Nitric oxide (NO) is a gas molecule with diverse physiological and cellular functions. In the eye, NO is used to maintain normal visual function as it is involved in photoreceptor light transduction. In addition, NO acts as a rapid vascular endothelial relaxant, is involved in the control of retinal blood flow under basal conditions and mediates the vasodilator responses of different substances such as acetylcholine, bradykinin, histamine, substance P or insulin. However, the retina is rich in polyunsaturated lipid membranes and is sensitive to the action of reactive oxygen and nitrogen species. Products generated from NO (i.e., dinitrogen trioxide (N2O3) and peroxynitrite) have great oxidative damaging effects. Oxygen and nitrogen species can react with biomolecules (lipids, proteins and DNA), potentially leading to cell death, and this is particularly important in the retina.
1. Introduction
Nitric oxide (NO) is a gas-signaling molecule with a short half-life and diverse physiological and cellular functions
[1]. It diffuses across cell membranes and cannot be stored in the human body.
In most cases, NO is synthesized by a reaction in which L-arginine is converted to
l-citrulline, with an intermediate reaction product, NG-hydroxy-
l-arginine. This reaction is catalyzed by the nitric oxide synthase (NOS) enzyme with the presence of oxygen and nicotinamide adenine dinucleotide phosphate (NADPH)
[2]. Vertebrates express three isoforms of the enzyme NOS: i) the constitutive calcium-dependent neuronal NOS (nNOS or NOS-I); ii) the endothelial NOS isoform (eNOS or NOS II) and iii) the inducible or calcium independent NOS isoform (iNOS or NOS III)
[3]. Flavin adenine dinucleotide (FAD), flavin mononucleotide (FMN) and (6
R-)5,6,7,8-tetrahydro-l-biopterin (BH
4) are cofactors for NOS isozymes
[4]. The availability of the substrate and cofactors (NADPH, FMN, FAD, BH
4) is important for the proper enzyme functionality; a low level of L-arginine or the oxidation of TBH4 protein can cause protein dysfunction.
Typically, eNOS is expressed in vascular endothelial cells; iNOS is mainly associated with inflammation or pathological states, and nNOS is usually expressed in the neurons of the peripheral and central nervous system but also in human skeletal muscles
[5]. nNOS and eNOS are constitutive, and nNOS is responsible for the largest proportion of constitutive NOS in humans. iNOS requires induction by immunological factors such as lipopolysaccharide, interferon, and tumor necrosis factor
[6].
l-arginine analogs inhibit NOS enzymes.
An alternative and important source of NO generation is the recycling of nitrates and nitrites. This NO synthesis route is named the nitrate–nitrite–nitric oxide pathway and is particularly relevant in hypoxic states
[7].
The only known NO receptor is the enzyme soluble guanylate cyclase. Thus, NO binds guanylate cyclase and increases intracellular cGMP concentrations
[7]. Nevertheless, NO may as well be transformed into reactive nitrogen species (RNS)
[8]. The term nitrosative stress describes the ability of NO to react with molecules such as proteins and DNA, and these reaction products may be harmful for the cells
[9].
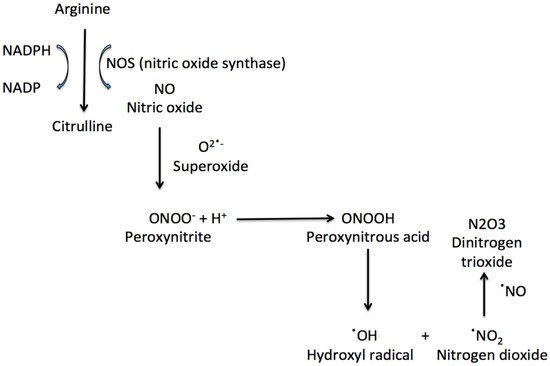
NO can react with superoxide anions to form peroxynitrite (ONOO). ONOO formation induces cytochrome c release, which can induce cell death and which blocks the respiratory chain
[8]. Peroxynitrous acid (ONOOH), peroxynitrite conjugate acid, reacts with nitrate without forming strong oxidant intermediates. ONOOH can also form OH and nitrogen dioxide, which can initiate fatty acid oxidation and amino acids nitration (
Figure 1). The peroxynitrite reactivity is also affected by carbon dioxide, with the formation of a reactive nitrosoperoxocarbonate
[10].
