Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Hadi Boukhatmi and Version 2 by Camila Xu.
Muscle stem cells (MuSCs) are essential for muscle growth, maintenance and repair. Skeletal musculature is one of the largest organs of the human body, comprising more than 600 muscles that enable body motion. Deciphering the molecular and cellular features of the MuSC would provide novel insights into their regenerative potential and to improve therapeutic strategies. Drosophila has proven to be a suitable model to study MuSC and offers a combination of genetics, live imaging and genomics approaches to uncover general principles governing MuSC biology in vivo.
- muscle stem cells
- satellite cells
- Drosophila
- myogenesis
1. Introduction
Skeletal muscles are made of multinucleated myofibers and possess the contractile properties to generate forces. The regeneration of adult muscles, being terminally differentiated, relies on a resident population of stem cells called Satellite Cells (MuSCs), first described by A. Mauro [1][2]. MuSCs reside underneath the basal lamina and, in healthy individuals, they are remarkably efficient to ensure the homeostasis and regeneration of the skeletal muscles [2][3][3,4].
Muscle repair is a multistep process. Upon muscle damage, MuSCs proliferate, migrate and divide asymmetrically to give rise to new stem cells and myogenic progenitors, which will ultimately differentiate and fuse with each other or with existing fibers to repair the muscle [4][5][5,6]. The cellular and molecular mechanisms regulating MuSC biology are presently a hot topic of investigation: both for answering basic stem cell questions and for possible therapeutic use in treating muscle-degenerative diseases [6][7][8][9][7,8,9,10]. To this end, a multitude of new experimental models is emerging [10][11][12][13][14][11,12,13,14,15]. Among them is the fruit fly, Drosophila, in which the genetic and cellular control of MuSC during the course of development and throughout adulthood can be efficiently assessed [15][16][17][16,17,18].
Drosophila has a long history as a genetic model to study myogenesis, both the muscle structure and core myogenic programs being highly conserved between flies and mammals [18][19][20][19,20,21]. Drosophila myogenesis proceeds in two distinct waves, leading to the formation of adequate sets of muscles for the stage-specific modes of locomotion. The first wave happens during embryonic development, and it forms the body wall muscles required for larval crawling. This group of muscles undergoes histolysis from larval to adult histolysis, the pupal transition. The second myogenic wave takes place during the metamorphosis (pupal stages) and gives rise to the adult musculature that allows the animal to feed, walk and fly [18][19]. Adult muscles are formed from a specific population of MuSC, known as adult muscles precursors (AMPs). They are specified in parallel to the first myogenic wave but are set apart and remain undifferentiated during the whole larval development and ultimately differentiate during metamorphosis to form the adult musculature [21][22][22,23]. The AMPs share several features with the vertebrates MuSCs [17][23][18,24].
2. Making a Muscle Stem Cell
2.1. Specification and Positioning of the AMPs
In each embryonic abdominal hemi-segment, there are six abdominal AMPs located in dorsal (D-AMP), dorsolateral (DL-AMP), lateral (L-AMP) and ventral (V-AMP) positions (Figure 1A). While abdominal AMPs are closely associated with the larval muscles and nerves [17][19][18,20], thoracic AMPs associates with the wing and leg imaginal discs (Figure 1B) [19][20]. They express markers specific to muscle progenitors such as the basic helix-loop-helix (bHLH) transcription factor Twist (Twi) [24][25][25,26]. In the embryo, AMPs are distributed in a stereotyped pattern that determines the final location of the adult muscles they will form (Figure 1A) [25][26][26,27]. Abdominal AMPs will form the adult body wall muscles, and thoracic AMPs associated with the wing and leg discs ultimately form the flight and leg muscles, respectively [21][22]. Thus, the early steps of AMPs specification are critical to building a proper and complete adult musculature.
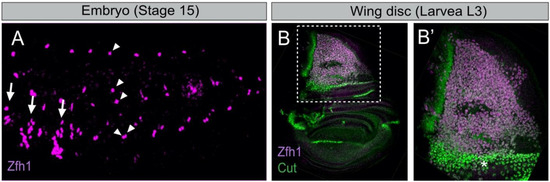
Figure 1. The Drosophila embryonic and wing disc associated muscle stem cells express the transcription factor Zfh1. (A) Stage 15 embryo stained for Zfh1 (magenta). Arrows and arrowheads indicate the thoracic and abdominal AMPs, respectively (adapted from [27][28]). The ventral abdominal AMP (V-AMP) is not shown in this sample. (B) Zfh1 (magenta) and Cut (green) are expressed in the wing disc AMPs. (B’) Higher magnification of boxed region in B shows that high Cut expressing AMPs (asterisk) have low levels of Zfh1 (adapted from [15][16]).
