 of lactation was linearly increased (
p
p < 0.05) piglet weight gain, such that the piglet weight gain for ARG15 tended to be greater than that for CON.
| 3.48 |
| B | |||||||||||||||||||||||
| 3.74 | |||||||||||||||||||||||
| AB | |||||||||||||||||||||||
| >2.0 kg, % | |||||||||||||||||||||||
| 14.5 | |||||||||||||||||||||||
| 2.9 | |||||||||||||||||||||||
| 10.4 | 5.67 | 0.15 | 0.70 | ||||||||||||||||||||
| Day 90 | 0.06 | 18.8 | 31.4 | 18.3 | 4.79 | 0.46 | 0.82 | 0.23 | |||||||||||||||
| Day 90 | 6.86 | 7.23 | 6.71 | 0.170 | 0.47 | 0.60 | 0.27 | Day 110 | 14.4 | 14.0 | |||||||||||||
| Colostrum | 4.72 | 4.35 | 4.31 | 0.187 | 0.65 | 0.44 | 0.60 | 14.4 | 0.87 | 0.98 | Day 110 | 6.93 | 6.73 | Day 21 of lactation | 20.1 | 19.8 | 18.6 | 0.79 | 0.71 | 0.43 | |||
| Piglet Uniformity at day 21 | 0.90 | ||||||||||||||||||||||
| 0.96 | 0.85 | 7.00 | 0.175 | 0.82 | 0.80 | Avg. BW, kg | 5.03 | 5.14 | 5.51 | - | - | - | - | BF loss (0–21 days) | −0.9 | −1.3 | −1.7 | 0.38 | 0.69 | ||||
| Citrulline, μmol/L | 0.40 | 0.85 | |||||||||||||||||||||
| SD | 1186.8 | 1198.1 | 1277.4 | 47.64 | 0.69 | 0.41 | |||||||||||||||||
| 0.57 | Day 70 | 128.0 | 109.3 | 114.5 | 5.64 | 0.54 | 0.480.84 | 0.34 | 0.96 | <0.01 | 0.58 | Lactation feed intake, kg/day | 4.82 | 4.95 | 4.97 | 0.114 | 0.85 | 0.62 | 0.77 | ||||
| CV | 24.6 | 24.4 | 25.1 | 1.17 | 0.97 | 0.85 | 0.89 | ||||||||||||||||
| Milk (21 days) | |||||||||||||||||||||||
| 5.77 | 6.26 | 5.91 | 0.134 | 0.32 | 0.86 | 0.14 | Urea, mg/dL | ||||||||||||||||
| Total Solid, % | Day 70 | 26.8 | 25.0 | 25.5 | 0.97 | 0.85 | 0.73 | ||||||||||||||||
| Colostrum | 21.9 | 20.1 | 20.9 | 0.62 | 0.76 | 0.67 | 0.67 | 0.430.99 | 0.01 | 0.93 | Day 90 | 114.8 | 133.8 | 127.3 | 6.99 | 0.54 | 0.58 | 0.35 | |||||
| Day 90 | 24.1 | 25.6 | |||||||||||||||||||||
| Milk (21 days) | 16.9 | 26.7 | 1.22 | 0.69 | 0.42 | 19.2 | 0.83 | 18.1 | 0.54 | 0.23 | 0.49 | Piglet Distribution into BW Classes at day 21 | |||||||||||
| 0.12 | Day 110 | 97.8 | 89.7 | 98.9 | 4.19 | 0.68 | Day 110 | 0.80 | 0.40 | 30.6 | |||||||||||||
| Solid not Fat, % | 31.1 | 30.1 | 1.22 | 0.96 | 0.84 | 0.83 | <3 kg, % | 9.5 | |||||||||||||||
| Colostrum | 8.1 | 13.3 | 9.3 | 12.71.41 | 11.90.91 | 0.99 | 0.67 | ||||||||||||||||
| Cystine, μmol/L | 0.68 | 0.74 | 0.44 | 0.93 | 3–4 kg, % | 11.5 | 14.0 | 11.9 | 1.46 | 0.77 | 0.99 | 0.48 | |||||||||||
| Day 70 | 2.00 | 0.33 | 1.00 | 0.408 | 0.38 | 0.47 | 0.20 | 0.70 | 0.50 | 0.75 | |||||||||||||
| Milk (21 days) | 10.7 | 11.0 | 10.6 | 0.17 | 0.64 | 0.66 | 0.41 | 4–5 kg, % | 25.8 | 22.1 | 23.3 | ||||||||||||
| Day 90 | 0.88 | 1.50 | 1.29 | 2.39 | 0.83 | 0.72 | 0.62 | ||||||||||||||||
| 0.308 | 0.71 | 0.68 | 0.49 | 5–6 kg, % | 26.3 | 29.9 | 19.4 | ||||||||||||||||
| Day 110 | 3.40 | 2.33 | 2.17 | 0.14 | 1.00 | 0.928 | 0.57 | 0.12 | 0.19 | ||||||||||||||
| 0.31 | 0.93 | 6–7 kg, % | 20.2 | 17.9 | 23.9 | 2.01 | 0.48 | 0.36 | 0.42 | ||||||||||||||
| >7 kg, % | 6.7 | 8.0 | 12.2 | 1.80 | 0.41 | 0.19 | 0.86 | ||||||||||||||||
The blood concentration of AAs in the gestating sows is presented in Table 5. Increasing arginine supplementation to late-gestating sows linearly increased (
p
p
p
p
p
p
p
p
p < 0.05) than those for CON and ARG10 treatments.