Figure 1. The synthesis of nitric oxide and some important nitrosative species.
NO can also react with proteins. S-nitrosation is a reaction of the NO moiety with the low-molecular thiols or cysteine amino acid residue of proteins to form S-nitrosothiols. An important intermediate reaction product in nitrosation is S-nitrosoglutathione
[9]. Nitration is a chemical reaction resulting in the formation of products such as 3-nitrotyrosine
[9]. Both the products of S-nitrosation and nitration may damage the cell. The dysregulation of nitrosation and nitration has been linked to human neurodegenerative disorders and is mostly related to the excessive production of NO that takes place through the excessive nNOS or iNOS activity via neuroinflammatory stimuli or several toxin actions.
[11][12][11,12]. Oxidative stress has been shown to convert eNOS from an NO-producing enzyme to an enzyme that generates O
2•−. This process has been referred to as NOS uncoupling
[4] and may be important in vascular diseases, such as diabetic retinopathy.
2. Nitric Oxide in the Retina
Different retinal regions present an NO production. nNOS expression has been found in the inner retina
[13]. The nNOS isoform has been located anatomically by immunohistochemistry in the mouse retina in the outer plexiform layer (OPL), in bipolar, amacrine and ganglionar somas and in cellular processes in the internal plexiform layer (IPL)
[14][15][16][17][14,15,16,17]. Studies of the expression of different NOS isoforms in mouse retinas suggest that the immunoreactivity for nNOS reflects most of the production of NO
[15].
iNOS is not constitutively expressed; its expression is inducible by pro-inflammatory stimuli. It is known that iNOS is expressed in the inner nuclear layer (INL) in an in vivo murine model of proliferative ischemic retinopathy where there is marked damage to the internal retina
[18]. Other authors have shown that, after infection of the retina by a murine cytomegalovirus, most cells that express iNOS in the retina are macrophages and microglia
[19]. The presence of iNOS in the outer segments of the photoreceptors has been demonstrated in the retinal degeneration rd2 mouse
[20]. iNOS may be also important in the normal phagocytosis of the retinal outer segment, in ischemic processes and in diabetic retinopathy pathogenesis
[6].
eNOS expression in endothelial cells has even been detected in the human fetal eye
[21]. eNOS immunoreactivity has also been demonstrated in photoreceptors, horizontal cells, bipolar cells, amacrine cells, Müller cells, and in the ganglion cell layer of the avian retina
[22]. After retinal injuries in rats, such as ischemia/reperfusion, it has been shown that eNOS immunoreactivity increased in retinal vessels and in the ganglion cell layer
[23].
Regarding the retinal NO function, NO is used to maintain a normal visual function. It is involved in photoreceptor light transduction, as 3’,5’cyclic guanosine monophosphate (cGMP) production, an essential intermediate in the visual transduction cascade, is catalyzed from GTP by guanylate cyclase, a target enzyme for the action of NO
[24]. In addition, NO acts as a rapid vascular endothelial relaxant and is involved in retinal blood flow control under normal circumstances, and it mediates the vasodilator responses of different substances such as acetylcholine, bradykinin, histamine, substance P or insulin
[25].
However, the retina is rich in polyunsaturated lipid membranes and is especially sensitive to the action of reactive oxygen and nitrogen species
[3]. Despite NO itself being a radical, its reactivity is low compared to the possible damage generated by the oxidative products that it generates, such as dinitrogen trioxide (N
2O
3) and peroxynitrite (ONOO−). Both can react with biomolecules, which can lead to cell death
[26]. In this sense, NO has been related to several ocular diseases, including diabetic retinopathy, retinitis pigmentosa, glaucoma or age-related macular degeneration (AMD)
[26].
3. Diabetic Retinopathy
Diabetic retinopathy (DR) is one of the leading causes of blindness in adults in developed countries. DR is categorized into non-proliferative DR (NPDR) and proliferative DR (PDR) phases according to the presence of visible ophthalmologic changes and evidence of retinal neovascularization
[27].
NPDR is a consequence of hyperglycemia, which accompanies these patients and weakens the capillary walls, leading to the formation of microaneurysms. This is followed by the rupture of vessels, leading to lipid by-products deposition. In addition, the nerve fibre layer may be obstructed, resulting in the accumulation of white spots or cotton wool spots
[27]. NPDR is followed by PDR, which is characterized by neovascularization. These newly-formed vessels are leaky and fragile. They may also lead to hemorrhages and tractional retinal detachment. Macular oedema, the last stage of DR, is the principal cause behind the loss of visual acuity
[27].