AMP patterning is, at least in part, governed by the Hox genes, which are differentially expressed along the anterior–posterior axis of the embryo [28][29][29,30]. For example, the gain of abdominal-A function converts the thoracic AMPs into abdominals AMPs suggesting that the Hox inputs control the spatial positioning of the AMPs [30][31]. Cell–cell communication between the abdominal AMPs also affects their positioning. They send out long cellular processes, which follow the peripheral nervous system and form a network of interconnected cells [31][32]. Ablation of these cellular processes perturbs their patterning and leads to excessive AMPs number, highlighting an additional level of control in their positioning [32][33].
The wing disc AMPs form a large pool of myoblasts, located in the notal part of the disc, underneath the epithelial cells (Figure 1B) [33][34]. The disc epithelial cells act as transient niche and provide cues governing AMP proliferation, maintenance and positioning. The role of the epithelial cells in localizing the AMPs at the right place was recently characterized by Everetts et al. [34][35]. This work revealed the contribution of the FGF signaling in guiding the AMPs to their notal localization. While the FGF-family ligands thisbe (ths) and pyramus (pyr) are detected in the epithelial cells of the notum, the receptor heartless (htl) is specifically expressed in the AMPs. Ectopic expression of the FGF ligands either in the pouch region or along the dorso-ventral axis provokes AMPs spreading towards the corresponding regions. Conversely, loss of either FGF-ligands or of Htl resulted in a reduction of AMPs number linked to increased apoptosis. Collectively, these results point to the role of FGF signaling both in localizing the AMPs to the notum region and sustaining their proliferation and survival.
2.2. The Control of the AMPs Diversity
Similar to vertebrate MuSCs, Drosophila AMPs are heterogeneous and express different markers [32][35][36][37][33,36,37,38]. Each abdominal AMP derives from the asymmetric division of a muscle progenitor (MP) that gives rise to both an AMP and a skeletal muscle founder cell (FC) [38][39][39,40]. While the AMPs remain quiescent, the FCs undergo several rounds of fusion with fusion-competent myoblasts to form the larval muscles [40][41]. The larval muscle shape, size and orientation reflect the early expression of specific combinations of ‘identity’ transcription factors (iTFs) in each FC [39][41][40,42]. Hence, every FC has an intrinsic code of iTFs that dictates the properties of the muscle they will form. Likewise, AMPs also differ by the expression of specific iTFs that control their fate and their competence to contribute to different muscle types. One well-studied AMP iTF is Ladybird early (Lbe; the Drosophila orthologue of mammalian Lbx1), which is expressed and required for the specification of the lateral abdominal AMPs (L-AMPs) [32][33]. Lbe is also involved in instructing the identity of leg-associated AMPs by dictating the shape, the structure and the functional properties of the leg muscles deriving from this population [42][43].
Wing disc-associated AMPs form two different types of adult flight muscles: the fibrillar indirect flight muscles (IFMs) and the tubular direct flight muscles (DFMs) [18][19][19,20]. These muscles have distinct physiologies, size, contractile properties and thus provided a well-suited system to study the mechanisms behind the early MuSCs divergence during development. The contribution of iTFs in such a divergence was first reported by Sudarsan et al. [33][34]. The studies identified two pioneer iTFs; Vestigial (Vg) and Cut (Ct), and showed that they are differentially expressed in the wing disc AMPs. AMPs expressing high levels of Vg and low levels of Ct form the IFMs, while the other AMPs (high Ct, no Vg) are required for the formation of DFMs (Figure 1B). Vg is activated by the Wingless signal emanating from the adjacent notal epithelial cells. Vg activates the expression of spalt-major (salm), a zinc finger TF [43][44]. Salm is a master regulator of the muscle fibrillar fate, which activates IFM-specific genes and repress genes involved in tubular muscle formation. Consistently, the ectopic expression of Salm in developing leg muscles is sufficient to switch their fate from a tubular to fibrillar organization [43][44][44,45]. Interestingly, the morphology of the flight muscle mitochondria is also determined by Salm [45][46]. salm expression is regulated by the homeodomain proteins Extradenticle (Exd) and Homothorax (Hth), which contribute to the fibrillar muscle fate [46][47]. Thus, Vg, Salm, Exd and Hth transcriptional cascades specify the IFM fate by promoting the expression of the fibrillar-specific genes in AMPs.
Besides Vg and Ct, little was known about the mechanism that distinguishes between AMPs leading to IFMs versus DFMs. Recent studies using single-cell transcriptomics (scRNA-seq) have considerably improved our understanding of the AMPs diversification [47][48][48,49]. Indeed, Zappia MP et al. [49][50] conducted single-cell RNA-sequencing on wing imaginal discs and showed that AMPs responsible for the formation of IFMs and DFMs have distinct transcriptional signatures and identified new genes differentially expressed between the two populations. Among them, the TF Zfh1 (the Drosophila homolog of ZEB1/ZEB2) was found to be highly expressed in the IFMs population, consistently with the dynamics of Zfh1 expression during AMPs specification (Figure 1B) [15][16]. Zappia et al. have also reported the existence of a large set of new DFM-specific genes, including kirre, midline (mid) and tenascin accessory (ten-a). Moreover, each of the two populations of AMPs is intrinsically heterogeneous and can be clustered in subpopulations (e.g., according to expression levels of the Notch target genes) that represent various states of myoblasts differentiation. Further analysis of one gene shed in light by this work, amalgam (ama), which encodes a membrane receptor, showed that its inactivation causes severe muscle phenotypes. This work thus provides compelling evidence that scRNA-seq can identify genes differentially transcribed in the two AMPs populations, while the specific functional requirement of these genes in the process of AMPs diversity remains to be fully characterized.