Table 56.
| Treatment | 1 | SEM | 2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glutamic Acid, μmol/L | |||||||||||||
| Day 70 | |||||||||||||
| 240.5 | |||||||||||||
| 260.3 | |||||||||||||
| 198.0 | |||||||||||||
| 30.72 | |||||||||||||
| 0.72 | |||||||||||||
| 0.58 | 0.66 | 0.60 | 0.07 | 0.81 | |||||||||
| Day 90 | 173.3 | 211.6 | 189.3 | 16.13 | 0.63 | 0.81 | 0.36 | ||||||
| Day 110 | 170.6 | 153.8 | 152.6 | 13.82 | 0.84 | 0.64 | 0.76 | ||||||
| Glutamine, μmol/L | |||||||||||||
| Day 70 | 260.5 | 220.7 | 249.5 | 23.86 | 0.84 | 0.95 | 0.58 | 0.27 | 0.01 | 0.48 | |||
| Day 90 | 343.8 | 328.9 | 289.4 | 14.64 | 0.32 | 0.14 | 0.88 | ||||||
| Day 110 | 299.8 | a | 330.8 | a | 259.9 | b | 10.39 | 0.03 | 0.04 | 0.04 | |||
| Glycine, μmol/L | |||||||||||||
| Day 70 | 1224.5 | 1186.7 | 1073.5 | 38.59 | 0.27 | 0.13 | 0.85 | 0.58 | 0.03 | 0.76 | |||
| Day 90 | 1206.0 | 1291.5 | 1204.9 | 30.62 | 0.43 | 0.83 | 0.20 | ||||||
| Day 110 | 1070.9 | 1105.3 | 1117.9 | 39.67 | 0.88 | 0.66 | 0.85 | ||||||
| Histidine, μmol/L | |||||||||||||
| Day 70 | 93.0 | 103.3 | 93.0 | 5.31 | 0.72 | 0.90 | 0.46 | 0.36 | 0.21 | 0.98 | |||
| Day 90 | 103.9 | 116.5 | 96.6 | 6.80 | 0.51 | 0.56 | 0.31 | ||||||
| Day 110 | 90.8 | 97.0 | 88.7 | 3.60 | 0.69 | 0.71 | 0.42 | ||||||
| Isoleucine, μmol/L | |||||||||||||
| Day 70 | 107.0 | 121.3 | 104.0 | 6.11 | 0.51 | 0.72 | 0.32 | 0.65 | 0.81 | 0.92 | |||
| Day 90 | 120.4 | 125.8 | 115.7 | 3.46 | 0.52 | 0.50 | 0.35 | ||||||
| Day 110 | 134.2 | 119.5 | 109.4 | 11.82 | 0.69 | 0.42 | 0.84 | ||||||
| Leucine, μmol/L | |||||||||||||
| Day 70 | 210.5 | 220.3 | 202.8 | 10.62 | 0.82 | 0.74 | 0.66 | 0.66 | 0.02 | 0.84 | |||
| Day 90 | 217.6 | 228.4 | 205.7 | 6.21 | 0.36 | 0.35 | 0.26 | ||||||
| Day 110 | 194.8 | 165.5 | 168.0 | 13.38 | 0.61 | 0.48 | 0.54 | ||||||
| Lysine, μmol/L | |||||||||||||
| Day 70 | 281.5 | 321.0 | 239.5 | 18.85 | 0.17 | 0.24 | 0.19 | 0.44 | <0.01 | 0.71 | |||
| Day 90 | 409.4 | 423.6 | 438.9 | 18.45 | 0.83 | 0.55 | 0.93 | ||||||
| Day 110 | 282.4 | 326.5 | 296.3 | 14.23 | 0.48 | 0.85 | 0.25 | ||||||
| Methionine, μmol/L | |||||||||||||
| Day 70 | 48.5 | 57.7 | 50.8 | 2.80 | 0.47 | 0.92 | 0.24 | 0.48 | <0.01 | 0.92 | |||
| Day 90 | 73.0 | 72.4 | 68.6 | 2.48 | 0.76 | 0.47 | 0.86 | ||||||
| Day 110 | 58.6 | 62.3 | 59.0 | 2.20 | 0.79 | 0.96 | 0.50 | ||||||
| Ornithine, μmol/L | |||||||||||||
| Day 70 | 208.0 | 163.3 | 146.0 | 17.04 | 0.43 | 0.22 | 0.58 | 0.12 | <0.01 | <0.01 | |||
| Day 90 | 149.6 | b | 194.1 | a | 228.1 | a | 9.85 | <0.01 | <0.01 | 0.32 | |||
| Day 110 | 114.4 | b | 143.8 | b | 184.6 | a | 9.38 | <0.01 | <0.01 | 0.80 | |||
| Phenylalanine, μmol/L | |||||||||||||