As we have just described, DR has been classically considered a microvascular disease. However, increasing evidence suggests that retinal neuron death occurs before vascular changes. In this sense, DR can now be described as a microvascular disease but also as a neurodegenerative disease
[28].
Although various biochemical pathways may be the basis of the DR aetiology, the main insult to the retina is due to oxidative stress and inflammation
[27]. Hyperglycaemia and other stress stimuli (including oxidative stress) trigger the generation of superoxide, which reacts with NO, producing peroxynitrite. Peroxynitrite can lead to cell damage. A question that has not yet been completely answered is which of the three NOS isoforms is responsible for the damages observed in retinal cells during DR.
In 2010, Li et al.
[29] induced diabetes by streptozotocin injection in eNOS knockout (−/−) mice. Their results showed that eNOS −/− mice exhibited a more severe retinal vascular permeability than control mice did. eNOS −/− mice showed an earlier and increased number of acellular capillaries, increased capillary basement membrane thickness, gliosis and total NO-relative products (assessed by measuring nitrate/nitrite using a fluorometric-based assay). The authors suggested that this NO concentration increase was due to the elevated iNOS expression in the diabetic eNOS −/− retina
[29].
NOS requires a physiologically relevant amount of L-arginine to produce NO. When L-arginine is limited, NOS becomes uncoupled, causing it to produce superoxide that will react with NO to form ONOO
− and reducing NO. This NO decrease may be responsible for the diabetes-induced decreases in the retinal blood flow that have been observed in diabetic humans and DR animal models
[30]. It is well known that a physiologically relevant production of NO by eNOS is necessary for the maintenance of healthy vessels and a proper blood flow.
Other studies had focused on the relevance of iNOS in DR. Zheng et al.
[31] induced diabetes in control and iNOS −/− mice and found that the retinas were thinner, that the number of acellular capillaries and pericyte ghosts increased in the retinas of diabetic mice and that these changes were not observed in iNOS −/− mice. However, the deletion of iNOS had no effect on the diabetes-induced abnormalities usually observed by means of an electroretinogram.
In humans, Sharma et al.
[32] studied 60 diabetic patients and classified the severity of their DR according to an early treatment diabetic retinopathy study (ETDRS) classification. Their study demonstrated that increased NO plasma concentrations are associated with an increased diabetic retinopathy severity. The aqueous humour NO concentration has also been determined in type 2 diabetic patients that were subjected to cataract surgery, and was found to increase when compared with control patients
[33].
In diabetic human post-mortem retinas, immunoreactivity for iNOS has been detected in ganglion cells, glial cells and cells of the INL, and nitrotyrosine immunoreactivity has been detected in vascular endothelial cells
[34]. In retinas from subjects without diabetes, there was no iNOS or nitrotyrosine immunoreactivity
[34].
Can NO Inhibition Have a Role in Diabetic Retinopathy Therapy?
A great number of studies have shown that the antioxidant administration in animal models of diabetes can reduce retinal cell death by apoptosis, and can thus contribute to the improvement in DR progression
[35][36][37][35,36,37]. However, the results of clinical trials that have used antioxidants in DR therapy are inconclusive
[38]. Consequently, new studies and approaches using antioxidants as adjuvant therapies in DR treatment should be performed. In this sense, the possible use of antioxidants and/or NOS inhibitors and NO scavengers has been suggested
[39]. It has been reported that aminoguanidine, a pharmacological inhibitor of iNOS and an inhibitor of advanced glycated end products, prevents the histological changes induced by diabetes in rats and that it is safe when administered intravitreally
[40]. Other NOS inhibitors, such as N(G)-nitro-
l-arginine methyl ester (
l-NAME), are able to reduce the increase in oxidized proteins observed in diabetic rat retinas
[41]. Moreover, it is known that in diabetic retinas, iNOS and the bradykinin type 1 receptor contribute to inflammation, oxidative stress, and vascular dysfunction and that the administration of a selective iNOS inhibitor for diabetic rats decreases these alterations
[42]. Conversely, a large septic shock trial was terminated prematurely because of a trend toward harm among patients receiving NO inhibitors
[43].
Further studies are needed in order to know which of these molecules may be useful and to determine their optimal doses, toxicity, administration form, etc.