When and how can AMPs initiate and maintain a specific transcriptional program? It has been shown that extrinsic signals emanating from the disc epithelium are important for patterning [33][34]. Wingless signaling that specifies the IFMs lineage is produced from the epithelial wing disc cells closely associated with IFM AMPs, which maintain high levels of Vg [33][34]. This regulation involves the importin Moleskin (Msk) that regulates the Wingless effector β-catenin/Armadillo (Arm) by controlling its stability and/or nuclear transport [50][51][51,52]. Conversely, the Hedgehog (Hh) pathway is required for the specification of DFM AMPs [34][35]. The Hh ligand is produced by a subset of posterior epithelial cells in close proximity to the AMPs. Although components of the Hedgehog pathway smoothened (smo) and cubitus interruptus (ci) are uniformly expressed in most AMPs, the patched (ptc) receptor is restricted to a subset of DFMs AMPs. Ptc expression expanded through the majority of AMPs when exogenous Hedgehog activity was supplied. Reciprocally, the reduction of smo levels in AMPs was sufficient to abolish ptc expression and to induce defects in adult DFMs. This work suggests that the specification of the DFMs AMPs is controlled by Hh signals emanating from the epithelial cells and supports the view that the microenvironment is pivotal in establishing the AMPs diversity.
3. Role of the Microenvironment in Muscle Stem Cell Maintenance and Activation
3.1. Connecting to the Muscles; ‘Homing Behavior’
Once specified and positioned in the right place, abdominal AMPs lie dormant during embryogenesis until the beginning of the larval life. To investigate how AMPs are maintained in a dormant state, Aradhya et al. [52][53] generated an AMP-sensor line (m6-gapGFP) that enables the visualization of cell shape changes and behavior of the AMPs during development. As described previously [32][33], embryonic AMPs send out long protrusions and form a network of interconnected cells. In addition, Aradhya et al. showed that AMPs produce numerous smaller filopodia tightly associated with neighboring muscles. The interconnecting cellular processes regulate the maintenance of AMPs since their ablation pushes the AMPs to proliferate prematurely. These connections persist until the first instar larval stage, and they are lost in the second instar larvae. The short filopodia remain, however, associated with the muscles, illustrating that, as with a vertebrate’s satellite cells [53][54], Drosophila AMPs display a homing behavior, likely necessary to sense and respond to instructive signals provided by the muscle fibers.
3.2. Muscle-Driven Insulin Signal Reactivates Dormant AMPs
At the mid-second larval instar, the AMPs are reactivated, exit the quiescent state and enter proliferation to provide the myoblasts that form the adult muscles. The important question is to understand what regulates the transition from a quiescent to an activated state. Aradhya et al. [52][53] showed that the AMP reactivation is driven by the neighboring muscles, which provide insulin-like peptide 6 (dilp6) to activate the insulin pathway in the AMPs. Filopodia of muscle-associated AMPs facilitate reception of the dilp6 signal from the muscle niche. Subsequently, insulin signaling triggers the Notch pathway in a ligand-independent way, involving the ubiquitin ligase Deltex. Genetic epistasis revealed that AMP proliferation is induced by dMyc, acting downstream of Notch. Thus, the AMPs reactivation requires a nutrient-dependent switch that is sensed by cell processes reaching the surrounding muscles [54][55].
3.3. Interplay between the AMPs and Motor Neurons
Soon after their specification, abdominal AMPs exhibit a round shape and are found in the vicinity of motor axons [26][31][32][55][27,32,33,56]. At later embryonic stages, AMPs elongate and send out long cellular processes that follow the main branches of the peripheral nervous system. Lavergne et al. [55][56] recently explored further the interactions between AMPs and the navigating motor axons during embryonic development. Using high-resolution imaging, they showed that the AMPs direct their filopodia towards the axons, suggesting that AMP protrusions may play an active role in guiding them. These studies revealed that one motor axon makes the first contact with a dorsolateral AMP (DL-AMP) and then a second contact with a dorsal AMP (D-AMP) before finding its final destination. They further showed that loss or mispositioning of AMPs affects pathfinding, branching and leads to defective muscle innervation. Interestingly the guiding molecules Sidestep and Side IV were found to be specifically expressed in some AMPs, suggesting a putative role in neuron’s pathfinding. This work showed for the first time that the muscle stem cells dynamically interact with the navigating neurons and ultimately contribute to the proper formation of the neuro-muscular system.