| Day 70 | 93.0 | 98.0 | 84.5 | 13.60 | 0.53 | 0.42 | 0.52 | 0.24 | 0.07 | 0.77 | |||
| Day 90 | 85.5 | 104.8 | 85.6 | 4.51 | 0.13 | 0.75 | 0.05 | ||||||
| Day 110 | 79.3 | 81.7 | 76.4 | 3.83 | 0.89 | 0.72 | 0.71 | ||||||
| Proline, μmol/L | |||||||||||||
| Day 70 | 408.0 | 422.0 | 379.5 | 21.51 | 0.73 | 0.59 | 0.67 | 0.75 | 0.21 | 0.75 | |||
| Day 90 | 389.5 | 400.6 | 434.3 | 13.71 | 0.42 | 0.19 | 0.87 | ||||||
| Day 110 | 349.8 | 386.5 | 377.7 | 15.44 | 0.60 | 0.55 | 0.47 | ||||||
| Serine, μmol/L | |||||||||||||
| Day 70 | 186.5 | 179.7 | 153.8 | 7.66 | 0.17 | 0.09 | 0.75 | 0.26 | 0.08 | 0.83 | |||
| Day 90 | 167.6 | 215.6 | 158.0 | 16.59 | 0.33 | 0.64 | 0.16 | ||||||
| Day 110 | 139.5 | 153.3 | 134.6 | 7.01 | 0.61 | 0.65 | 0.35 | ||||||
| Taurine, μmol/L | |||||||||||||
| Day 70 | 141.5 | 119.0 | 83.0 | 13.35 | 0.22 | 0.10 | 0.96 | 0.13 | <0.01 | 0.14 | |||
| Day 90 | 83.0 | 92.9 | 88.4 | 4.40 | 0.67 | 0.72 | 0.42 | ||||||
| Day 110 | 67.8 | 66.2 | 72.9 | 2.96 | 0.68 | 0.44 | 0.62 | ||||||
| Threonine, μmol/L | |||||||||||||
| Day 70 | 177.0 | 191.7 | 161.3 | 13.47 | 0.68 | 0.61 | 0.56 | 0.05 | 0.05 | 0.82 | |||
| Day 90 | 138.1 | 149.1 | 126.0 | 6.86 | 0.42 | 0.39 | 0.31 | ||||||
| Day 110 | 133.2 | 167.2 | 105.6 | 11.06 | 0.11 | 0.16 | 0.08 | ||||||
| Tryptophan, μmol/L | |||||||||||||
| Day 70 | 61.0 | 76.0 | 62.5 | 4.37 | 0.36 | 0.92 | 0.19 | 0.10 | 0.40 | 0.85 | |||
| Day 90 | 62.8 | 67.1 | 64.1 | 2.35 | 0.75 | 0.91 | 0.46 | ||||||
| Day 110 | 54.6 | 66.5 | 60.0 | 3.00 | 0.29 | 0.62 | 0.15 | ||||||
| Tyrosine, μmol/L | |||||||||||||
| Day 70 | 98.0 | 104.7 | 91.5 | 7.10 | 0.77 | 0.69 | 0.63 | 0.17 | 0.23 | 0.96 | |||
| Day 90 | 98.1 | 120.9 | 97.7 | 6.37 | 0.24 | 0.77 | 0.10 | ||||||
| Day 110 | 87.1 | 101.3 | 89.7 | 4.03 | 0.36 | 0.99 | 0.16 | ||||||
| Valine, μmol/L | |||||||||||||
| Day 70 | 306.5 | 330.3 | 284.0 | 15.04 | 0.46 | 0.48 | 0.39 | 0.25 | 0.01 | 0.93 | |||
| Day 90 | 276.9 | AB | 293.6 | A | 253.4 | B | 7.49 | 0.09 | 0.12 | 0.10 | |||
| Day 110 | 252.0 | 230.3 | 202.7 | 19.14 | 0.57 | 0.31 | 0.93 | ||||||
The dietary arginine levels for the late-gestating sows had no significant influence on the blood concentrations of BUN, creatinine, total protein, and urea for sows in the late gestation period (Table 6). The blood concentrations of BUN, creatinine, and urea for day 110 of gestation were greater (
p < 0.05) than those for day 70 of gestation. There were no interactions between arginine and the date effect in the blood concentrations for BUN, creatinine, total protein, and urea. Increasing the dietary arginine levels for sows during late gestation did not affect the milk composition of colostrum and milk (day 21 of lactation) in lactating sows (Table 7).
Table 67.
| Treatment | 1 | SEM | 2 | p | -Value | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
4. Discussion
4. Discussion
Supplementing sow diet with L-arginine during the late gestation did not influence the gain of BW and BF in sows for the late gestation period and loss of BW and BF in sows for the lactation period. This was in agreement with the result of Quesnel et al. [25], who reported that dietary supplementation with 25.5 g/day L-arginine from day 77 to farrowing did not affect the BW after farrowing and BF before farrowing of sows. Additionally, Bass et al. [26] reported that dietary supplementation with 1% L-arginine for gestating sows from day 93 to day 110 had no influence on BW loss, following farrowing or during lactation. However, they observed that improved late gestation BW gain of sows fed a diet with 1% L-arginine was revealed in parity 0 and parity 1, whereas, there was no difference in late gestation BW gain compared to the control animals and 1% L-arginine-supplemented sows in parity 2+, implying that supplementation with 1% L-arginine partially met the arginine requirement in gilts or sows with parity 1 sows, not in sows with parity 2+. The difference between the current study and that of Bass et al. [26] with regard to the effect of dietary arginine in late gestation on the changes of BW or BF for sows could have been due to the difference in the parities of sows in the treatment group. Additionally, the lack of difference in the changes of BW and BF for sows during the whole experiment period could partly have been due to the same nitrogen content of diets among the dietary treatments, with the addition of L-Alanine as the isonitrogenous control.
References
Dietary effects of L-arginine was reported such that supplementation of L-arginine was found to increase nitric oxide, enhancing the delivery of essential nutrients from maternal to fetal blood [27][28][29], and increasing the polyamines necessary for embryogenesis and placental growth [30][31][32]. Since the nitric oxide and polyamines are important for angiogenesis and embryogenesis, arginine enhanced the growth of fetus and placenta development [32][33]. The litter size, including the number of total born or born alive was not affected by the dietary arginine effect in the present study, which was in agreement with the results from the studies of Quesnel et al. [25] and Nantapaitoon et al. [34], who reported that 1% L-arginine supplementation for late-gestating sows had no influence on the number of piglets at birth. However, previous studies reported that arginine supplementation enhanced conceptus survival or sow litter size in the period of early-gestation [19][35], or whole gestation [21][22][24]. The number of developing embryos is decided in early gestation, because most of the embryonic losses occurred during early gestation [36], and the dead embryos were not reabsorbed by the uterus after day 40 of gestation [37]. Thus, the differences among studies with regard to the effects of dietary arginine on the litter size of sows could have been due to differences in the supplemented period for the diet with L-arginine.
- Quesnel, H.; Quiniou, N.; Roy, H.; Lottin, A.; Boulot, S.; Gondret, F. Supplying dextrose before insemination and-arginine during the last third of pregnancy in sow diets: Effects on within-litter variation of piglet birth weight. J. Anim. Sci. 2014, 92, 1445–1450. [Google Scholar] [CrossRef]
- Bass, B.E.; Bradley, C.L.; Johnson, Z.B.; Boyd, R.D.; Usry, J.L.; Maxwell, C.V.; Frank, J.W. Influence of dietary L-arginine supplementation to sows during late gestation on sow and litter performance during lactation. Res. Ser. 2011, 597, 151–155. [Google Scholar]
- Nuntapaitoon, M.; Muns, R.; Theil, P.K.; Tummaruk, P. L-arginine supplementation in sow diet during late gestation decrease stillborn piglet, increase piglet birth weight and increase immunoglobulin G concentration in colostrum. Theriogenology 2018, 121, 27–34. [Google Scholar] [CrossRef]
- Bérard, J.; Bee, G. Effects of dietary L-arginine supplementation to gilts during early gestation on foetal survival, growth and myofiber formation. Animal 2010, 4, 1680–1687. [Google Scholar] [CrossRef]
- Foxcroft, G.R.; Dixon, W.T.; Novak, S.; Putman, C.T.; Town, S.C.; Vinsky, M.D.A. The biological basis for prenatal programming of postnatal performance in pigs. J. Anim. Sci. 2006, 84, E105–E112. [Google Scholar] [CrossRef]
- Town, S.C.; Patterson, J.L.; Pereira, C.Z.; Gourley, G.; Foxcroft, G.R. Embryonic and fetal development in a commercial dam-line genotype. Anim. Reprod. Sci. 2005, 85, 301–316. [Google Scholar] [CrossRef]